廣東不同地區引種油棕葉片解剖結構對油棕抗寒力的影響
2018-09-11 09:45:38曾憲海焦云飛廖子榮潘登浪林位夫
廣東農業科學 2018年8期
關鍵詞:結構
曾憲海,焦云飛,廖子榮,潘登浪,林位夫
(1.中國熱帶農業科學院橡膠研究所/農業農村部儋州油棕種質資源圃,海南 儋州 571737;2.華中農業大學園藝林學學院,湖北 武漢 430070;3.海南大學熱帶農林學院,海南 海口 570228)
油棕是典型的熱帶植物,適宜生長溫度為22~33℃[1],我國熱區地處熱帶北緣,風、寒、旱等自然災害時有發生,其中低溫寒害往往給熱作生產造成重大損失[2]。油棕的抗寒能力存在個體差異且因品種而異[3],具有明顯的生態型特點,是油棕遺傳適應性對生態環境表現出的適應能力的差異所致,不但表現在生長上,也表現在其高產特性上。前期廣東引種調查發現了一些油棕植株無明顯寒害癥狀且能正常生長結果,這些優株在當地經過了長期的自然馴化,有可能存在重大的有益變異[4]。因此,評價、挖掘與利用這些優株對發展我國油棕產業具有重要意義。
目前,國內外油棕抗寒性研究主要集中在田間寒害調查、組織形態觀察、生理生化和分子生物學評價等方面[3,5],其中應用電導率及Logistic方程比較植物組織的低溫半致死溫度(LT50)能較為準確地鑒定植物抗寒性并取得比較一致的結果[6-8]。葉片是植物對低溫脅迫最敏感的器官之一,葉片的解剖結構對不同環境條件具有響應和適應機制,是評價林木抗寒性的重要指標[9]。Luis等[10]比較了溫室和試管培養條件下油棕實生幼苗的葉片解剖結構差異,發現兩種生長環境條件下葉片組織結構的基本組成相同,但溫室苗的表皮細胞和角質層比試管苗的厚且中脈較粗。曹紅星等[11]研究了油棕實生幼苗的葉片在不同低溫處理下解剖結構的差異,認為葉片總厚度和細胞組織疏松度在不同低溫處理下變化明顯,可作為油棕抗寒種質鑒定的結構指標。