碭山酥梨與豐水梨正、反交群體果實(shí)性狀的遺傳特征
2018-09-07 08:54:26秦夢帆
江蘇農(nóng)業(yè)科學(xué) 2018年16期
白 冰, 劉 倫, 秦夢帆, 谷 超, 吳 俊
(南京農(nóng)業(yè)大學(xué)梨工程技術(shù)研究中心,江蘇南京 210095)
梨屬于薔薇科、梨亞科(Powaceae)、梨屬(PyrusL.)植物,是重要的溫帶果樹之一,距今已有近3 000年的栽培歷史[1]。2013年世界上梨的產(chǎn)量和栽培面積在薔薇科果樹中僅次于蘋果,位居第2(數(shù)據(jù)來源于聯(lián)合國糧食及農(nóng)業(yè)組織,http://faostat.fao.org)。梨的許多性狀,如果實(shí)大小、果形、可溶性固形物含量、石細(xì)胞含量及果實(shí)的綜合品質(zhì)等是消費(fèi)者關(guān)注的重要經(jīng)濟(jì)性狀,也是育種工作者研究的重點(diǎn)。
目前,已有一些關(guān)于梨果實(shí)性狀遺傳規(guī)律的報道。果實(shí)大小為多基因控制的數(shù)量遺傳性狀,雜種后代果實(shí)趨于變小,但出現(xiàn)超高親植株[2-3];吳忠華等以11個梨雜交組合為材料進(jìn)行研究發(fā)現(xiàn),雜種后代平均果實(shí)質(zhì)量均大于親中值,果實(shí)質(zhì)量遺傳表現(xiàn)出超中偏大現(xiàn)象[4];何天明等研究發(fā)現(xiàn),果實(shí)中的可溶性固形物含量表現(xiàn)為超親遺傳,遺傳效應(yīng)中的加性效應(yīng)所占比例較大[5-7];關(guān)于石細(xì)胞遺傳規(guī)律的研究表明,后代果實(shí)石細(xì)胞含量呈增多趨勢[8-9]。由于試材各異,且研究方法不同,研究所得結(jié)論不完全一致;同時由于構(gòu)建雜交群體的難度較大,關(guān)于相同親本正、反交后代果實(shí)性狀的遺傳特性和差異的研究較少,因此,梨果實(shí)相關(guān)性狀的遺傳規(guī)律還不明確,無法為雜交育種中親本的合理選擇以及雜種后代果實(shí)性狀的預(yù)測提供科學(xué)依據(jù),因此有必要開展深入系統(tǒng)的研究。
本試驗(yàn)以碭山酥梨和豐水梨及其正、反交后代個體為材料,連續(xù)2年對其雜交后代果實(shí)的單果質(zhì)量、縱徑、橫徑、果形指數(shù)、可溶性固形物含量、石細(xì)胞含量的遺傳規(guī)律進(jìn)行分析,以期豐富梨的雜交育種遺傳理論,指導(dǎo)雜交親本選配,實(shí)現(xiàn)目標(biāo)性狀的遺傳改良。
