激活型人源化抗人CD137單克隆抗體的研制與生物學活性鑒定
2018-09-05 03:28:48王華菁徐依云楊煥鳳
安徽醫科大學學報 2018年9期
關鍵詞:檢測
何 妍,王華菁,陸 婷,徐依云,黃 勇,楊煥鳳
CD137(CD137/TNFRSF9)可能是第一個被確定為免疫治療靶標的TNFRSF成員[1-2]。 CD137由調節性T細胞(Treg)和幾種先天免疫細胞群體組成型表達[3], CD137抗體通過三聚體連接人腫瘤壞死因子受體相關因子1(TRAF1)和人腫瘤壞死因子受體相關因子2(TRAF2),調節免疫細胞功能通路NF-κB的活化[4],能誘導干擾素-γ(IFN-γ)和白介素2的增殖和產生,并通過上調抗細胞凋亡途徑(包括Bcl-xL)來增強細胞存活,CD137通過刺激抗體依賴性細胞介導的細胞毒性(ADCC)增強NK細胞的增殖。CD137對腫瘤反應性T細胞存在優先表達及增加T細胞募集到腫瘤部位的能力,可以保護T細胞免受活化誘導的細胞死亡反應(activiation-induced cell death,AICD),同時增強T細胞的毒活性[5],然而,目前已研發的CD137抗體對于CD137蛋白的特異性識別結合能力有限,對NF-κB信號通路的激活能力有限,促進PBMC細胞增殖有限,或誘導免疫細胞分泌細胞因子的能力有限,因此亟待開發新的人源化CD137抗體以用于新藥研發。
1 材料與方法
1.1試劑與材料
1.1.1試劑 PCR buffer、dNTP、rTaqDNA聚合酶、DL 2000和DL 15000 DNA Marker、限制性核酸內切酶均購自日本TaKaRa公司;DNA片段回收試劑盒和質粒小提試劑盒,蛋白Marker均購自北京天根公司;Gibson Asse、bly Master Mix購自美國NEB公司;完全弗氏佐劑、不完全弗氏佐劑、Accuspin管均購自美國Sigma公司;脫脂奶粉、PE-Mouse anti-hCD137抗體購自美國BD公司;Lipofectamine 2000轉染試劑、RPMI 1640和DMEM 培養基、GIBCO胎牛血清(FBS)均購自美國Thermo Fisher Scientific公司;HRP標記山羊抗小鼠IgG、goat anti-mouse-647均購自美國Abcam 公司; Protein A 純化柱購自美國GE公司。
1.1.2菌種與質粒 真核表達載體PCDNA3.4由該治療性疫苗研究中心實驗室保存;大腸桿菌DH5α感受態及大腸桿菌BL21購自北京天根公司。
1.1.3細胞與病毒 小鼠骨髓瘤細胞SP2/0購自中國科學院典型培養物保藏委員會細胞庫,人胚胎腎細胞293F為治療性疫苗研究中心實驗室保存。PBMC細胞來自于志愿者,標本獲取與使用方法得到上海長征醫院醫學倫理委員會的批準。
