茉莉素與植物生物脅迫反應
2018-08-18 01:44:14吳德偉汪姣姣謝道昕
生物技術通報 2018年7期
吳德偉 汪姣姣 謝道昕
(1. 揚州大學生物科學與技術學院,揚州 225009;2. 清華大學生命科學學院,北京 100084)
茉莉素(Jasmonate)是一類新型植物激素,包括茉莉酸(JA)及其環戊烷酮類衍生物。茉莉素在植物界中廣泛存在,對植物的生存至關重要[1]。茉莉素既是植物生長發育的調節物,也是植物防御反應的信號分子。茉莉素調控植物生長發育的諸多方面,包括根的生長、雄蕊發育、葉片衰老、表皮毛形成等[1,2];同時,茉莉素也介導植物對病原侵染、昆蟲噬咬等多種生物脅迫和低溫、干旱、臭氧、紫外線等多種非生物脅迫的抗性反應[3-7]。近年來,茉莉素的生物合成、信號轉導和生理功能等方面均得到了廣泛研究,取得了許多重要進展。本文將主要從茉莉素對植物生物脅迫反應的調控和病原生物對植物茉莉素途徑的操控兩方面進行綜述。
1 茉莉素生物合成的調控機理
茉莉素的生物合成經歷了依次發生于葉綠體、過氧化物酶體和細胞質中的多步催化過程(圖1-A)。首先,葉綠體內膜上釋放出來的α-亞麻酸(α-LeA)在脂肪氧合酶(LOX)、丙二烯氧化物合成酶(AOS)、丙二烯氧化物環化酶(AOC)等酶的催化作用下形成12-氧-植物二烯酸(OPDA);隨后,OPDA被轉運至過氧化物酶體中,在OPDA還原酶3(OPR3)的作用下形成3-氧代-2(順2’-戊烯基)-環戊烷-1-辛酸(OPC-8),然后經過三輪β-氧化形成茉莉酸[8]。近期有研究表明,除了經典的OPR3途徑外,OPDA也可先經過三輪β-氧化形成4,5-雙脫氫茉莉酸(ddh-JA),然后經OPR2催化形成茉莉酸[9]。茉莉酸在過氧化物酶體中合成后會被釋放到細胞質進行進一步的修飾,形成多種茉莉酸的衍生物,如茉莉酸甲酯、茉莉酸氨基酸偶聯物等。其中,經JAR1催化形成的(+)-7-iso-JA-L-Ile(JA-Ile的一種構象)是植物體內最早被發現的具有活性的茉莉素形式[10-11]。 最 近 的 研 究 表 明,(+)-7-iso-JA-Leu、(+)-7-iso-JA-Val、(+)-7-iso-JA-Met 和(+)-7-iso-JA-Ala也是具有活性的植物內源茉莉素形式[12]。值得注意的是,JA-Ile具有四種不同的構象,但只有(+)-7-iso-JA-L-Ile這一種構象是具有生物活性的;同時,(+)-7-iso-JA-L-Ile不穩定,在環境因素(如pH等)的影響下很容易轉化為沒有生物活性的構象形式[10-12]。
茉莉素的合成受到嚴格的調控。在正常生長狀態下,植物體內的茉莉素處于較低水平,以降低茉莉素對植物生長的抑制作用;而當植物受到損傷等外界刺激或接收到特定生長發育信號時,植物體內的茉莉素含量迅速升高,從而激活茉莉素響應基因的表達[1,13]。