阮志平[12]研究認為,布迪椰子、沼地棕和油棕的抗寒性差異與葉片是否分化出柵欄組織和海綿組織有顯著相關性,同時,具有薄的角質層和上柵欄組織,且柵欄組織細胞較長、細胞排列疏松與油棕的弱抗寒性有一定的關系。劉蕊等[13]通過對5個椰子品種植株一年生幼苗的葉片解剖結構進行觀察,認為柵欄組織厚度、葉片總厚度、上(下)角質層厚度、上(下)表皮厚度和海綿組織厚度均可作為抗寒性鑒定的參考指標。本研究在前期調查基礎上,對引種廣東深圳、東莞、茂名、化州、湛江和雷州6個地區的油棕居群共38株油棕大樹進行了葉片解剖結構和低溫半致死溫度的觀測和比較分析,旨在為進一步篩選利用抗寒高產油棕種質提供參考。
1 材料與方法
1.1 試驗材料
以從海南引種廣東深圳市區(114°01′E、22°32′N,以下簡稱深圳)、東莞植物園(113°44′E、22°58′N,以下簡稱東莞)、茂名市區(110°56′E、21°39′N,以下簡稱茂名)、化州廣東農墾熱作所(110°30′E、21°38′N,以下簡稱化州)、湛江市區(110°24′E、21°12′N,以下簡稱湛江)和雷州市區(110°05′E、20°54′N,以下簡稱雷州)6個地區(居群)中的38株油棕大樹為研究對象,其中深圳12株(株號為SZ1~SZ12)、東 莞 2株( 株 號 為 DG1~DG2)、 茂 名 8株(株號為MM1~MM2)、化州8株(株號為HZ1~HZ8)、湛江 5株(株號為 ZJ1~ZJ5)、雷州3株(株號為LZ1~LZ3)。油棕的具體地理分布信息可參考Zeng等[8]的報道。葉片解剖結構特征觀察所需材料于2014年4月初采集油棕樹冠向陽面中部健康葉片(約第17片葉)葉軸中間兩邊的小葉各3~4片,于每片小葉中間處剪取寬1~2 cm的葉片材料進行FAA液固定處理24 h以上。選取其中4個葉樣做成石蠟,每個石蠟制片9~12個,選取4個較好的切片進行觀察。LT50測定所需材料于2014年4月初晴天下午采集油棕葉片樣品,按單株采集,每株3份葉樣,葉樣選取約第17片葉葉軸中部兩邊小葉,用長柄拉刀緊靠葉軸中部快速割斷小葉,選取健康、完整小葉若干,將小葉葉柄切口處用濕潤沙布包裹,然后放入塑料袋(平放20 cm×35 cm),封口并迅速帶回實驗室測定。
