擬南芥熱形態建成中PIF4下游基因研究
2018-08-18 01:44:14汪碩丁嵐劉建祥韓佳嘉
生物技術通報 2018年7期
關鍵詞:植物
汪碩 丁嵐 劉建祥,2 韓佳嘉,2
(1. 復旦大學生命科學學院,上海 200433;2. 浙江大學生命科學學院,杭州 310027)
隨著溫室效應的加劇,近些年全球氣候逐漸變暖,20世紀全球平均氣溫約提高0.6℃[1],經預測,到2050年平均溫度將可能提高2-5℃[2]。高溫會對植物的生長發育產生直接影響,在較高環境溫度下植物會出現葉片偏下、開花提前及下胚軸伸長等表型,與光形態建成對應,植物在環境溫度升高條件下的一些形態上的改變被稱為熱形態建成(thermomorphogenesis)。現有研究表明,調控這些表型的信號途徑與植物遮蔭下的分子信號途徑多有重疊,表型也十分類似[3]。
光和環境溫度作為最重要的環境信號,極大地影響了植物的生長發育。光敏色素互作因子(Phytochrome interacting factors,PIFs),是一類可以與光受體光敏色素蛋白相互作用的bHLH家族轉錄因子。PIFs在光調控的植物生長發育過程中十分重要,能夠影響植物的下胚軸伸長、幼苗的暗形態建成、促進避蔭反應等[4]。29℃的溫和高溫(mild high temperature)能誘導擬南芥野生型幼苗下胚軸伸長,而PIF4缺失突變體pif4對溫和高溫不敏感,下胚軸幾乎不伸長[4]。隱花素(cryptochrome,CRY)是光裂解酶類似的藍光受體,在擬南芥苗期主要調控其下胚軸伸長。CRY1對溫度響應十分重要,而溫和高溫下CRY1通過與PIF4以藍光依賴的形式相結合,進而調控PIF4在溫和高溫下的轉錄活性[5]。最近,有報道指出光敏色素B(phyB)不僅可作為光的受體,同時能感應環境溫度的變化[6],但溫和高溫下phyB到PIF4的信號傳遞目前不是很清楚。
另外一條獨立于PIF4的調控植物響應光和溫度信號,控制下胚軸伸長的信號通路包含COP1(CONSTITUTIVE PHOTOMORPHOGENIC 1)和HY5(ELONGATED HYPOCOTYL 5)。HY5是 bZIP家 族轉錄因子,不但在白光下抑制擬南芥下胚軸伸長,同時在常溫下也能通過與PIF4競爭下游靶標基因啟動子來抑制下胚軸伸長[7]。在黑暗條件下或環境溫度升高后,HY5被泛素連接酶COP1降解,從而解除了HY5 對下胚軸伸長的抑制效果[7-10]。COP1在細胞核中發揮作用,但溫和高溫下COP1如何進核的分子機理目前不清楚。