損傷如何誘導茉莉素的合成是茉莉素領域一個重要的科學問題。曾有研究表明,機械損傷能夠誘導植物細胞的膜電位變化,進而誘導茉莉素的快速合成和茉莉素響應基因的表達;而與離子通道調控相關的谷氨酸受體基因家族中部分基因(GLR3.2,GLR3.3和GLR3.6)的突變會抑制損傷誘導的膜電位變化和茉莉素響應基因表達,表明植物細胞離子通道狀態改變引起的膜電位變化在損傷誘導茉莉素合成過程中發揮重要作用[14]。最近的研究表明,VQ家族蛋白JAV1可能是連接離子通道狀態變化/細胞膜電位變化和細胞內茉莉素合成的關鍵分子[15]。當植物處于正常生長狀態時,JAV1蛋白能夠與JAZ8、WRKY51互作并形成JAV1-JAZ8-WRKY51蛋白復合體(簡稱JJW復合體),結合并抑制茉莉素合成基因的表達,維持植物體內茉莉素含量處于較低水平;而當植物受到昆蟲咬噬等損傷后,細胞內鈣離子濃度迅速升高,從而激活了鈣調蛋白依賴的JAV1磷酸化,而磷酸化的JAV1則不能與JAZ8互作、其亞細胞定位也發生改變(由細胞核定位轉變為細胞質定位)、并被26S蛋白酶體降解,從而使JJW復合體解體、解除了JJW復合體對茉莉素合成的抑制作用,最終導致茉莉素能夠在損傷后迅速大量合成[15-16](圖 1-B)。
2 茉莉素的信號轉導機制
1998年,COI1基因的克隆[17]開啟了茉莉素信號轉導研究的序幕,此后茉莉素信號通路中的關鍵因子陸續被分離出來,信號轉導過程逐漸清晰。目前認為,茉莉素信號轉導過程大致如下:當植物處于靜息狀態時,體內的茉莉素含量較低,大量積累的負調控因子JAZ蛋白與下游一系列轉錄因子結合并抑制其活性,從而抑制了茉莉素反應;而當植物受到傷害等外界刺激或接收到特定生長發育信號時,體內的茉莉素含量迅速升高,SCFCOI1復合體中的COI1蛋白與活性茉莉素分子結合,進而與底物JAZ蛋白相互作用并將其泛素化降解,解除了JAZ蛋白對下游轉錄因子的抑制作用,從而激活了下游茉莉素反應[5,8,18]。
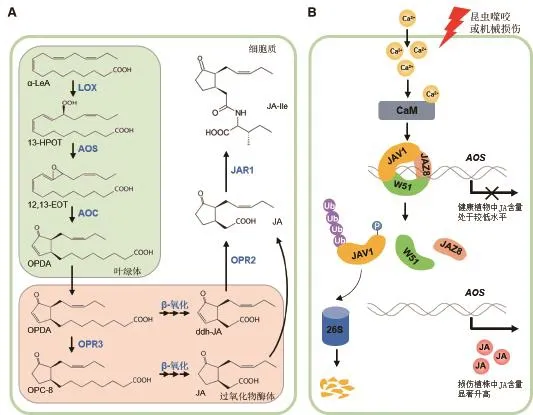
圖1 茉莉素的生物合成與調控
茉莉素受體COI1是一個F-box蛋白[17],可與SKP、CUL1和RBX1等蛋白形成SCFCOI1復合體[19],作為一種泛素連接酶特異性識別底物JAZ蛋白[20-22]。JAZ蛋白可以在茉莉素誘導下與COI1蛋白發生直接相互作用并被SCFCO1泛素化,進而被26S蛋白酶體所降解[21-22]。生化及晶體學實驗結果表明,COI1能直接與活性茉莉素分子JA-Ile或COR結合[23],而JAZ蛋白可進一步穩定COI1與JA-Ile的結合[23-25]。此外,SCFCOI1復合體的完整性對維持COI1蛋白自身的穩定性也至關重要,SCFCOI1復合體組分SKP、CUL1的突變會導致COI1蛋白的穩定性大大降低[26]。
作為茉莉素途徑的負調控因子,JAZ蛋白通過抑制下游一系列轉錄因子的活性、從而抑制各種茉莉素反應。已報道的可與JAZ蛋白直接互作的下游轉錄因子有20余個,包括MYC2、MYC3、MYC4、MYC5、bHLH3、bHLH13、bHLH14、bHLH17、MYB21、MYB24、FIL、ICE1、ICE2、EIN3、EIL1、TOE1、TOE2、RSS3、WRKY57、GL3、EGL3、TT8、MYB75、GL1 等[8,18]。