1.2 試驗方法
1.2.1 葉片解剖結構觀察 參照李正理的植物葉片石蠟切片法[14]并進行適當改良,具體步驟包括取材與固定、洗滌與脫水、透明、浸蠟、包埋和修蠟、切片、展片和粘片、脫蠟和染色、封片處理。采用Leica DMLB研究用生物顯微鏡對切片進行觀察并用Leica DFC500數字圖象照相機進行拍照,采用ImageJ軟件測量葉片總厚度(LT)、葉肉細胞厚度(MCT)、柵欄組織(PT)和海綿組織厚度(ST)、上角質層(DCT)和下角質層厚度(BCT)、上表皮層(DET)和下表皮層(BET)厚度、上內皮層(DHT)和下內皮層(BHT)厚度以及中脈維管束長度(MVBL)和寬度(MVBW),以上指標單位均為微米(μm),并計算細胞結構緊密度(CTR,%)、細胞結構疏松度(CSR,%)和柵欄組織厚度與海綿組織厚度比(P/S,柵海比)。葉片中脈維管束的厚度選擇中脈中較大的5個維管束進行測量并以中脈維管束的長度(與表皮垂直方向)和寬度(與表皮平行方向)來對其厚度進行描述。每份樣品觀測4個切片,每個切片每個指標取10個視野測量。
1.2.2 LT50測定 參照曾憲海等[7]的方法,油棕植株LT50(單位為℃)的測定數據均為3次重復平均值,油棕居群的數據均為所在居群全部植株測定數據的平均值。
1.2.3 方差分析、表型相關性分析和通徑分析 6個居群共38株油棕植株的15個葉片解剖結構特征參數與油棕居群和植株的LT50觀測數據進行方差分析、表型相關分析和通徑分析,其中通徑分析參照曾憲海等[7]的方法,以分析決定性狀之間的內在關系。本研究為闡明油棕葉片解剖結構特征對油棕抗寒力的影響,以LT50為自變量、葉片解剖結構特征為因變量,考察因變量和自變量之間的因果關系。一個性狀j對因變量(y)的直接影響(p)是通過一個單方向路徑揭示的,對因變量的直接影響用pij表示;而其間接影響是通過交互路徑揭示的,即一個性狀i對因變量(y)的影響是經由另一個性狀j來實現,性狀ij的相關系數用rij表示,i≠j對因變量的間接影響用rijpjy表示。
