浙江省極小種群多脈鐵木年齡結構與動態
2018-08-24 08:57:10吳世斌庫偉鵬周小榮周海偉紀美芬吳家森
浙江農業科學 2018年8期
吳世斌,庫偉鵬,周小榮,周海偉,紀美芬,吳家森*
(1.文成縣林業局,浙江 文成 325300; 2.浙江農林大學 環境與資源學院,浙江 杭州 311300; 3.文成縣苗圃,浙江 文成 325300; 4.文成縣石垟林場,浙江 文成 325300; 5.文成縣葉勝林場,浙江 文成 325300)
植物種群結構是指種群內不同年齡、大小個體的分布狀況,反映了各齡級個體的組配、立地條件和環境間的相互關系[1]。生命表和存活曲線能直觀地展現不同齡級個體的現有的生存數、死亡數[2-3],分析過去的種群結構和受干擾情況,預測未來的種群變化,了解種群對環境條件的適應性[4],對于瀕危植物的保護和管理具有重要意義。
多脈鐵木(Ostryamultinervis)為樺木科鐵木屬植物,分布于湖北利川、湖南紫云山、四川東南部和貴州等地,其分布區孤立局限,具有樹體高大,干形通直,材質優良的特點[5],為我國特有種植物,是浙江省珍稀瀕危野生植物和極小種群保護對象。已有研究對多脈鐵木的繁育技術、林分特征進行了初步研究[6-7],而有關多脈鐵木種群的年齡結構及數量動態的研究未見報道。本文詳細調查了浙江省文成縣石垟林場多脈鐵木群落的所有個體,利用徑級結構分析,編制了種屬靜態生命表和種群存活曲線,預測種群時間序列的變化趨勢,旨在為多脈鐵木的種群恢復策略的制定提供理論和參考價值。
1 研究地區與方法
1.1 研究區概況
研究區位于浙江省文成縣石垟林場猴王谷景區倒臼源,多脈鐵木的分布中心地理位置為27°50′N, 119°50′E。研究地所在的石垟林場屬中亞熱帶季風性氣候,年平均氣溫12.8 ℃,最冷月(1月)平均氣溫為4.0 ℃,極端最低氣溫-14.0 ℃,最熱月(7月)平均氣溫為23.6 ℃,極端最高氣溫37.3 ℃,年降水量1 604 mm,年均相對濕度84.0%,年日照時數1 755 h,全年無霜期288 d[8]。該區域屬天然常綠闊葉林,森林覆蓋率達90%以上。多脈鐵木群落分布區域的海拔介于500~700 m,坡度30°~35°,南坡,坡中部,地表有石塊層覆蓋,厚度20~30 cm,石塊大小20~40 cm。土壤為發育于花崗巖的紅壤土類,表層土壤(0~20 cm)理化性質為pH值4.6,有機碳63.2 g·kg-1,堿解氮、有效磷、速效鉀含量分別為145.2、21.82、187 mg·kg-1。
多脈鐵木種群中植物資源豐富,共有維管植物60種(含變種),隸屬于40科52屬。伴生的喬木樹種有8種,分別是褐葉青岡、小葉櫟、四照花、缺萼楓香、短尾柯、鉤栲、尾葉冬青、麂角杜鵑;灌木層主要植物有披針葉茴香、毛花連蕊茶、光葉石楠、長裂葛蘿槭、紅楠、三尖杉、猴歡喜、大葉冬青、秀麗槭、青榨槭;草本層植物主要有里白、黑足鱗毛蕨、麥冬、求米草、三穗薹草、瘤足蕨、江南卷柏、毛葉腹水草、大葉唐松草、延羽卵果蕨。
1.2 研究方法
1.2.1 調查
2017年8月上旬,全面調查種群內所有的多脈鐵木個體,對其所有個體進行每株檢尺(起測徑階≥5.0 cm),測定樹高、胸徑、冠幅,記錄<5.0 cm的幼苗數量,并訪問林場老職工,了解該種群分布歷史及利用情況。
1.2.2 徑階結構劃分
根據多脈鐵木的生物學特性和野外調查數據的特點,將多脈鐵木種群劃分為10個徑階,每個徑階對應相應的齡級。徑階<5 cm為第Ⅰ齡級,5~<10 cm為第Ⅱ齡級,10~<15 cm為第Ⅲ齡級,15~<20 cm為第Ⅳ齡級,20~<25 cm為第Ⅴ齡級,25~<30 cm為第Ⅵ齡級,30~<35 cm為第Ⅶ齡級,35~< 40 cm為第Ⅷ齡級,40~<45 cm為第Ⅸ齡級,D≥45 cm為第Ⅹ齡級。
1.2.3 數據分析
多脈鐵木種群的年齡結構、靜態生命表及存活曲線、動態量值及指數、時間序列預測等指標參考申仕康等[1]的方法進行。
2 結果與分析
2.1 多脈鐵木種群的年齡結構
調查研究表明,多脈鐵木種群在其全分布范圍內共找到118株,其年齡結構如圖1所示。從圖中可知,Ⅰ齡級的幼苗有13株,占總數的11.0%,而Ⅱ齡級的幼樹僅1株,而后隨著年齡的增大,植株個體數量先增加而后下降,多數集中于Ⅴ~Ⅶ齡級,占整個種群的52.%。多脈鐵木的年齡結構近似正態分布的特點,種群暫時穩定。但Ⅱ齡級個體數量急驟減少,表明該種群在演替過程中受到嚴酷的環境篩影響,導致Ⅰ齡級向Ⅱ齡級過渡中死亡率達92.3%,嚴重影響著種群的發展。