JAZ 蛋白通過作用于不同的下游轉錄因子,介導茉莉素對植物生長發育及抗性不同方面的調控,例如,JAZ通過與MYC2、MYC3、MYC4互作介導茉莉素對植物抗性、根的生長和葉片衰老等方面的調控[27-31],通過與MYB21、MYB24互作介導茉莉素對植物育性的調控[32]。目前認為,JAZ蛋白至少可通過下列三種方式抑制下游轉錄因子的活性:(1)通過與下游轉錄因子互作,干擾下游轉錄因子與MED等轉錄共激活因子的結合[33-34];(2)通過直接招募 HDA6,抑制組蛋白乙酰化[35];(3)通過自身EAR結構域(如JAZ5/6/7/8)直接招募或通過NINJA間接招募轉錄共抑制因子 TPL[36-37]。
3 茉莉素對植物生物脅迫反應的調控
茉莉素在調控植物對昆蟲和腐生型真菌的抗性反應中發揮重要作用。茉莉素合成突變體(如fad3 fad7 fad8、aos、opr3等)和受體突變體(如coi1-1)對昆蟲和腐生型真菌的抗性都顯著低于野生型植物[1]。目前,已經鑒定到了一些調控植物對昆蟲和腐生型真菌抗性的茉莉素途徑基因,它們大多為轉錄因子,通過誘導植物表面物理屏障(如表皮毛等)的形成、調控次生代謝產物的合成、誘導防御基因表達等多種方式,介導茉莉素對植物生物脅迫反應的調控。
MYC2、MYC3和MYC4在調控茉莉素介導的植物抗性反應中發揮重要作用。MYC2、MYC3和MYC4功能冗余地調控植物對昆蟲的抗性,myc2 myc3 myc4三突變體對灰翅夜蛾(Spodoptera littoralis)和甜菜夜蛾(Spodoptera exigua)的抗性顯著低于野生型植物[28-29,38]。進一步研究發現,MYC2、MYC3和MYC4可通過促進芥子油苷的合成增強植物對昆蟲的抗性:MYC2、MYC3和MYC4能夠與調控芥子油苷合成的一些MYB家族轉錄因子直接互作,形成轉錄因子復合體調控芥子油苷合成基因的表達;同時,MYC2、MYC3和MYC4也能夠結合到許多芥子油苷合成基因的啟動子序列上,直接調控這些基因的表達[28]。此外,MYC2還能夠直接與萜類化合物合成基因TPS10、TPS11和TPS21的啟動子結合并激活它們的表達,促進植物萜類化合物的合成[39],從而增強萜類化合物介導的植物對病蟲害的直接和間接抗性[6]。值得注意的是,不同于對害蟲抗性的調控作用,MYC2在植物對病原菌的抗性中起負調控作用,myc2突變體對灰霉菌(Botrytis cinerea)等病原菌的抗性強于野生型,這可能是由于MYC2與EIN3互作并抑制EIN3的功能,從而抑制了乙烯介導的植物對病原菌的抗性[38]。
茉莉素可顯著誘導植物花色素苷的積累和表皮毛的產生。花色素苷和表皮毛分別作為植物的一種化學和物理屏障,能有效提高植物對昆蟲、微生物等病蟲害的抗性,降低紫外線等非生物脅迫對植物造成的傷害[40-41]。擬南芥花色素苷的合成和表皮毛產生均受WD-repeat/bHLH/MYB復合體的調控[40,42]。JAZ 蛋白能夠與 WD-repeat/bHLH/MYB 復合體的bHLH組分(GL3、EGL3和TT8)和R2R3 MYB組分(MYB75、GL1)直接互作,抑制WD-repeat/bHLH/MYB復合體的活性,從而抑制植物花色素苷的積累和表皮毛的產生;茉莉素通過誘導JAZ蛋白降解,解除JAZ蛋白對WD-repeat/bHLH/MYB復合體的抑制作用,從而促進色素苷的積累和表皮毛的產生,增強植物對病蟲害和紫外等非生物脅迫的抗性[43]。此外,JAZ蛋白也可與YAB家族轉錄因子FIL互作調控色素的合成:FIL能夠結合到MYB75的啟動子上并激活MYB75的表達,從而促進色素合成;而JAZ蛋白通過與FIL互作,抑制FIL對MYB75的轉錄激活,從而抑制色素合成[44]。