利用Excel軟件對原始數據進行統計分析,采用DPS 7.05數據處理系統進行方差分析、表型相關性分析和通徑分析[15]。
2 結果與分析
2.1 油棕葉片解剖結構的基本特征
油棕葉片上、下表皮細胞層數均為1層,表面均覆蓋著一層角質層,且上、下表皮細胞中均有氣孔結構;上、下內皮層細胞分別位于上、下表皮細胞的下方,呈無色且比表皮細胞大,細胞層數絕大部分為2層;葉肉由柵欄組織和海綿組織組成,柵欄組織無分化,僅有下柵欄組織,柵欄組織細胞層數大部分為2~3層,而海綿組織為4~6層,均為等徑形狀的薄壁細胞組成,屬同質綠色組織,細胞間隙小(圖1,封二)。葉片中脈維管束數量4~12個,為外韌型維管束(木質部在葉背面、韌皮部在葉腹面),內含原生木質部、后生木質部、韌皮部和厚壁組織,并形成一個密集的厚壁組織鞘,其中較大的中脈維管束含多個獨立韌皮部。中脈維管束排列形式多樣,其中以卵形、菱形和五邊形居多(圖2,封二)。
2.2 油棕植株和居群的葉片解剖結構及其抗寒力的差異比較分析
油棕的15個葉片解剖結構特征參數以及LT50在38株植株個體之間均存在極顯著差異(表1)。深圳居群中的SZ3植株葉片的上內皮層和下表皮層厚度最小,SZ5的上角質層厚度、柵欄組織厚度、下內皮層厚度、葉肉細胞厚度、細胞結構緊密度、柵海比和最大的細胞結構疏松度最小,SZ6的海綿組織厚度和細胞結構疏松度最小,SZ7的葉片總厚度最小,SZ8的中脈維管束長度最小。HZ5的下角質層厚度最小,HZ6的中脈維管束寬度最小,HZ2則以
上表皮層厚度最小、上皮內層厚度最大。MM2的海綿組織厚度和下角質層厚度最大,而細胞結構緊密度和柵海比則以MM3最大,此外,MM4、MM6和MM7以下表皮層厚度、上表皮層厚度和下內皮層厚度最大。ZJ4的柵欄組織厚度、葉肉細胞厚度和葉片總厚度最大,而ZJ5中脈維管束寬度最大,LZ3則以上角質層厚度和中脈維管束長度最大。