1 材料與方法
1.1 研究材料
親本及正、反交群體后代果實(shí)均采自南京農(nóng)業(yè)大學(xué)江浦園藝實(shí)驗(yàn)站梨雜交育種圃。碭山酥梨×豐水梨(正交)、豐水梨×碭山酥梨(反交)雜交群體實(shí)生樹分別于2006、2005年定植。雜交后代實(shí)生樹株距、行距分別為0.6、3.0m。親本株距、行距分別為4.0、6.0m。所有的植株均在盛花后30d左右進(jìn)行疏花,并進(jìn)行相同的管理。
1.2 樣品采集與處理
于后代及親本的果實(shí)成熟期進(jìn)行采摘,果實(shí)成熟期根據(jù)果實(shí)硬度、種子變褐程度和往年對果實(shí)成熟期的記錄進(jìn)行確定。所有樣品均由同一個人采集,以控制成熟度的一致性,減小誤差。由于果樹大小年及部分植株結(jié)果不理想,2013年,在碭山酥梨×豐水梨、豐水梨×碭山酥梨2個群體中分別采集52、72株后代;2014年,2個群體中分別采集44、79株后代,取2年測定值的平均值進(jìn)行統(tǒng)計分析。采用四分法在每株樹冠外圍不同方向隨機(jī)選取10個大小一致、無病蟲害的果實(shí)用于各項(xiàng)指標(biāo)的測定。
1.3 指標(biāo)測定
果實(shí)縱徑、橫徑用游標(biāo)卡尺測量,單位為mm;果形指數(shù)的計算方法為果形指數(shù)=果實(shí)縱徑/果實(shí)橫徑;單果質(zhì)量用百分之一天平測定,單位為g;果實(shí)可溶性固形物含量采用手持折射儀(RHBO-90)測定[10];采用鹽酸處理[11]和冷凍分離[12]相結(jié)合的方法進(jìn)行石細(xì)胞分離,石細(xì)胞含量的具體測定方法參考王丹陽等的方法[13]。
遺傳傳遞力=F/MP×100%;
變異系數(shù)=標(biāo)準(zhǔn)差/F×100%。
式中:F為后代各指標(biāo)的平均值;MP為親中值。
1.4 數(shù)據(jù)分析
應(yīng)用Excel2013和SPSS20.0軟件對數(shù)據(jù)進(jìn)行統(tǒng)計分析。
2 結(jié)果與分析
2.1 碭山酥梨與豐水梨正、反交后代單果質(zhì)量的遺傳變異