IIId亞家族bHLH家族轉錄因子bHLH3、bHLH13、bHLH14和bHLH17作為轉錄抑制因子,抑制茉莉素響應基因的表達[45-48]。bhlh3 bhlh13 bhlh14 bhlh17四突變體表現出對茉莉素超敏的表型,且對昆蟲(甜菜夜蛾)和腐生真菌(灰霉菌)的抗性增強[46]。bHLH3、bHLH13、bHLH14 和 bHLH17雖不具有轉錄激活活性,但具有DNA結合能力,能夠通過拮抗MYC2、MYC3、MYC4等轉錄因子發揮轉錄抑制功能[45-48]。
JAV1是茉莉素途徑的一個負調控因子[15-16]。如前文所述,JAV1能夠與JAZ8和WRKY51形成JAV1-JAZ8-WRKY51復合體,抑制茉莉素合成基因的表達[15]。JAV1的突變體對灰霉菌和甜菜夜蛾的抗性強于野生型,而其超表達植株則對灰霉菌和甜菜夜蛾更敏感[16]。同樣,JAV1與WRKY51的雙突變體在傷誘導后會比野生型植物積累更多的茉莉素,其對甜菜夜蛾的抗性也明顯強于野生型植物[15]。植物抗性上的增強往往伴隨著生長上的抑制[49]。然而有趣的是,JAV1雖然在植物抗性中發揮重要作用,卻對植物生長發育沒有明顯影響[16]。
近期有研究表明,植物對茉莉素的響應水平以及對昆蟲的抗性受植物年齡影響,與幼嫩植物相比,成株植物對茉莉素的響應水平較低,但對昆蟲的抗性更強[50]。進一步研究發現,不同年齡植物對茉莉素的響應水平受miRNA156的靶標SPL9調控:SPL9能夠直接與JAZ蛋白互作,并抑制COI1介導的JAZ蛋白降解;隨著植物逐漸生長成熟,miR156水平逐漸降低,SPL9逐漸積累,使得JAZ蛋白含量逐漸升高,最終導致茉莉素響應水平逐漸降低[50]。值得注意的是,雖然成株植物的茉莉素響應水平低于幼嫩植物,但由于成株植物中積累了更多的芥子油苷等抗性物質,因此成株植物對昆蟲的抗性強于幼嫩植物[50]。
4 病原生物對植物茉莉素途徑的操控
茉莉素在調控植物對昆蟲和病原微生物的抗性反應中發揮重要作用。相應地,昆蟲、細菌、真菌、病毒及植原體等諸多植物病原生物都進化出了多種策略來操控茉莉素途徑,通過激活或抑制茉莉素途徑,促進自身的侵染或傳播(圖2)。
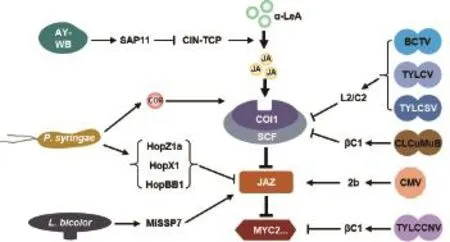
圖2 微生物對茉莉素途徑的操控策略
4.1 昆蟲對茉莉素途徑的操控
一些昆蟲通過激活水楊酸(SA)途徑間接抑制茉莉素途徑。甜菜夜蛾分泌的唾液含有葡萄糖氧化酶,能夠催化葡萄糖形成過氧化物,激活水楊酸途徑,從而間接抑制茉莉素途徑[51]。馬鈴薯甲蟲(Leptinotarsa decemlineata)通過其口器內的共生微生物激活水楊酸途徑,間接抑制茉莉素途徑,其對茉莉素途徑的抑制作用在經抗生素處理后消失,而經移植共生菌群后又得到恢復,表明了共生微生物在馬鈴薯甲蟲抑制茉莉素途徑中的關鍵作用[52]。