植株LT50以SZ1為最大(-2.69℃)、SZ7最小(-6.19℃)。在6個油棕居群的15個葉片解剖結構特征參數中,僅葉片的中脈維管束長度(P= 0.0856)和細胞結構疏松度(P= 0.3718)在居群之間沒有顯著差異(表2)。而居群的LT50以雷州居群最大(-3.13℃)、東莞居群最小(-5.44℃)。
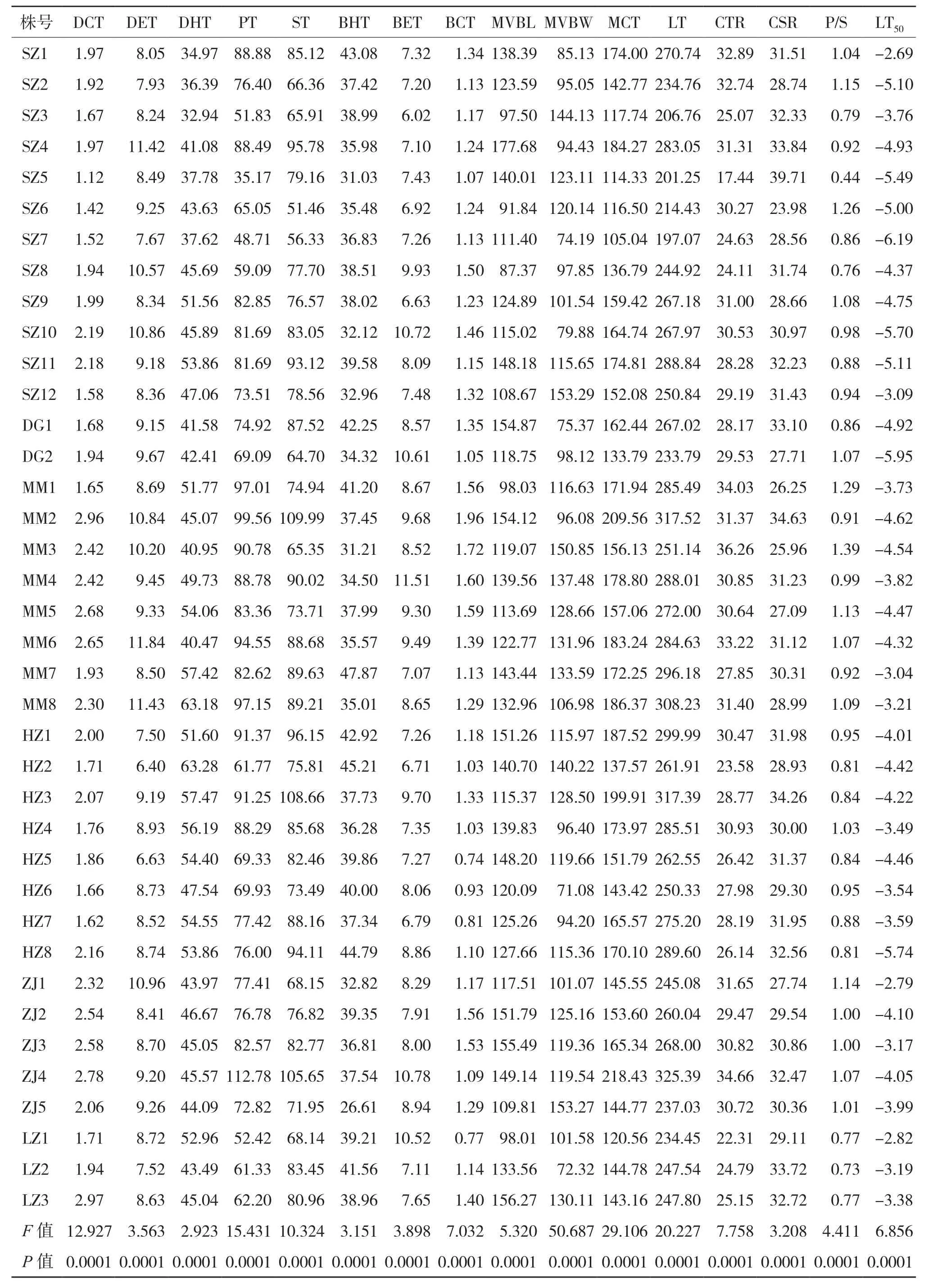
表1 油棕植株的葉片解剖結構指標測定結果
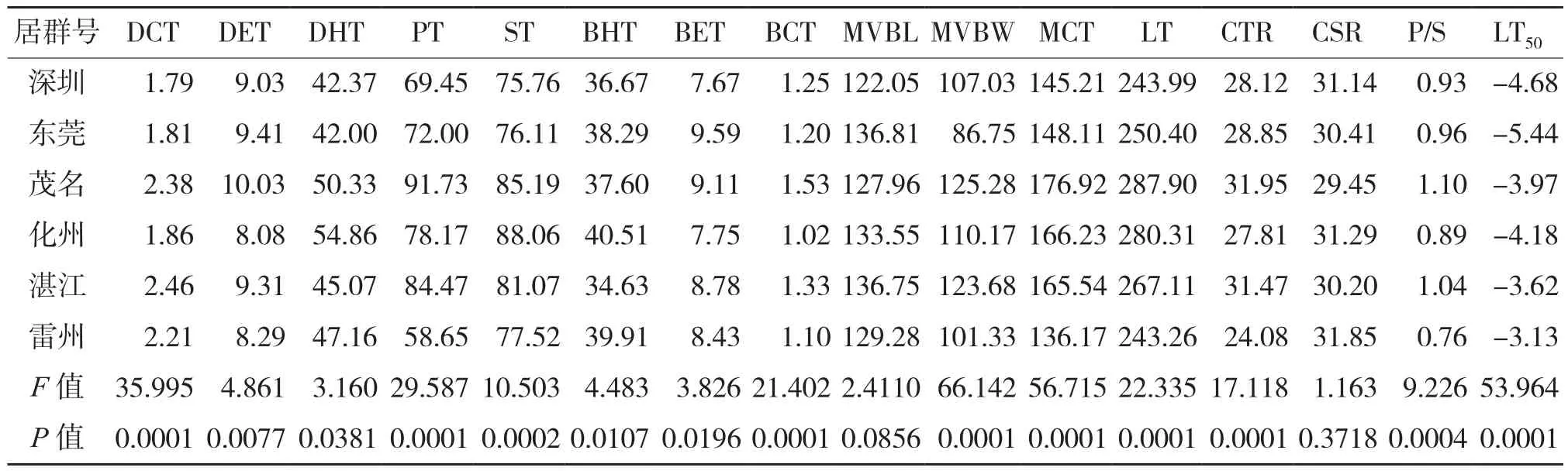
表2 油棕居群的葉片解剖結構指標測定
2.3 油棕植株和居群的葉片解剖結構與其抗寒力的相關性分析
相關系數越大表明關系強度越高。Cohen將相關系數(r)為 r ≥ 0.70、0.69 >r> 0.50、0.49 >r> 0.30和0.29 >r≥ 0.10分別評價為關系強度非常高、高、中等和低4個等級[16]。相關分析結果(表3~表4)表明,無論是油棕植株還是居群,其葉片的中脈維管束寬度與其他解剖結構特性之間均無顯著相關性,居群葉片的上角質層厚度、下內皮層厚度、下表皮層厚度和中脈維管束長度與其他葉片解剖結構特性之間也沒有顯著相關性。油棕植株和居群的葉片上表皮層厚度與下角質層厚度(分別為r= 0.46**和r= 0.93**)和細胞結構緊密度(分別為r= 0.41**和r= 0.80*)均呈顯著正相關關系,但居群的相關性較強;柵欄組織厚度與葉肉細胞厚度、葉片總厚度、細胞結構緊密度和柵海比均呈非常強的正相關關系;海綿組織厚度與葉肉細胞厚度、葉片總厚度和上內皮層厚度呈顯著正相關關系;下角質層厚度與細胞結構緊密度、柵海比顯著正相關,但居群的相關性較強;葉肉細胞厚度與葉片總厚度、細胞結構緊密度顯著正相關;細胞結構緊密度與細胞結構疏松度成顯著負相關(分別為r= - 0.45**和r= - 0.94**)、與柵海比呈顯著正相關(分別為r= 0.88**和r= 0.99**);細胞結構疏松度與柵海比呈顯著負相關(分別為r= - 0.79**和r= - 0.97**)。而油棕植株和居群的葉片解剖結構特征與其所對應的植株和居群的林LT50之間均沒有顯著相關性。