1.1.4實驗動物與組織來源 SPF級6~8周齡Balb/c、F1小鼠,購自上海西普爾-必凱實驗動物有限公司,該實驗對動物的處置均符合動物倫理學標準。由復旦大學實驗動物中心協助飼養。
1.2方法
1.2.1蛋白真核表達質粒的構建與鑒定 通過DNAstar軟件比較人CD137胞外段的親水性與免疫原性,選取CD137第86~186個氨基酸所在的胞外段作為免疫靶向片段,并設計成在重組肽段羧基端(c端)融合有一個His標簽的形式。隨后,將以上所對應的基因片段5′端添加HindIII酶切位點、3′端添加Xbal酶切位點后交由上海桑尼生物公司合成,通過酶切位點HindIII和Xbal將該基因片段插入pCDNA3.4載體中,構建CD137真核表達質粒pCDN3.4,轉化大腸桿菌DH5α感受態,通過HindIII/Xbal雙酶切和PCR鑒定(上游引物p1:5′-CCCAAGCTTATGGGAAACAGCTGTTAC-3′;下游引物p2:5′-GCTCTAGATCAATGGTGATGGTGATGATGCTGCGGAGAGTGT-3′)篩選陽性克隆后由上海桑尼生物公司測序。
1.2.2重組蛋白的表達與純化 運用脂質體轉染法將構建成功的pCDNA3.4/hCD137-his 重組載體轉染 293F細胞。 取對數生長期 293F 細胞,于37 ℃、120 r/min的CO2細胞搖床中培養,培養密度為1.0×105/ml,細胞生長密度達到倍數增殖時,進行轉染;將脂質體-載體混合液加入細胞,擴大培養7 d后收集上清液進行純化,其對照為轉染pDNA3.4-GFP空載體的293F細胞(293/mock)。
1.2.3鼠抗人CD137單克隆抗體制備 將純化的重組蛋白人CD137與等體積的弗氏佐劑混合乳化均勻(初免使用完全弗氏佐劑,加強免疫使用不完全弗氏佐劑),隨后對3只Balb/c雌鼠經皮下多點注射免疫,免疫劑量為100 μg/只,體積為300 μl/只,共免疫3次,時間間隔為2周。初免后每2周從小鼠尾靜脈采血,在第3次免疫完成后進行一次腹腔沖擊免疫,免疫劑量為50 μg/只,3 d后取該小鼠脾細胞與SP2/0細胞做融合實驗;HAT法篩選10 d后換HT培養基。在第14天時吸取雜14交瘤細胞培養上清液進行人CD137包被的ELISA檢測,對檢測結果為陽性的亞克隆采用有限稀釋法進行至少2次以上的克隆化,直到篩選出穩定分泌單克隆抗體的融合細胞株。
1.2.4表位鑒定 為了確定CD137蛋白抗體的表位結合區,在公開的犬CD137蛋白序列(Ref. Seq. XM 845243)上對人CD137蛋白(4-1BB蛋白)的胞外結構域進行一系列突變。具體的突變位點參見表1,將突變后的序列構建到pHAGE-CMV-MCS-IRES-ZsGreen病毒包裝載體上,并感染293FT細胞通過流式分選培養制成穩定表達細胞株,將細胞株結合純化的抗體,無突變的CD137轉基因細胞株做為陽性對照,通過流式檢測法中陽性比例確定表位。