一些昆蟲利用茉莉素途徑下游ERF和MYC分支間的拮抗作用操控茉莉素途徑。菜青蟲(Pieris rapae)通過口器分泌物激活ERF分支,抑制MYC分支介導的植物對昆蟲的抗性,同時增強了ERF分支介導的植物對腐生菌的抗性,減輕其它病原生物對宿主植物侵害,以更多地從植物宿主攝取營養[53]。
4.2 細菌對茉莉素途徑的操控
一些細菌通過合成茉莉素類似物激活植物茉莉素途徑,以利于侵染。丁香假單胞菌(Pseudomonas syringae,簡稱P.syringae)能夠合成冠菌素(茉莉素類似物),冠菌素的結構與(+)-7-iso-JA-L-Ile較為相似,能夠被茉莉素受體COI1識別、誘導COI1與JAZ蛋白結合、激活茉莉素途徑[23-25](圖2)。冠菌素具有較強的茉莉素活性,與(+)-7-iso-JA-L-Ile的生物活性相當,比商業化生產的JA-Ile(4種構象的混合物)的生物活性高約100倍[10]。丁香假單胞菌利用冠菌素激活茉莉素途徑,從而促進植物氣孔打開,利于丁香假單胞菌從植物氣孔入侵;同時,由于茉莉素途徑與水楊酸途徑具有拮抗作用,冠菌素激活的茉莉素途徑干擾了水楊酸途徑,從而抑制了水楊酸途徑介導的植物抗性反應,利于丁香假單胞菌侵染[54-56]。
一些細菌利用自身分泌的效應蛋白激活植物茉莉素途徑,以利于侵染。丁香假單胞菌效應蛋白AvrB可通過與MPK4及分子伴侶HSP90/RAR1互作,促進MPK4的磷酸化,從而激活茉莉素途徑[57];此外,AvrB還可通過與RIN4互作激活AHA1,促進COI1與JAZ蛋白的互作,從而激活茉莉素途徑[58]。但AvrB通過RIN4/AHA1或MPK4/HSP90/RAR1激活茉莉素途徑的具體分子機制還不清楚。分別來源于丁香假單胞菌Psy株系和Pta株系的效應蛋白HopZ1a和HopX1都通過作用于植物JAZ蛋白來激活茉莉素途徑[59-60],但它們的作用機制有所不同。HopZ1a具有乙酰轉移酶活性,通過誘導JAZ蛋白的乙酰化,從而促進COI1依賴的JAZ蛋白的降解[59];而HopX1則利用其自身具有的半胱氨酸蛋白酶活性降解JAZ蛋白,此降解過程不依賴于COI1[60](圖2)。最近的研究表明,效應蛋白HopBB1也參與了丁香假單胞菌對植物茉莉素途徑的激活[61]。TCP14能夠抑制茉莉素響應基因的表達,而HopBB1通過與TCP14和JAZ3互作,促使TCP14被SCFCOI1泛素化降解,解除了TCP14對茉莉素響應基因的抑制作用;此外,HopBB1與JAZ3的互作也阻礙了JAZ3對MYC2的抑制作用,這些機制都使得HopBB1能夠激活茉莉素途徑[61](圖 2)。
4.3 真菌對茉莉素途徑的操控
腐生型病原真菌或共生真菌往往通過抑制茉莉素途徑、降低茉莉素介導的植物抗性,利于腐生型病原真菌的侵染或共生真菌與植物的共生。灰霉菌產生的一種胞外多糖 β-(1,3)(1,6)-D-glucan能夠誘導植物體內水楊酸的合成,利用水楊酸-茉莉素途徑間的拮抗,抑制茉莉素介導的植物抗性[62]。核盤菌(Sclerotinia sclerotiorum)編碼的效應蛋白SSITL通過抑制茉莉素和乙烯途徑促進侵染,但SSITL抑制茉莉素和乙烯途徑的具體分子機制還不清楚[63]。楊樹共生真菌雙色蠟蘑(Laccaria bicolor,簡稱L.bicolor)編碼的效應蛋白MiSSP7能夠與楊樹的JAZ6蛋白互作并抑制JAZ6蛋白的降解,從而抑制楊樹茉莉素信號通路以促進雙色蠟蘑與楊樹的共生[64](圖 2)。