研究結(jié)果(圖1)表明,碭山酥梨與豐水梨正、反交后代果實(shí)單果質(zhì)量呈現(xiàn)連續(xù)性變異,為多基因控制的數(shù)量遺傳性狀。由表1可知,正、反交后代的單果質(zhì)量平均值都小于親中值,且反交后代小于低親。在所研究的后代個體中,正交后代低于低親的株數(shù)達(dá)59.76%,反交后代達(dá)77.73%,表明對豐水梨來說,作母本更容易將其小果型傳遞給后代;超高親率前者為13.29%,后者為2.58%,說明碭山酥梨作母本時更易于將大果型傳遞給后代。正、反交群體的變異系數(shù)分別為34.53%、35.08%,極值相差較大,說明后代廣泛分離,這是由有性雜交過程中雙親的非加性效應(yīng)解體所致,進(jìn)而導(dǎo)致雜交后代呈現(xiàn)回歸變異。


表1 碭山酥梨和豐水梨正、反交后代單果質(zhì)量的遺傳變異
2.2 碭山酥梨和豐水梨正、反交后代果實(shí)縱徑、橫徑的遺傳變異
研究結(jié)果(圖2、圖3)表明,碭山酥梨和豐水梨正、反交后代果實(shí)的縱徑和橫徑均呈現(xiàn)連續(xù)性變異,為數(shù)量性狀遺傳,呈退化負(fù)向趨勢。由表2、表3可知,正、反交后代果實(shí)的縱徑、橫徑平均值都小于親中值。正、反交后代中縱徑低于低親的個體分別有58.91%、70.07%,正交后代橫徑低于低親的個體比反交后代少,說明以豐水梨為母本時,縱徑和橫徑較小的性狀更易傳遞給后代。正交群體中出現(xiàn)的超高親個體多于反交群體,表明以碭山酥梨作母本時,后代更易出現(xiàn)大果型。由此可見,果實(shí)縱、橫徑的遺傳特征與單果質(zhì)量十分類似,而單果質(zhì)量與縱徑、橫徑共同決定了梨果實(shí)的大小。

2.3 碭山酥梨和豐水梨正、反交群體后代果形指數(shù)遺傳變異
從圖4、表4可以看出,碭山酥梨和豐水梨正、反交后代果形指數(shù)連續(xù)變異,呈正態(tài)分布,其中正交后代果形指數(shù)的平均值小于親中值,反交后代的果形指數(shù)大于親中值,甚至大于高親值,且遺傳傳遞力比較強(qiáng)。正、反交后代的超高親率分別為 38.60%、54.82%,低于低親率分別為48.45%、28.93%,表明在果形指數(shù)的遺傳上受父本的影響較大。在育種中,根據(jù)不同的育種目標(biāo),要特別注意父本的合理選擇。后代果形指數(shù)向兩極變異,介于兩親之間的個體較少。
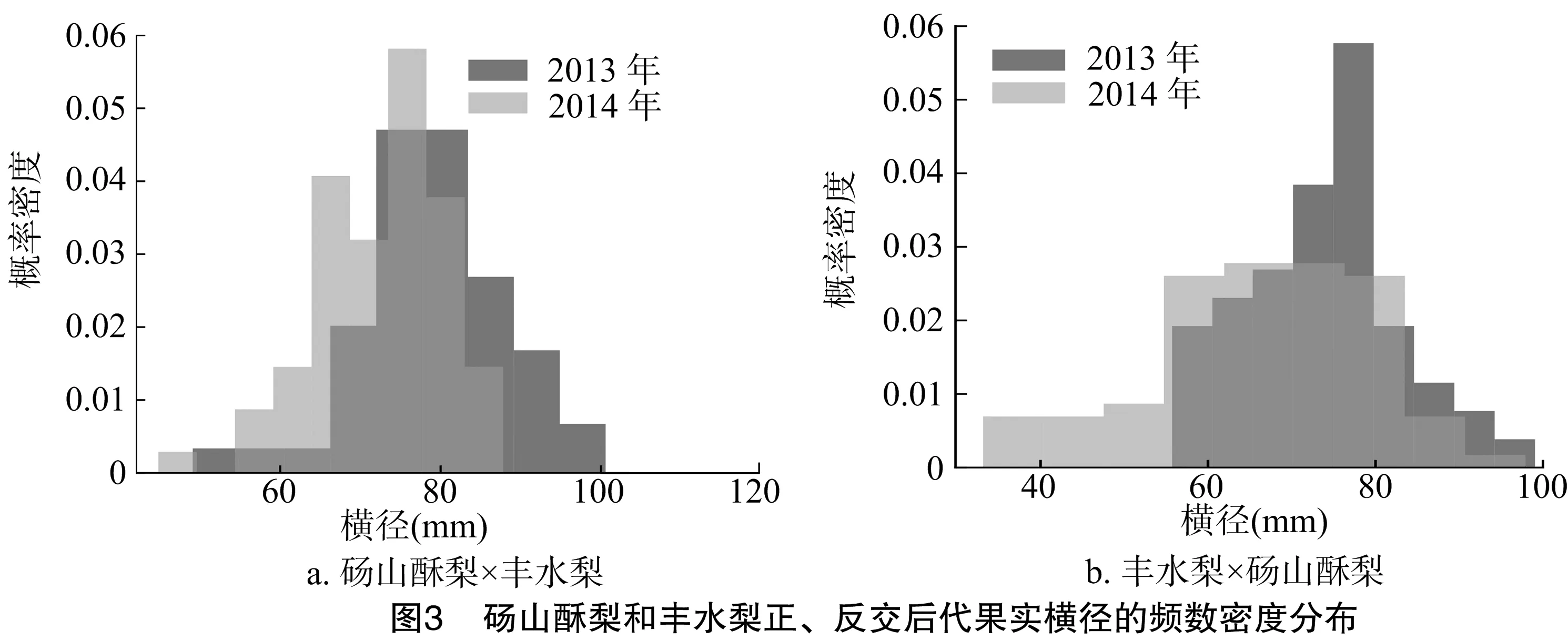
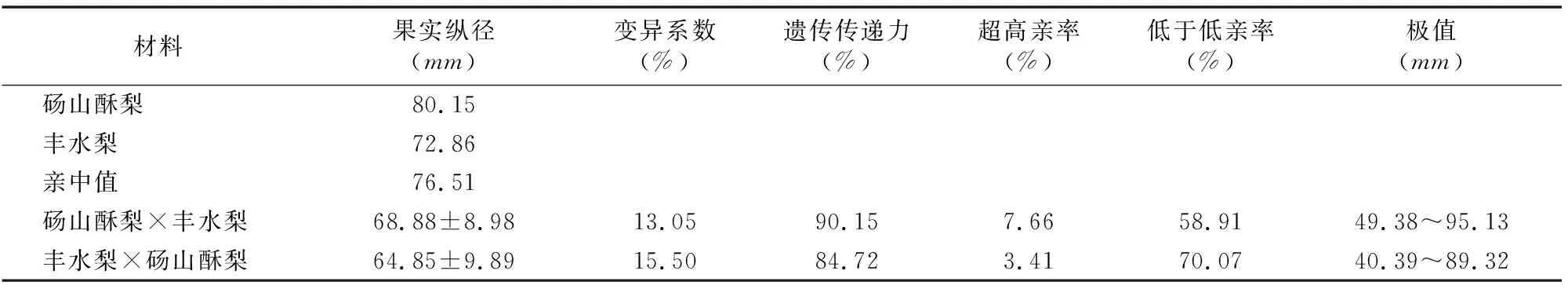