溫度的升高會使生長素表達水平上升,進而調節下胚軸伸長等植物生長活動[11]。有研究表明當溫度升高時,擬南芥中生長素響應基因表達水平也有所變化[12]。擬南芥在響應溫和高溫時,PIF4能夠直接結合生長素合成基因,如YUC8,TAA1,CYP79B2等基因的啟動子。PIF4通過激活TAA1和CYP79B2基因的表達,進而介導吲哚乙酸(IAA)的合成[13]。而IAA的積累能夠促進AUX/IAA蛋白的降解,進而消除對ARF蛋白的抑制作用。最后ARF通過激活SAUR19等基因的表達促進胚軸伸長[14]。其它植物激素,例如,油菜素內脂BR和赤霉素GA在植物熱形態建成中也發揮著重要的作用[12,15-17]。
本實驗利用擬南芥PIF4缺失突變體和PIF4過表達等材料開展研究。遺傳分析表明,PIF4、PIF5和PIF7中,PIF4在熱形態建成中的作用最重要。通過對RNA-Seq結果和結合CHIP-Seq分析找到了擬南芥熱形態建成中PIF4調控的下游基因以及結合靶標基因。
1 材料與方法
1.1 材料
實驗所使用的植物材料有擬南芥(Arabidopsis thaliana)Columbia生 態 型(Col-0), 擬 南 芥Columbia背景突變體pif4-101,擬南芥野生型背景下PIF4∶PIF4-myc轉基因植株,擬南芥pif4-101背景下PIF4∶PIF4,PIF4∶PIF5,PIF4∶PIF7轉基因植株。所使用的菌株有大腸桿菌(Escherichia coli)TOP10, 農 桿 菌 菌 株(Agrobacterium tumefaciens)GV3101。這些材料均為本實驗室獲得和保存。
1.2 方法
1.2.1 擬南芥的平皿培養 將同批收獲的種子放于1.5 mL離心管中,用1 mL 體積比為50%的酒精迅速清洗1次,用體積比0.3%的NaClO消毒15 min后,于超凈臺內用無菌水清洗3次,播種在含質量濃度1.2%蔗糖的1/2MS固體培養基上。4℃春化2-3 d后置于22℃光照培養箱,光16 h/暗8 h。
1.2.2 下胚軸長度測量和分析 將相應材料于22℃培養4 d后,在第5天光照開始時移至29℃培養4 d,以22℃培養的植物作為對照。用相機分別拍下對應天數的不同材料的擬南芥幼苗生長照片,利用ImageJ軟件進行數據測量分析。