也有一些真菌通過激活茉莉素途徑促進侵染。尖孢鐮刀菌(Fusarium oxysporum)、漆蠟蘑(Laccaria laccata)、黑曲霉(Aspergillus niger)等真菌都已被發現能夠合成JA、JA-Ile等茉莉素類物質[65]。尖孢鐮刀菌Fo5176株系還可通過其效應蛋白SIX4激活植物茉莉素途徑進而促進侵染[66]。值得一提的是,這些激活茉莉素途徑的真菌并不都是通過間接抑制SA途徑促進侵染的。例如,coi1突變體對尖孢鐮刀菌的抗性強于野生型植物,說明茉莉素途徑促進尖孢鐮刀菌的侵染,但尖孢鐮刀菌在coi1單突變體和coi1/NahG(過量表達SA羥基酶而不能積累SA)雙突變體上的侵染能力相近,說明尖孢鐮刀菌并不是利用JA-SA之間的拮抗關系來促進侵染的[67]。此外,SIX4并不顯著影響植物SA途徑,進一步映證了尖孢鐮刀菌并不是通過操控JA途徑間接影響SA途徑來促進侵染的[66]。這些研究暗示,茉莉素介導的植物與真菌間的互作可能比我們預想的更加復雜。
4.4 病毒和植原體對茉莉素途徑的操控
一些以昆蟲為介體進行傳播的病毒和植原體,通過抑制植物茉莉素途徑,抑制植物對介體昆蟲的抗性或提高植物對介體昆蟲的吸引性,從而促進自身的傳播。翠菊黃化植原體AY-WB株系(Aster Yellows phytoplasma strain Witches' Broom,AY-WB)分泌的效應蛋白SAP11通過與CIN-TCP轉錄因子互作并促進CIN-TCP的降解,抑制CIN-TCP對茉莉素合成基因(LOX2等)的轉錄激活,從而抑制茉莉素的合成,進而抑制茉莉素介導的植物對其傳播介體翠菊葉蟬的抗性,以促進自身的傳播[68](圖2)。黃瓜花葉病毒(Cucumber mosaic virus,CMV)能夠顯著干擾植物茉莉素途徑,進而影響宿主植物與介體昆蟲間的互作[69-70]。最近的研究表明,CMV編碼的2b蛋白能夠與植物JAZ蛋白直接互作,抑制SCFCOI1介導的JAZ蛋白降解,從而抑制茉莉素途徑,增強植物對病毒傳播媒介蚜蟲的吸引力,以利于病毒傳播[71](圖2)。番茄黃曲葉病毒(Tomato yellow leaf curl China virus,TYLCCNV)編碼的 βC1蛋白通過與植物AS1和MYC2蛋白互作,抑制植物茉莉素途徑,降低植物對其介體昆蟲煙粉虱的抗性[72-74](圖 2)。