2.4 影響油棕抗寒力的主要葉片解剖結構因子
以LT50作為抗寒力鑒定指標,LT50越小,抗寒力越強。由表5可知,在油棕植株葉片解剖結構特征中,海綿組織厚度對植株LT50的影響最大且有負向直接影響(p2y= - 4.0199),其次為柵/海比(p11y= - 2.341),且葉肉細胞厚度經由海綿組織厚度也具有最大的負向間接影響(r72p2y= - 3.506);而葉片總厚度有最大的正向直接影響(p8y= 2.6694),其次為細胞結構緊密度(p9y=1.369),且葉肉細胞厚度經由葉片總厚度也具有最大的正向間接影響(r78p8y= 2.572)。表明植株葉片的海綿組織厚度和柵/海比越大,植株的LT50越小、抗寒力越強,而葉片總厚度和細胞結構緊密度的影響作用則相反。此外,由于決定系數(0.27839)偏小、剩余通徑系數(0.84948)較大,說明27.8%的LT50表型變異是由表5中所列的結構因子決定的,但未知因素可能占LT50表型變異的72.2%,表明影響LT50的更重要因素尚未考慮在內[17],因此本研究中的葉片解剖結構特征應是油棕植株抗寒力形成的次要因素。
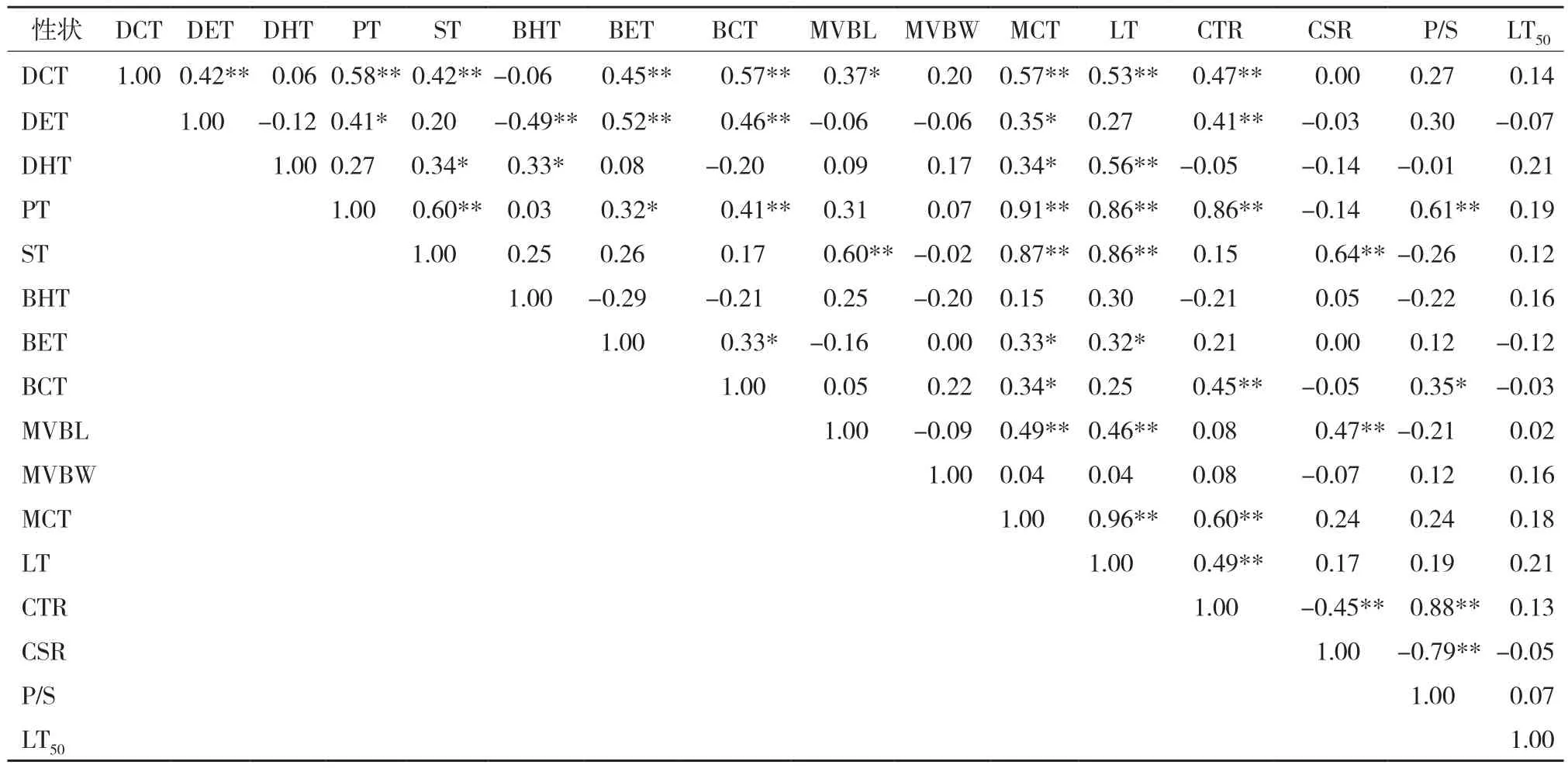
表3 油棕植株抗寒力與葉片解剖結構特征的相關性分析
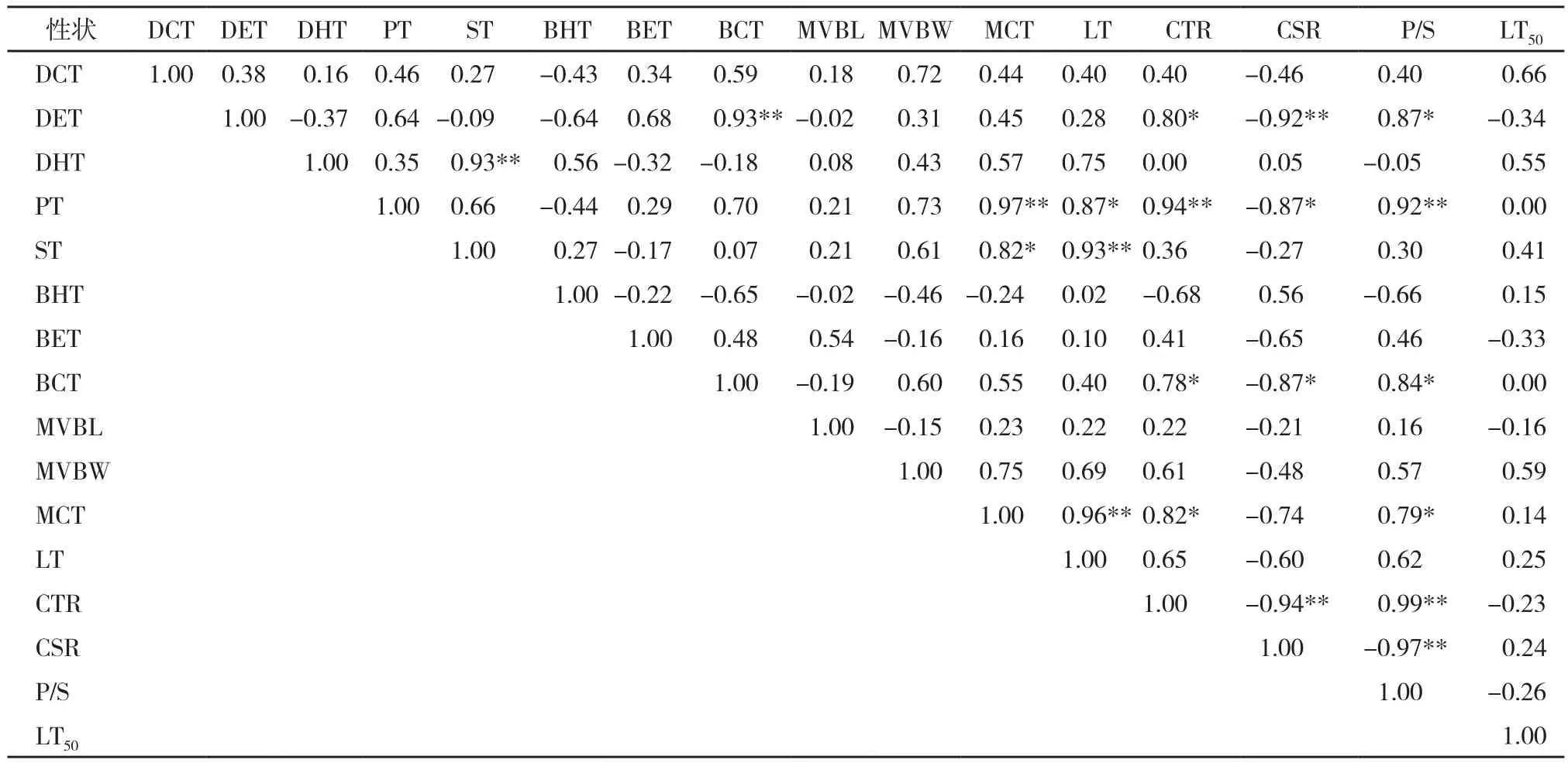
表4 油棕居群抗寒力與葉片解剖結構特征的相關性分析
從表6可以看出,上角質層厚度是影響油棕居群LT50的主要決定因子且具有最大的正向直接作用(p1y=1.0149),其次為細胞結構疏松度(p4y=0.648),影響強度最低的為上內皮層厚度(p2y=0.230),而細胞結構疏松度經由上角質層厚度則具有最大的負向間接影響(r11p1y=-0.463)。維管束長度具有最大的負向直接作用,但影響強度低(p3y=-0.149),而維管束長度經由上角質層厚度則具有最大的正向間接影響(r31p1y=0.178)。表明較低的上角質層厚度和細胞結構疏松度有利于提高油棕居群的抗寒力。