表2 碭山酥梨和豐水梨正、反交后代果實(shí)縱徑的遺傳變異

表3 碭山酥梨和豐水梨正、反交后代果實(shí)橫徑的遺傳變異


表4 碭山酥梨和豐水梨正、反交后代果形指數(shù)的遺傳變異
2.4 碭山酥梨和豐水梨正、反交群體后代果實(shí)可溶性固形物含量的遺傳變異
從圖5、表5可以看出,碭山酥梨和豐水梨正、反交后代果實(shí)的可溶性固形物含量呈現(xiàn)數(shù)量性狀遺傳并表現(xiàn)為超親遺傳,其平均值均大于親中值,且均大于高親值,其遺傳傳遞力分別為125.00%、120.18%,說明果實(shí)可溶性固形物含量的遺傳主要由基因間的加性效應(yīng)控制,非加性效應(yīng)的影響較小,有增高趨勢。正、反交后代的超高親率分別為87.30%、88.13%,具有明顯的雜種優(yōu)勢,正、反交后代中分別出現(xiàn)含量為17.80%、18.44% 的高親株,組合中親本的位置關(guān)系對后代果實(shí)可溶性固形物含量并無較大影響。正交后代的變異系數(shù)為 12.95%,極值差為7.8%;反交后代的變異系數(shù)是13.49%,極值差是8.99%,后代分離程度相當(dāng)。
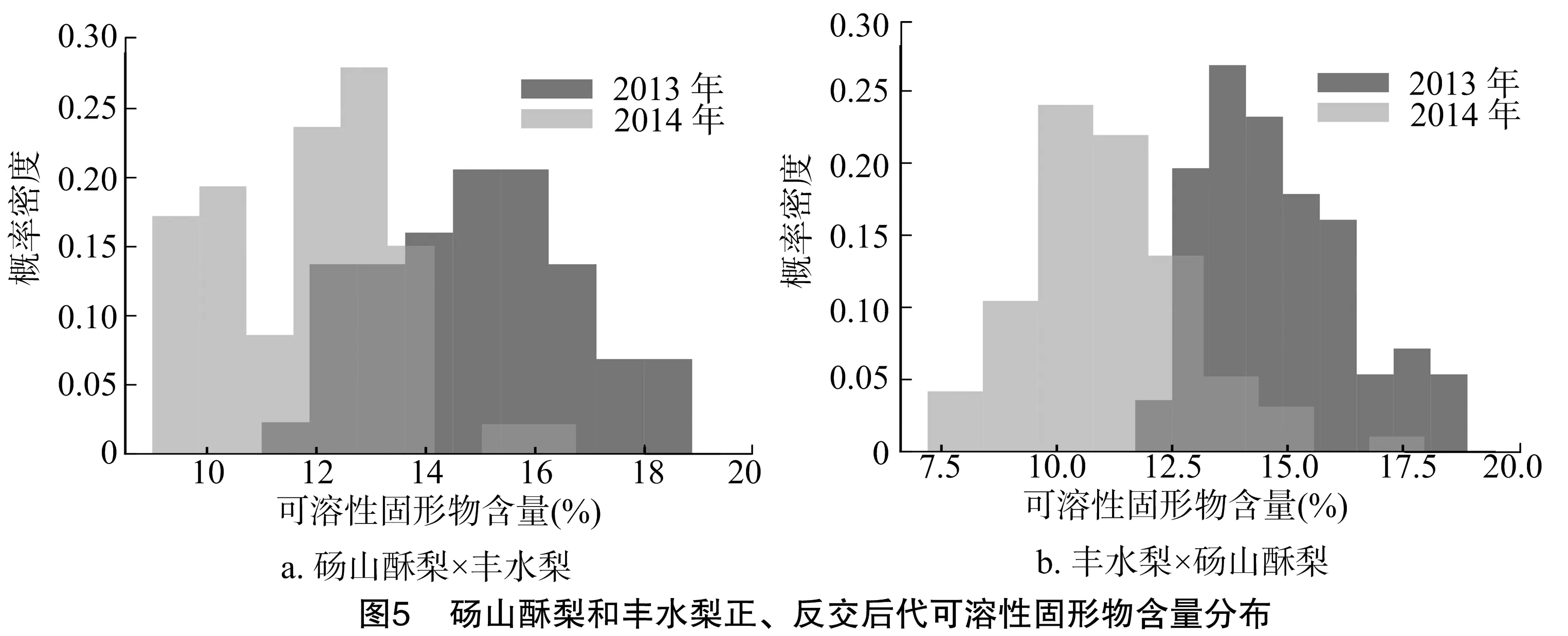

表5 碭山酥梨和豐水梨正、反交后代果實(shí)可溶性固形物含量的遺傳變異
2.5 碭山酥梨和豐水梨正、反交群體后代果實(shí)石細(xì)胞含量遺傳變異
由圖6可知,碭山酥梨和豐水梨正、反交后代果實(shí)的石細(xì)胞含量呈連續(xù)變異。由表6可知,正、反交后代的果實(shí)石細(xì)胞含量介于雙親之間,呈明顯的趨中分布。其中,反交后代石細(xì)胞含量高于正交后代。正交群體在連續(xù)2年調(diào)查中沒有出現(xiàn)超高親的后代,其后代石細(xì)胞含量有所下降,而反交群體則出現(xiàn)了 17.49% 的超高親個體。豐水梨中石細(xì)胞含量較低,以其作父本的后代果實(shí)石細(xì)胞含量比作母本時低,說明石細(xì)胞含量的遺傳比較復(fù)雜,在育種過程中,若選擇低石細(xì)胞含量的株系作為母本,則須要注意選擇石細(xì)胞含量低的父本。群體變異系數(shù)高,后代廣泛分離,說明本雜交組合具有選育低石細(xì)胞含量后代的潛力。

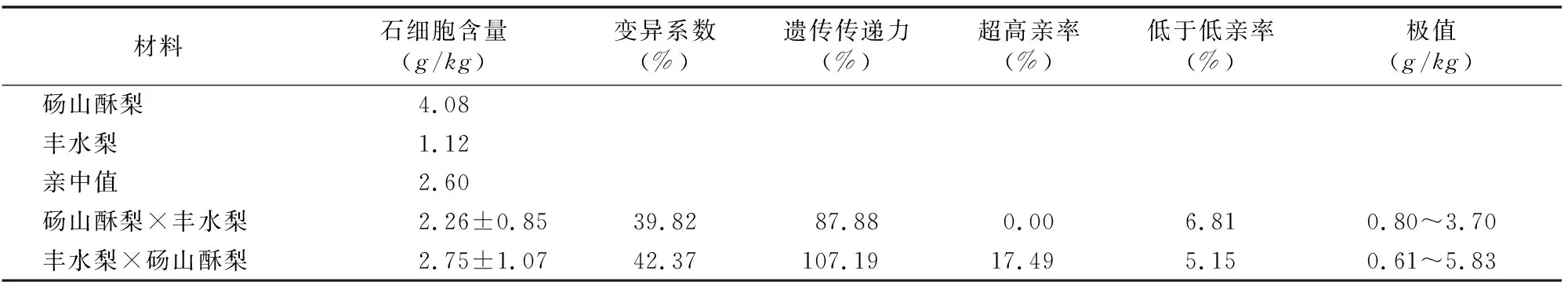
表6 碭山酥梨和豐水梨正、反交后代果實(shí)石細(xì)胞含量的遺傳變異