1.2.3 RNA-Seq 擬南芥Columbia野生型(wild-type,WT)和pif4-101突變體4℃春化2 d后于22℃培養7 d。在光照開始時將幼苗移至29℃處理1 d,以22℃培養的植物作為對照。于黑暗周期結束后收集擬南芥幼苗用于RNA-Seq。根據Illumina標準步驟構建cDNA文庫,Illumian Hiseq3000測序(晶能公司)。GO 分析采用 AgriGO(v2.0)進行分析[18]。
1.2.4 染色質免疫共沉淀-定量PCR(ChIP-qPCR)分別將擬南芥Columbia野生型及PIF4∶PIF4-myc轉基因植株于22℃培養,第12天29℃處理24 h,以22℃培養的植物作為對照。用甲醛固定液固定,后用2.5 mol/L甘氨酸解交聯,MilliQ水清洗材料3次。去除水分,凍于-80℃保存。將材料用液氮研磨成粉末,加入30 mL EB1,用2層Miracloth過濾。離心后用1 mL EB2重懸,離心并用300 μL EB3重懸沉淀,并覆蓋于600 μL EB3上。離心并用300 μL NLB重懸沉淀,使用biorupt進行超聲,超30 s停30 s。離心取部分上清做Input。其余每個重復加入50 μL protein A beads去除與beads的非特異性結合。取上清加入2 μg myc抗體,于4℃旋轉過夜。次日加protein A beads 110 μL,4℃ 2 h 后低速離心留 beads。分別用低鹽、高鹽和LiCl和TE緩沖液清洗beads。分兩次加入300 μL 洗脫緩沖液65℃煮15 min,合并。用洗脫緩沖液將Input補齊到500 μL,與免疫沉淀后的DNA-蛋白復合物一起于65℃解交聯約6 h。用蛋白酶K去除蛋白,RnaseI去除RNA。沉淀DNA,并用無水乙醇及70%乙醇清洗沉淀,晾干后用無菌水溶解。
以MYC抗體沉淀的DNA為模板,進行qPCR擴增,以actin啟動子序列作為陰性對照。qPCR實驗 使 用 SYBR Premix Ex TaqTMII(TaKaRa) 試 劑,在Bio-Rad CFX96TM儀器上完成。反應體系按照TaKaRa及Bio-Rad CFX96TM儀器配制。得到數據后分析處理并作圖。
1.2.5 實時熒光定量PCR實驗(qRT-PCR) RNA提取使用植物總RNA提取試劑盒(TIANGEN),總RNA保存于-80℃。cDNA使用 M-MLV(TaKaRa)反轉錄得到,保存在-20℃。
qRT-PCR實驗使用SYBR Premix Ex TaqTMII(TaKaRa)試劑,在Bio-Rad CFX96TM儀器上完成。反應體系按照TaKaRa及Bio-Rad CFX96TM儀器配制。得到數據后分析處理并作圖。