表5 油棕葉片解剖結構特征對植株抗寒力的直接(對角線)和間接(非對角線)影響
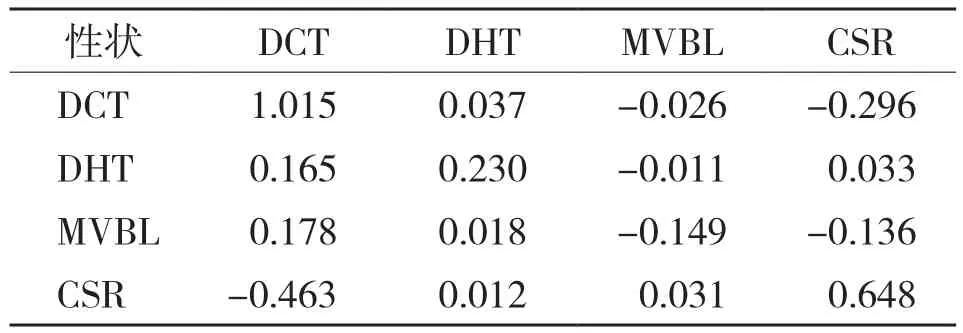
表6 油棕葉片解剖結構特征對居群抗寒力的直接(對角線)和間接(非對角線)的影響
3 結論與討論
低溫影響油棕的生長和產量[3]。低溫脅迫時,植物細胞膜透性增大,細胞內電解質外滲,因此,可通過測定相對電導率的大小來反映植物的抗寒性[6],這在許多植物上得到了應用[18-19]。而葉片是植物進化過程中對環境變化較敏感且可塑性較大的器官,其結構特征最能體現環境因子對植物生長的影響或植物對環境的適應,常被用作植物抗寒性鑒定依據之一,但因不同植物在解剖結構上的抗寒機制不同,因此影響植物抗寒力的主要結構因子也不盡一致[20]。Luis等[10]研究發現溫室和試管生長條件下的油棕實生幼苗的葉片解剖結構發育存在差異,且較大的葉片角質層厚度有利于提高植物的環境適應性。曹紅星等[11]認為葉片總厚度和細胞結構疏松度因低溫脅迫時變化明顯,可作為油棕抗寒種質鑒定的結構指標。馬婷等[18]在比較不同核桃品種的抗寒性時發現,葉片細胞結構疏松度、海綿組織厚度和葉片總厚度越大,核桃抗寒力越強,認為是這評價核桃抗寒性的重要指標。
本研究發現油棕植株之間的葉片解剖結構和LT50均存在顯著差異,但葉片中脈維管束長度和細胞結構疏松度在居群之間則沒有顯著差異。此外,油棕植株葉片中脈維管束寬度以及居群葉片的上角質層厚度、下內皮層厚度、下表皮層厚度、中脈維管束長度和寬度與其他解剖結構沒有顯著相關性,且油棕植株和居群的葉片解剖結構與其所對應的LT50均無顯著相關性。以LT50為抗寒力鑒定指標,LT50越小,抗寒力越強。