3 討論與結(jié)論
梨在種間或品種間進(jìn)行有性雜交時,由于親本的非加性效應(yīng)解體,后代出現(xiàn)分離,劣變率往往很高,但也會出現(xiàn)超高親的植株,仍可為選擇理想的品種提供有利條件,這已成為育種工作者關(guān)注的重點(diǎn)。本研究中,碭山酥梨和豐水梨正、反交后代果實(shí)的單果質(zhì)量呈現(xiàn)廣泛的連續(xù)性偏小分布,平均單果質(zhì)量均小于親中值,這與前人的研究結(jié)果[9,14-15]相一致。正、反交后代均出現(xiàn)超高親植株,說明即使后代廣泛分離,但由于雜種優(yōu)勢的作用,仍使獲得大果型品種成為可能。雜種后代的縱徑、橫徑與雙親相比也出現(xiàn)趨小變異現(xiàn)象,與單果質(zhì)量的遺傳規(guī)律相似。
碭山酥梨和豐水梨正、反交后代果形指數(shù)的遺傳傳遞力較強(qiáng),果形指數(shù)向兩極分化,介于雙親之間的個體少,正、反交后代的超高親植株分別為38.60%、54.82%,這與崔艷波等的結(jié)論[9]不同。本研究中正、反交后代果形指數(shù)變異系數(shù)較低、極值差較小、分離程度較低,這與郭黃萍等得出果形指數(shù)變異系數(shù)較高、分離廣泛的結(jié)論[3]不同,原因可能與碭山酥梨和豐水梨的果形相似有關(guān)。
碭山酥梨和豐水梨雜交后代可溶性固形物含量表現(xiàn)出超親遺傳,后代果實(shí)的平均含量連續(xù)2年均大于親中值,遺傳傳遞力都超過了100%,具有很強(qiáng)的雜種優(yōu)勢。其他雜交組合也有相關(guān)報道,王家珍等以蘋果梨為中心的7個雜交組合為試材,研究表明,雜交后代可溶性固形物平均含量超過親中值[6];崔艷波等認(rèn)為,鴨梨和京白梨的正、反交后代可溶性固形物含量均呈升高趨勢,雜種優(yōu)勢明顯[9];賈立邦等發(fā)現(xiàn),其研究中的所有雜交組合后代可溶性固形物含量均高于雙親,且只有2個組合低于高親[2]。上述研究結(jié)果說明,梨果實(shí)中可溶性固形物含量是受多基因控制的數(shù)量遺傳性狀,遺傳效應(yīng)中加性效應(yīng)所占比例較大。而李先明等的研究結(jié)果[7]與以上研究稍有不同,可能是由于試材和研究方法不同所致,同時自然氣候條件及栽培技術(shù)措施的影響也不容忽視。