2 結果
2.1 PIF4在擬南芥熱形態建成中的主要生物學功能
本實驗在擬南芥PIF4突變體pif4-101背景下采用PIF4自身啟動子分別回補PIF4、PIF5和PIF7,利用這3種互補遺傳材料來研究PIF4及同源蛋白在擬南芥熱形態建成中的生物學功能的區別。野生型擬南芥和pif4-101、PIF4∶PIF5、PIF4∶PIF7在22℃生長8 d后,4種材料的幼苗下胚軸長度基本一致(圖1-A),而PIF4∶PIF4的下胚軸則明顯長于其他材料。而在22℃生長4 d、29℃生長4 d的條件下,PIF4∶PIF4的下胚軸伸長更長,野生型和PIF4∶PIF5的下胚軸比22℃時伸長的幅度接近,而pif4-101、PIF4∶PIF7則幾乎不伸長(圖1-B)。
基因表表達分析(圖1-C)表明,PIF4∶PIF4植物中正常條件下PIF4表達量比野生型中高,可能是PIF4∶PIF4植物正常溫度條件下下胚軸比較長的原因。這些結果表明,擬南芥熱形態建成中PIF7的功能較小,PIF5也具有PIF4類似的功能,但PIF4的功能明顯比PIF5的功能強。因此,PIF4在擬南芥熱形態建成中具有重要的功能。
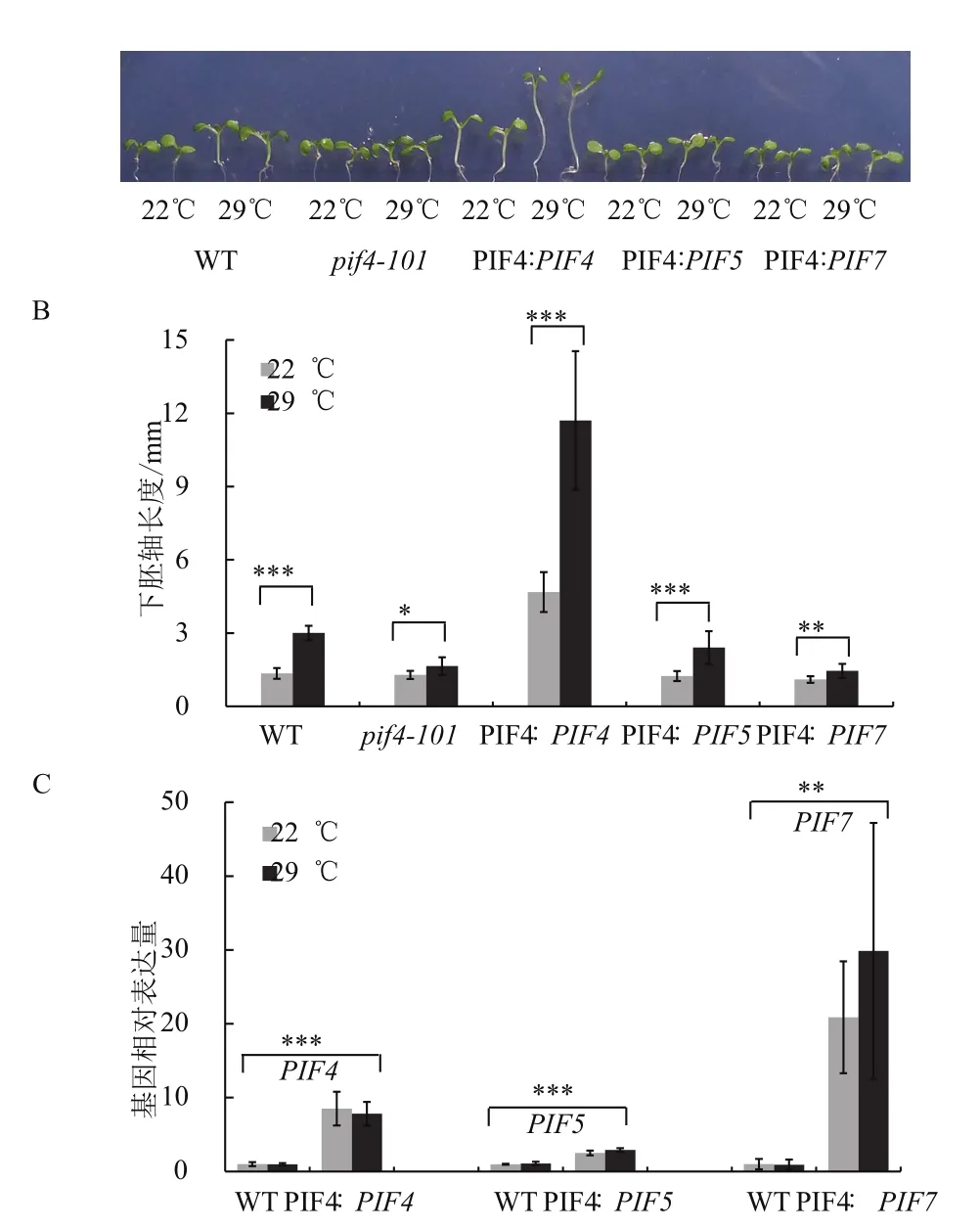
圖1 PIF4突變體的表型遺傳互補分析
2.2 熱形態建成中PIF4調控的下游基因
為了在全基因組水平上理解熱形態建成過程中PIF4調控的下游基因表達,將擬南芥野生型和突變體pif4-101在22℃ 培養7 d后,于光周期中白光開始時用29℃處理24 h,在黑暗結束后取樣,以不處理的植物作對照,收集幼苗全苗進行轉錄組測序(RNA-Seq)。對轉錄組數據進行統計分析后顯示,在擬南芥野生型與pif4-101中分別有602個基因和512個基因受溫和高溫處理上調,并有529個基因和484個基因受溫和高溫處理下調。其中有248個基因只在野生型中特異上調,198個基因只在野生型中特異下調(圖2,附表)。
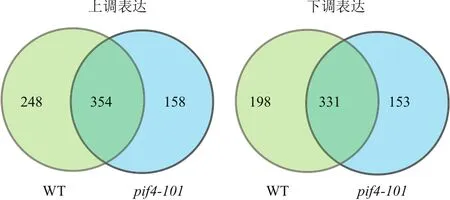
圖2 PIF4突變體與野生型熱處理的RNA-SEQ分析
對于這些只在野生型中上調的248基因,我們認為它們是PIF4依賴的熱響應基因。GO分析(圖3)顯示,PIF4依賴的熱響應基因中主要參與脫落酸響應、生長素響應、幾丁質響應、脫水響應、鹽脅迫、冷馴化等過程。綜上所述,這些結果表明,PIF4依賴的熱響應基因主要參與植物激素信號通路和逆境應答。