一些植物病毒通過抑制茉莉素途徑促進侵染。水稻齒葉矮縮病毒(Rice ragged stunt virus,RRSV)的侵染能夠誘導miR319的積累,降低miR319靶標基因TCP21的表達,削弱了TCP21對茉莉素合成基因的激活作用,抑制了茉莉素的合成和茉莉素響應基因的表達,從而抑制了茉莉素介導的植物對RRSV的抗性,利于RRSV的侵染[75]。甜菜曲頂病毒(Beet curly top virus,BCTV)編碼的L2蛋白及其在番茄黃化曲葉病毒(Tomato yellow leaf curl virus,TYLCV)和番茄黃葉卷撒丁病毒(Tomato yellow leaf curl Sardinia virus,TYLCSV)中的同源蛋白C2,通過與植物COP9復合體中的CSN5亞基互作,影響SCF復合體組分CUL1的去RUB化修飾,從而影響了包括SCFCOI1在內的E3泛素連接酶的活性,進而抑制了茉莉素途徑介導的植物病毒抗性,促進了病毒侵染[76]。木爾坦棉花曲葉病毒(Cotton leaf curl Multan virus,CLCuMuB)編碼的βC1蛋白通過與煙草SKP1蛋白互作,抑制SCFCOI1復合體的形成,從而抑制茉莉素途徑,促進病毒侵染[77](圖2)。
值得注意的是,許多操控植物茉莉素途徑的病毒效應蛋白(包括CMV編碼的2b蛋白、TYLCCNV編碼的βC1蛋白、BCTV編碼的L2、CLCuMuB編碼的βC1蛋白等)同時也是RNA沉默抑制子(Viral suppressor of RNA silencing,VSR)[78-81],暗示病毒VSR蛋白功能的多重性,即既通過抑制植物RNA沉默促進病毒在植物內的增殖,又通過抑制茉莉素途徑促進病毒在植物內的增殖或在植物間的傳播。
5 展望
植物固定生長不能移動,在長期的進化過程中,植物形成了獨特而復雜的防御機制來抵抗昆蟲和微生物的入侵;與此同時,昆蟲和微生物也進化出了各種機制來逃避、克服或利用植物的防御系統,以實現對宿主植物的寄生。茉莉素在調控植物對昆蟲和微生物的抗性反應中發揮重要作用。相應地,包括昆蟲、真菌、細菌、病毒在內的許多生物都通過多種策略操控茉莉素途徑,直接或間接影響植物抗性,從而促進侵染或傳播。近年來,茉莉素信號轉導機制的逐漸明晰和部分重要植物病原全基因組測序的完成,為系統和深入研究茉莉素介導的植物與病原生物互作提供了極大便利,相關研究領域也取得了很大進展。
已有的研究表明,茉莉素在調控植物對病毒的抗性方面發揮一定作用,但對于不同的植物病毒,茉莉素的調控作用卻不盡相同。例如,茉莉素正調控植物對RRSV(雙鏈RNA病毒)、CLCuMuB(DNA病毒)和BCTV(DNA病毒)的抗性[75-77],卻負調控植物對煙草花葉病毒(單鏈RNA病毒)的抗性[82]。茉莉素究竟如何調控植物對病毒的抗性還有待進一步研究。此外,茉莉素在調控植物對腐生型真菌的抗性反應中發揮關鍵作用,然而與細菌、病毒等病原微生物相比,腐生型真菌操控茉莉素途徑的研究還很少見,作用于茉莉素途徑的腐生型真菌效應蛋白還有待更加廣泛地鑒定和研究。對這些重要科學問題的探索,將使人們對茉莉素介導的植物與病原生物的互作有更加全面和深入的認識,從而為抗病蟲害植物分子育種奠定理論基礎。
猜你喜歡
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
中國衛生(2016年3期)2016-11-12 13:23:26
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