石細(xì)胞的含量直接影響梨品質(zhì)的優(yōu)劣,育種工作者傾向于選擇石細(xì)胞少的品種[16]。李俊才等報道認(rèn)為,梨果實(shí)石細(xì)胞含量為多基因控制的數(shù)量遺傳性狀,后代果實(shí)石細(xì)胞含量有明顯增多的趨勢[8];楊宗駿通過調(diào)查36個梨雜交組合得出后代石細(xì)胞含量偏多的結(jié)論[17]。本試驗(yàn)中,2個雜交群體連續(xù)2年的石細(xì)胞含量大多介于兩親本之間,豐水梨作父本時后代果實(shí)的石細(xì)胞含量較作母本時低。正、反交群體的變異系數(shù)分別為39.82%、42.37%,變異系數(shù)較大,后代石細(xì)胞含量廣泛分離。本研究的結(jié)果與之前的報道存在一些差異,可能是由于所選擇的親本石細(xì)胞含量差別較大,兩者都對后代的石細(xì)胞含量產(chǎn)生影響,因此大多表現(xiàn)為趨中遺傳。
正、反交群體是研究雜交后代是否有母本或者父本效應(yīng)的良好材料。母本效應(yīng)在甜菜[18]、枇杷[19]、大豆[20]等多種植物中都有研究報道。Liu等以碭山酥梨和豐水梨的正、反交群體為材料,研究發(fā)現(xiàn),母本對后代果實(shí)中的有機(jī)酸含量有顯著影響[21]。本試驗(yàn)中,碭山酥梨的單果質(zhì)量高于豐水梨,以碭山酥梨為母本時后代群體的單果質(zhì)量在2年中均明顯高于反交群體,碭山酥梨的大果型對后代產(chǎn)生較大影響,具有母本效應(yīng)。果實(shí)的縱徑、橫徑遺傳特征與單果質(zhì)量十分類似,同樣具有母本效應(yīng)。果形指數(shù)受父本影響較大,碭山酥梨作父本時更易于將其大果形指數(shù)傳遞給后代。豐水梨的果實(shí)石細(xì)胞含量低,以豐水梨為父本時后代果實(shí)石細(xì)胞含量較低,受父本的影響較大。

- 江蘇農(nóng)業(yè)科學(xué)的其它文章
- 基于流程的豬肉供應(yīng)鏈透明度評價指標(biāo)體系
- 農(nóng)業(yè)保險對農(nóng)業(yè)技術(shù)效率影響的差異性分析
——基于動態(tài)面板數(shù)據(jù)的GMM估計 - 基于熵權(quán)-灰色TOPSIS方法的農(nóng)產(chǎn)品綠色物流發(fā)展評價研究及應(yīng)用
- 養(yǎng)殖戶綠色運(yùn)營模式影響因素分析
——基于湖北省規(guī)模養(yǎng)豬戶調(diào)查 - 基于多因素綜合評價法的河南省農(nóng)村居民點(diǎn)整理潛力測算
- 基于共詞聚類分析的農(nóng)村土地法律問題熱點(diǎn)研究