對于354個在野生型和突變體pif4-101中上調的下游基因,對其上調幅度也進行了比較。結果(表1)顯示,有21個基因在pif4-101突變體中的上調倍數是野生型(WT)中上調倍數的1/2及以下,我們把這些基因視作是部分依賴PIF4調控的熱形態建成基因。
2.3 熱形態建成中PIF4調控的結合靶標基因
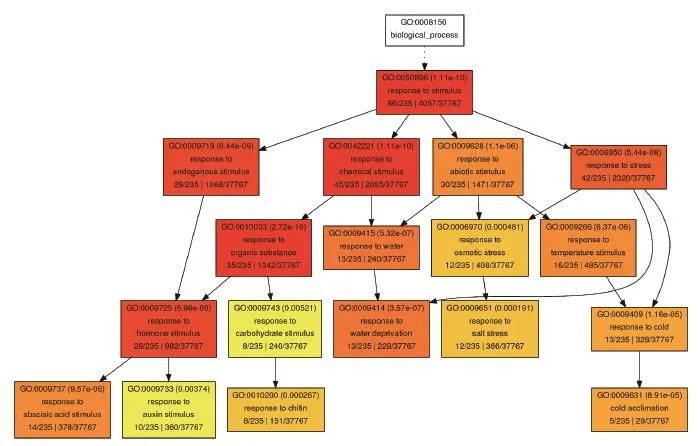
圖3 PIF4依賴的熱響應基因GO分析

表1部分依賴PIF4調控的熱形態建成基因
為了檢測上述PIF4依賴和部分依賴的基因是否是PIF4的直接靶標基因,我們在擬南芥野生型背景下過表達myc標簽融合的PIF4-myc材料(PIFox)。野生型WT和PIF4ox-5、PIF4ox-8轉基因材料在22℃正常生長8 d后,PIF4過表達轉基因材料表現出下胚軸變長的表型。在22℃生長4 d后移入29℃培養4 d,野生型的下胚軸出現一定程度的伸長,PIF4過表達轉基因材料下胚軸伸長現象更加明顯(圖4)。這表明PIF4過表達材料正常溫度下可能已經激活部分下游基因,并且能響應環境溫度升高的變化,這些材料可以用于后續結合靶標基因的實驗。
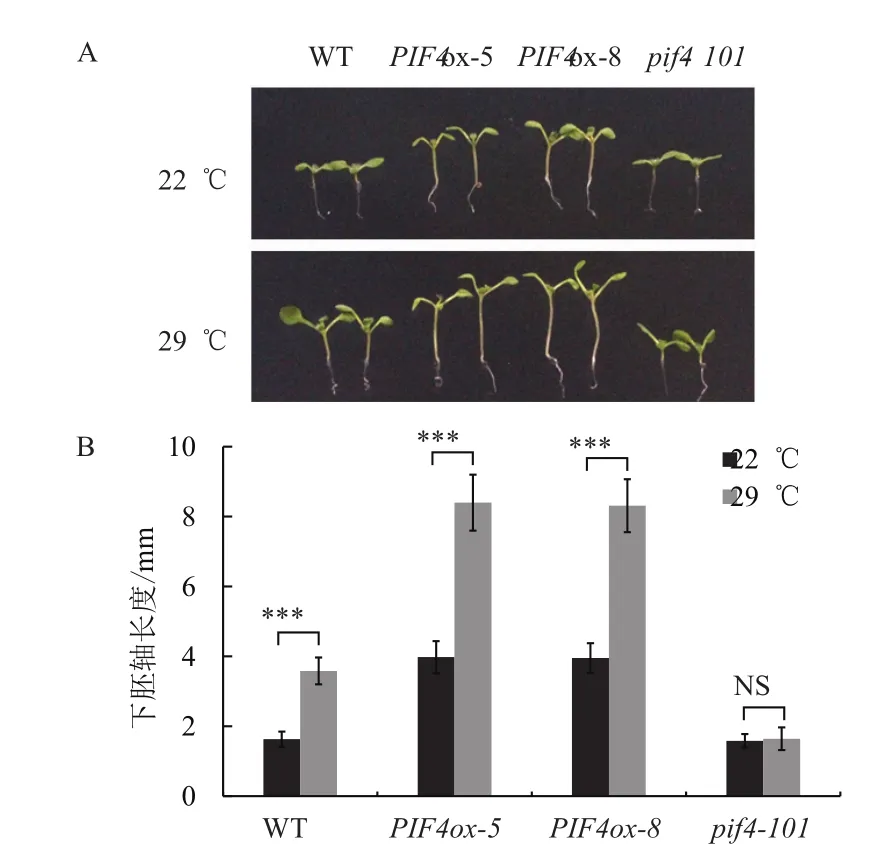
圖4 PIF4過表達的表型分析
在研究植物激素BR調控下胚軸伸長過程時,王志勇課題組采用pifq四突變體背景下自身啟動子驅動的PIF4-myc過表達材料,正常生長條件下進行染色質共免疫沉淀深度測序(ChIP-Seq)分析,得到4583個候選的PIF4結合靶標基因[15-16]。
為了研究熱形態建成中PIF4的結合靶標基因,我們比較了上述248個PIF4依賴的基因以及PIF4候選的結合靶標基因,得到70個熱形態建成過程中被PIF4結合的候選靶標基因。
我們隨機選擇了7個靶標基因,包括AT1G29395(COR413IM1)、AT1G73120(F-box 蛋白 )、AT2G47780(LDAP2)、AT3G28857(PRE5)、AT4G14130(XTR7)、AT5G07010(ATST2A)、AT5G18050(SAUR22),利用PIF4-myc過表達材料(PIFox-5)進行ChIP-qPCR實驗。結果(圖5)表明,AT1G29395、AT3G28857、AT5G18050這 3個基因的啟動子區域在正常溫度下就可以被PIF4-myc富集(≥1.5倍,P < 0.05),在環境溫度升高后7個候選靶標基因中,6個基因(AT1G29395、AT1G73120、AT2G47780、AT3G28857、AT5G07010、AT5G18050)的啟動子區域被PIF4-myc富集(≥1.5倍,P < 0.05),說明通過生物信息分析得到的70個PIF4結合靶標基因中絕大部分為實際PIF4結合靶標基因(附表)。