表1 CD137人至犬序列突變體表
1.2.5人源化抗體的制備 使用TRIzoL(購自上海生工生物)提取雜交瘤細胞株的總RNA,用逆轉錄試劑盒將mRNA逆轉錄成cDNA,以cDNA為模板做PCR,使用Ig-Primer Sets試劑盒分別克隆抗體重鏈和輕鏈可變區序列;將克隆得到的抗體重鏈和輕鏈可變區序列測序。將PCR產物克隆入pCDNA3.4載體進一步擴增表達。構建嵌合抗體,將上述測序正確的質粒構建成真核表達載體 pcDNA3.4(+)(VHCH)和 (VLCL)。將重、輕鏈表達載體后共同瞬時轉染293F細胞表達抗體蛋白。在37 ℃、5%CO2的搖床中培養7 d后收集上清液,用瓊脂糖凝膠protein A層析填料純化上清液中的嵌合抗體。
人源化過程根據以下原則進行:在比對鼠源抗體與胚系模板的基礎上,完全保留CDR區的氨基酸序列,參考已有文獻報道的關鍵性骨架區殘基,以及已上市/臨床的基于IGKV4-1*01的人源化抗體確定人源化序列,對骨架區的關鍵鼠氨基酸進行選擇性保留,產生初步人源化序列,之后再根據突變對親和力的影響逐漸減少骨架區鼠氨基酸數。共有四個突變體,c6F5為人鼠嵌合抗體,c6F5-IgG2為鼠可變區構建在人的IgG2骨架上,c6F5-HR1為可變區保留一個鼠源氨基酸,發現表達量下降后將影響的氨基酸序列回復突變為hu6F5 A/V back人源化抗體。
全基因合成人源化抗體hu6F5重鏈可變區和輕鏈可變區,將抗體重鏈可變區和輕鏈可變區分別插入pcDNA3.4(+)-sshkappa和pcDNA3.4(+)-sshIgG2表達載體,將重、輕鏈表達載體后共同瞬時轉染293F細胞表達抗體蛋白。在37 ℃、5%CO2的搖床中培養7 d后收集上清液,用瓊脂糖凝膠protein A層析填料純化上清液中的人源化抗體hu6F5。
1.2.6抗體體外功能鑒定
1.2.6.1NF-κB信號通路檢測 制備表達人CD137以及穩定整合的NF-κB螢光素酶報道基因的293T細胞。收獲細胞,洗滌及重懸浮于DMEM完全培養基,細胞密度為6×105個/ml。將50 μl細胞鋪板于白色96孔平板的每個測定孔中。在存在2.5:1比率的交聯抗體Fab'山羊抗人IgG Fc的條件下,分別加入human IgG2(同型對照)以及各抗體。加入抗體后將平板在37 ℃溫育5 h。加入75 μl的Bright-Glo Luciferase試劑,使用酶標儀測量螢光素酶活性的量。
1.2.6.2人源化CD137抗體誘導細胞因子表達 用不同濃度人源化CD137抗體、抗CD3-e克隆UCHTl以及UTOMILUMAB(作為陽性對照)、cG33-IgG2(作為陰性對照)刺激外周血單個核細胞(PBMC)。離心后檢測培養上清液。通過ELISA法檢測上清液中的細胞因子。受試樣品和標準品加入到抗細胞因子包被的96孔板中。在室溫放置2 h后,在PBS-T中洗板3次,先與工作檢測抗體保溫,然后加入底物。450 nm處檢測吸光度,基于標準曲線計算濃度。
1.2.7抗體親和力鑒定
1.2.7.1BLI檢測抗體親和力 結合人 CD137的某些抗體的結合動力學使用 Fortebio octet 384儀器通過生物膜干涉(BLI)技術測量。包含氨基酸24-186的重組人CD137/his標簽蛋白由本研究室制備。將蛋白質溶解于含PBS、0.1%牛血清白蛋白(BSA)及0.02%Tween 20的緩沖液中。抗體通過與AMQ傳感器固定,在PBS、0.1%BSA及0.02% Tween 20中將抗體濃度稀釋為50 nmol/L,將重組蛋白人CD137在1.56~100 nmol/L濃度范圍以1 500 r/min結合10 min,隨后以相同轉速在PBS、0.1% BSA及0.02% Tween 20中進行10 min解離。結合的復合物通過甘氨酸脈沖再生。使用octet Data Analysis 9.0軟件進行數據分析。傳感圖擬合為簡便的1 ∶1 Langmuir 結合模型。抗體示出與重組人CD137的可逆結合。
1.2.7.2FACS檢測抗體親和力 3倍稀釋不同人源化抗體,加入到CD137-293細胞中,再用Alexa Fluor?647-GOAT Anti-Human Ab染色后上流式細胞儀檢測熒光。當抗原為一定數量,逐漸增加抗體濃度,抗體抗原結合達到飽和,然后求出抗原飽和50%時結合的抗體濃度。其倒數即為親和常數,畫出飽和曲線,EC50即為親和常數。
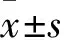
2 結果
2.1真核表達質粒的構建與鑒定將pCDNA3.4真核表達載體和合成的人CD137胞外段基因片段都進行HindIII/Xbal雙酶切處理,分別回收后在T4連接酶作用下進行連接,并轉化至大腸桿菌DH5α感受態中構建重組質粒。提取質粒,挑取經HindIII/xbal雙酶切和PCR方式鑒定均可獲得約558 bp片段的陽性克隆(圖1A),由上海桑尼生物公司測序。經鑒定,真核表達重組質粒pCDNA3.4-CD137構建成功。
2.2重組蛋白的表達純化與鑒定以293F細胞和293F/mock為對照,用 ELISA 方法檢測其上清液中hCD137蛋白的表達。結果顯示,293F、293F/mock上清液中hCD137的表達均為陰性,293F/hCD137-His 基因轉染細胞上清液hCD137表達水平較高,表達的目的蛋白能夠以分泌型形式存在。SDS-PAGE結果見圖1B。