圖1 多脈鐵木的齡級結構
2.2 多脈鐵木種群的靜態生命表及存活曲線
從靜態生命表(表1)和存活曲線(圖2)中可知,多脈鐵木幼苗個體數量較多,種群存活曲線趨于Deevey-Ⅲ型,多脈鐵木個體由第Ⅰ齡級向第Ⅱ齡級演替轉化的過程中,其數量顯著下降,僅有7.7%的個體能進入到第Ⅱ齡級生長,這表明幼年個體向成年階段的發育是不連續的,該時間已成為多脈鐵木種群發展的“瓶頸”。多脈鐵木Ⅲ~Ⅶ齡級的個體數目先增加而后保持穩定,到第Ⅷ齡級后,其個體數量又呈現減少趨勢。

表1 多脈鐵木種群的靜態生命

圖2 多脈鐵木種群存活曲線

圖3 多脈鐵木種群死亡率(Qx)和損失度(Kx)曲線
多脈鐵木種群死亡率(Qx)和損失度(Kx)的變化趨勢相似(圖3),Kx在第Ⅰ齡級和第Ⅷ齡級出現峰值,第Ⅰ齡級的峰值是因為幼苗以高損失率為代價向第Ⅱ齡級轉換,這與存活曲線的變化規律是一致的;第Ⅷ齡級的峰值是由于存活個體的生理衰老退化,從而形成損失高峰。多脈鐵木Ⅲ~Ⅶ齡級的Qx和Kx接近于0,表明該年齡段其種群結構穩定。
2.3 多脈鐵木種群的動態量值

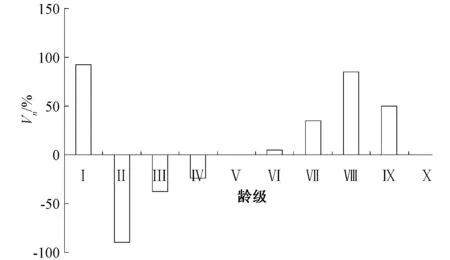
圖4 多脈鐵木種群的動態量值

圖5 多脈鐵木種群動態指數
2.4 多脈鐵木種群的時間序列預測
多脈鐵木的時間序列預測(表2)表明,從2019年起,在之后的2 a內,多脈鐵木種群Ⅱ齡級以上的個體數量呈現增長趨勢,而5 a后,該物種僅在Ⅴ齡級后才有個體增加,表現為低齡級的個體數量不足,種群面臨衰退。

表2 多脈鐵木齡級結構的時間序列預測
3 小結與討論
某一植物種群的年齡結構是其生物學特性與環境因子共同作用的結果[9],年齡金字塔呈紡錘形的種群為穩定的群落結構[10]。本研究結果表明,多脈鐵木種群從第Ⅱ齡級開始的年齡結構金字塔基本呈紡錘形,近似于正態分布,即中齡個體數量占種群總體數量的比例較高,而幼齡和老齡個體較少,表現為暫時的穩定結構特征。盡管Ⅰ齡級的多脈鐵木幼苗數量較多,但也僅13株,而Ⅱ齡級的幼樹數量僅1株,出現明顯的瓶頸現象,受到環境篩的顯著影響。種子產量低、品質差,種子向幼苗的轉化率低,形成的幼苗數量少是瀕危植物存在的較普遍問題[11]。多脈鐵木種子的種皮堅硬、透水性差,吸水主要通過苞片網脈,種子不育、空癟粒多,種子飽滿率僅12%。因此種子品質差是造成多脈鐵木幼苗少的內在因素;其次,多脈鐵木種群所在的立地環境中,表層有大量石塊覆蓋,種子較輕、有種翅,成熟后隨風散布,很難進入土壤層,種子難以在土壤上著床,不能萌芽成苗,也是造成多脈鐵木幼苗較少的關鍵外在因素。另外隨著生態公益林保護力度的增大,多脈鐵木群落各種樹木的生長,森林的郁閉度達90%以上,喜光的多脈鐵木幼苗得不到足夠的光照條件,因而在Ⅰ、Ⅱ齡級遭遇1次死亡高峰。從第Ⅲ齡級開始,多脈鐵木種群保持較高的穩定性。
種群生命表可以判斷植物種群發展趨勢,存活曲線能夠直觀反映種群個體在各齡級的存活狀況[12],Deevey-Ⅲ型存活曲線主要表現為早期死亡率高,一旦存活到某一年齡,死亡率就較低,可視為增長種群[13]。本研究結果表明,多脈鐵木種群的存活曲線屬于Ⅲ型,Ⅰ、Ⅱ齡級的幼齡個體受到嚴酷的環境篩作用,對幼齡樹的生存不利;但到Ⅲ齡級后,種群呈增長型結構,這與種群的動態量值和動態指數的結果也是一致的。從種群的時間序列分析,5 a后,該物種在Ⅴ齡級前均沒有新的個體增加,低齡級的個體數量嚴重不足,種群可能面臨衰退。
針對多脈鐵木種群的現狀和發展趨勢,可從以下兩方面考慮該種群的保護和恢復策略。一方面從種群個體自身因素考慮,在加強對原生境及植株的保護與監測的基礎上,加大科研力度,開展多脈鐵木生殖生態學研究,如種子萌發率、成苗率和高效繁育技術等,提高實生苗的數量,以擴大其種群分布范圍;另一方面是采取“近地保護”措施,將人工繁育的實生苗在多脈鐵木現有分布區附近進行適當種植,建立人工保護點。從種群所處生境考慮,適度開展人為干預,搬動部分大石塊,展露部分區域土壤,以保證多脈鐵木種子順利入土,促進種子萌發。在此基礎上,間伐過密的非目標樹種,適當擴大林窗,以降低森林郁閉度,增加林內光照,以利于幼苗的生長,從而突破多脈鐵木由Ⅰ齡級向Ⅱ齡級轉化的“瓶頸”。