圖5 PIF4體內結合下游基因啟動子的分析
我們也比較了上述21個PIF4部分依賴的基因以及PIF4候選的結合靶標基因,得到4個PIF4熱形態建成過程中被PIF4結合的靶標基因(表1)。我們把這74個PIF4結合靶標基因視作熱形態建成中PIF4結合的靶標基因,這些靶標基因是否被PIF4直接結合可以采用凝膠遷移EMSA實驗進行證明。
2.4 PIF4過表達對下游基因的影響
為了研究PIF4過表達對下游基因表達的影響,將野生型(WT)和PIF4-myc過表達材料(PIF4ox-5)進行不同溫度處理,檢測以上提到的7個基因的表達情況。結果(圖6)表明,6個 基 因(AT1G29395、AT1G73120、AT2G47780、AT3G28857、AT4G14130、AT5G18050)正常溫度下PIF4ox-5中的表達量高于野生型中的表達量,只有1個基因(AT5G07010)在溫和高溫處理后PIF4ox-5中表達量才高于野生型中的表達量。
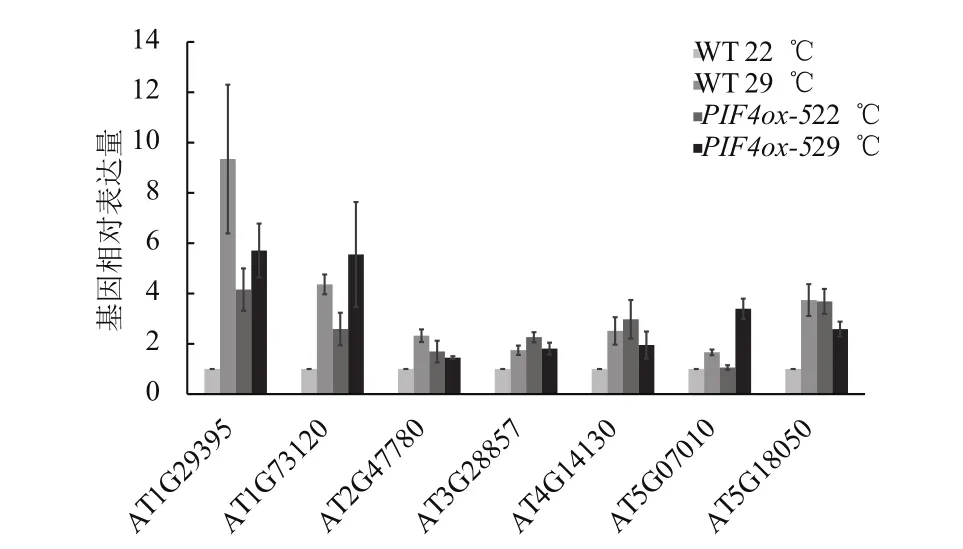
圖6 PIF4過表達對下游基因表達的影響
3 討論
bHLH家族轉錄因子PIF4與關敏色素phyB互作,參與植物光形態建成,后續更多的研究表明,PIF4是植物熱形態建成中的重要調控節點[19-23]。我們的實驗結果也表明,在幾個PIFs中,PIF4在植物熱形態建成中功能最重要。PIF4基因的表達受時鐘的調控,同時也受溫和高溫的誘導[20,24]。PIF4蛋白也受到翻譯后修飾調控。例如,在白光下phyB和BIN2可以磷酸化PIF4,導致PIF4的降解[25-26]。環境溫度升高后磷酸化的PIF4積累[27],這主要與DET1穩定PIF4有關[7]。此外,PAR1和CRY1也可以與PIF4互作,溫和高溫下抑制PIF4 的轉錄活性[5,28-29]。
PIF4的靶標基因中有很多是植物激素合成和響應基因[12,15],因此一般認為PIF4在熱形態建成中主要通過植物激素,如生長素途徑發揮作用[11,23]。事實上,生長素合成基因比如YUC8和生長素響應基因比如IAA4 和IAA29,受溫和高溫的誘導上調并且依賴于PIF4[13,20]。但是,PIF4在全基因組水平的熱形態建成中的下游基因并未見報道。