圖1 CD137 表達載體、重組蛋白及CD137單抗6F5的鑒定A: h-CD137基因的克隆和pCDNA3.4重組載體的酶切驗證;B: h-CD137-his融合蛋白的蛋白純化及Western blot驗證;C: h-CD137單抗6F5的蛋白純化
2.3鼠抗人CD137抗體的表位測定將突變后的序列構建到pHAGE-CMV-MCS-IRES-ZsGreen病毒包裝載體上,并感染293FT細胞通過流式分選培養制成穩定表達細胞株,將細胞株結合6F5純化抗體,無突變的CD137轉基因細胞株做為陽性對照,通過流式檢測法中陽性比例確定表位,結果見表2。表位在a.a.30~100之間。

表2 抗體6F5表位圖
2.4人源化抗人CD137抗體的信號通路檢測使用Bright-Glo Luciferase 檢測試劑盒檢測加入抗體后反應5 h的CD137轉基因細胞株內熒光素酶的表達情況,讀值見圖2。由圖可見6F5能夠激活NFκB信號通路,并產生熒光素酶來使底物產生熒光值,該值與陰性值相比差異有顯著差異(P<0.01)。

圖2 NF-κB信號通路的檢測1:m6F5;2:c6F5;3:c6F5-IgG2;4:c6F5-HR1;5:hu6F5-A/V back;6:UTOMILUMAB;7:Cg33-IgG2 ;與Cg33-IgG2組相比:**P<0.01
2.5抗體對PBMC細胞激活功能的鑒定用不同濃度人源化后不同突變hCD137抗體和抗 CD3-e克隆UCHTl刺激 PBMC, 使用試劑盒檢測培養上清液中的IL-2(圖3A)、IFN-γ含量(圖3B)。結果顯示 hCD137 抗體能顯著提高細胞因子的分泌,與正常PBMC培養上清液中細胞因子含量相比差異有顯著性。
2.6抗體親和力分析分別使用BLI和FACS的方法檢測人源化后不同突變抗體的親和力。生物膜層光學干涉技術 (BiolayerInterferometry,BLI) 是通過固化CD137單抗6F5,結合CD137蛋白(結合濃度依次為100、50、25、12.5、6.25、3.12、1.56 nmol/L),檢測6F5抗體及其他不同人源化抗體與CD137蛋白的親和力(圖4A)。結果顯示人源化抗體親和力未因氨基酸突變而下降(表3)。

圖3 人源化抗人CD137對PBMC的影響A:PBMC與SEA及CD137抗體共培養4 d后的IL-2含量;B:PBMC與CD3抗體及CD137抗體共培養4 d后的IFN-γ含量;與Cg33-IgG2 組相比:***P<0.001; 與PBMC組相比:##P<0.01;1:m6F5;2:c6F5;3:c6F5-IgG2;4:c6F5-HR1;5:hu6F5-A/V back;6:UTOMILUMAB;7:Cg33-IgG2 ;8. PBMC
不同抗原抗體反應呈現的平均熒光強度直接定量評價抗體親和力大小。hCD137-293T應用于分析抗體親和力,抗體的EC50進行分析。各抗體的LogEC50顯示人源化抗體的親和力未下降,同BLI數據相同(圖4B)。
3 討論
腫瘤微環境富含CD137,因為其在該位置由效應子和調節性T淋巴細胞表達[6]。更重要的是,必須將CD137激動劑靶向或局部遞送至腫瘤,以暴露最大化并限制系統毒性(例如在肝和骨髓中)。事實上,大多數表達的CD137僅存在于腫瘤微環境中[7]。但是這些信號機制由TNFR家族和其他表面受體系統的其他成員共享[8],因此可能會有脫靶副作用。總的來說,基于CD137的免疫治療可能為臨床和轉化發展提供了許多機會,因此得到有功能性的CD137人源化抗體成為迫切需要。目前得到的人源化抗體已經證明了體外功能試驗及親和力。該人源化抗體的表位和現有CD137抗體表位不同,可能會降低藥物毒性。

表3 各抗體KD值

圖4 抗體親和力定量分析結果A:BLI測定不同CD137單抗的親和力;B:FACS測定不同CD137抗體親和力
下一步研究方向是證實該人源化抗體在體內實驗上抗腫瘤效果,建立抗體藥物體內外藥效學評價標準,并且可能開展聯合用藥評價體內外藥效學。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