通徑分析表明,葉片海綿組織厚度是是決定油棕植株抗寒力的主要結構因子(p2y= - 4.0199),海綿組織厚度越大,植株抗寒力越強,但上角質層厚度則是對油棕居群抗寒力起重要決定作用的結構因子(p1y= 1.0149),即較薄的上角質層厚度有利于誘導油棕居群抗寒力的增強,依此可分別篩選出MM2(109.99 μm)和SZ6(51.46 μm)為抗寒性最強(-4.62℃)和抗寒性最弱(-5.00℃)植株,深圳(1.79 μm)和湛江(2.46 μm)為抗寒性最強(-4.68℃)和抗旱性最弱(-3.62℃)居群。而篩選的結果與LT50的實際測定結果不一致,且與其他學者的研究結果不盡相同。Luis等[10]認為,葉片角質層厚度的增加,在植物自身防御與抗逆等方面扮演重要角色,如較厚的角質層限制了病原體的侵染,減少了營養元素以及其他物質的滲出,提高了植物環境適應性。阮志平[12]研究認為,油棕葉片的海綿組織厚度、柵欄組織的層數和厚度、細胞組織緊密度和疏松度以及柵海比等指標都不宜用于鑒定植株抗寒能力,而與柵欄組織分化顯著相關。蔡永立等[21]研究認為,植物葉片的上、下表皮層細胞厚度、柵欄組織和海綿組織厚度以及葉片總厚度的增加是植物對低溫環境的反應。王明啟等[22]通過薔薇屬切片發現,耐寒品種的柵欄組織厚度較大,而不耐寒品種的海綿組織厚度較大。李劍等[23]認為植物葉片解剖結構的細胞組織緊密度越大、細胞組織疏松度越小,其抗寒性越強。鑒于植物的抗寒性是一個復合性狀,不但存在遺傳差異,也易受到環境和栽培條件等影響,因此利用單一指標很難反映植物抗寒性實質,應同時采用多種方法進行相互驗證才能得到比較準確的結論[6]。
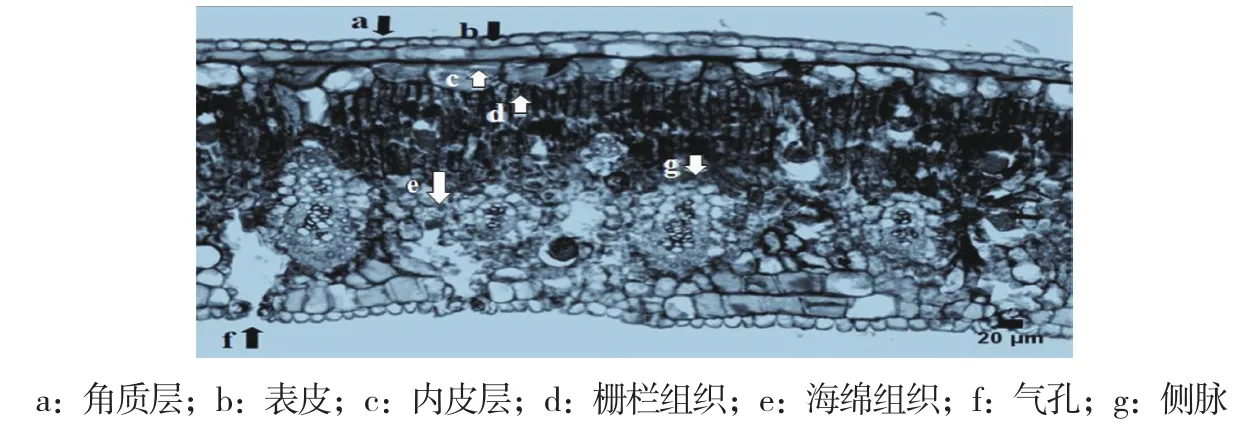
圖1 油棕葉片橫切面(100×)
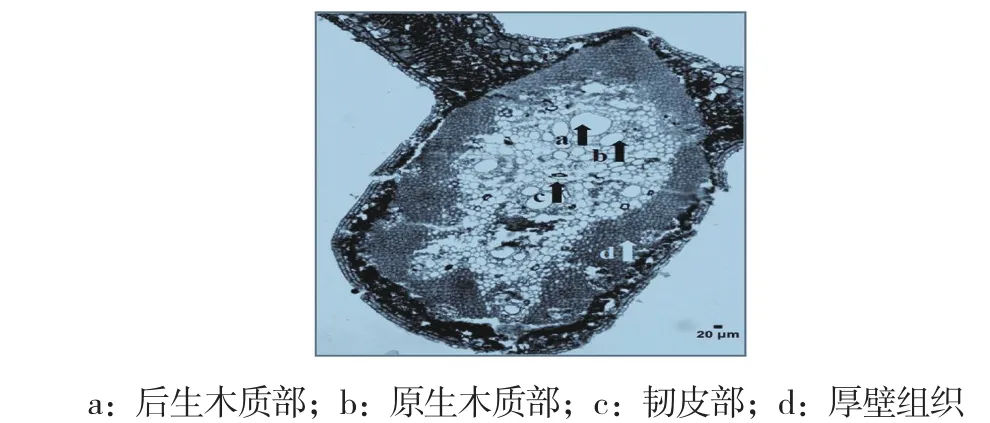
圖2 油棕葉片中脈橫切面(100×)
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