我們采用RNA-Seq的方法,得到了溫和高溫處理后受PIF4調控的269個下游基因(包括21個PIF4部分依賴的下游基因)。這些基因中生長素響應途徑的基因(IAA6、IAA19、IAA34、SAUR22、SAUR57、SAUR59)和脫落酸響應途徑的基因得到顯著富集。此外,一些細胞壁伸長相關基因和逆境脅迫響應基因也得到富集。我們的結果為理解植物熱形態建成的基因調控提供了幫助。
這些PIF4調控的熱形態建成下游基因中,據估算大約有25%為熱形態建成中PIF4調控的結合靶標基因,其他為PIF4間接調控的下游基因。例如,編碼膜蛋白COR413IM1的基因AT1G29395被證明是PIF4的直接調控基因。溫和高溫處理下的該基因的誘導表達依賴于PIF4,野生型中組成性表達PIF4正常溫度下該基因上調表達。而編碼細胞壁成分、改變有關的木葡聚糖內糖基轉移酶XTR7的基因AT4G14130并不被PIF4直接結合,是PIF4間接調控的下游基因。溫和高溫處理下該基因的誘導表達也依賴于PIF4,野生型中組成性表達PIF4正常溫度下該基因也上調表達。
此外,PIF4對下游靶標基因的結合能力也有差別,當PIF4組成性表達后,有的靶標基因在正常溫度下就可以被PIF4結合,而有些靶標基因只有在溫和溫度處理后才被PIF4結合。例如,編碼磺基轉移酶ATST2A的AT5G07010啟動子只有在溫和高溫處理下才顯著被PIF4結合。溫和高溫處理下該基因的誘導表達依賴于PIF4,野生型中組成性表達PIF4,正常溫度下該基因沒有上調表達,但溫和高溫處理后該基因明顯誘導表達。這些結果暗示其他的溫和高溫處理依賴的轉錄調控因子可能與PIF4一起調控了該基因的表達。PIF4過表達植物正常條件下下胚軸伸長,溫和高溫下有進一步的伸長,而在檢測的7個基因中,只有1個基因(ATST2A)的表達量在PIF4過表達植物中溫和高溫下有進一步上調表達,說明一部分PIF4調控基因在溫和高溫下對下胚軸的伸長非常重要。
4 結論
獲得擬南芥熱形態建成中248個PIF4完全依賴和21個PIF4部分依賴的下游基因,這些下游基因主要參與植物激素和非生物逆境響應。這些PIF4調控的熱形態建成下游基因有74個基因是PIF4結合靶標候選基因。選擇7個PIF4候選靶標基因,有6個基因被PIF4直接結合。
注:本論文中附表見電子版(http://biotech.caas.cn)
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
