基因轉染建立高表達轉化生長因子β1大鼠肝星狀細胞的實驗研究
2018-09-20 10:09:36周霞黃效模張一曾德珍
浙江臨床醫學 2018年7期
周霞 黃效模 張一 曾德珍?
肝纖維化的形成機制是一個有眾多因素參與復雜的病理過程,肝纖維化實質是慢性肝病損傷修復反應,肝星狀細胞(HSC)在肝纖維化過程中起核心作用,各種原因肝損傷時,引起肝臟細胞外基質(ECM)合成增加,降解減少,進而造成肝內ECM不斷積聚,最后導致肝纖維化甚至肝硬化。TGFβ1是HSC活化最重要的因子之一[1],目前大多數抗纖維化研究均以HSC為靶標,2017年3月至12月作者通過實驗研究,擬建立高表達TGFβ1的HSC陽性克隆,為肝纖維化的防治提供細胞實驗基礎。
1 材料與方法
1.1 材料 大鼠肝星狀細胞株HSC-T6由武漢協和醫院胃腸病實驗室提供。pcDNA3.1(+)質粒、感受態大腸桿菌DH-5ɑ購自武漢晶賽生物工程技術公司,限制性核酸內切酶 NheI、XbaI、SmaI、Bg1II、EcoRI、T4多聚核苷酸激酶和Taq酶購自TAKARA公司,T4 DNA連接酶購自New England Biolabs,凝膠回收試劑盒和小量質粒提取試劑盒購自寧波中鼎公司,寡核苷酸DNA單鏈由上海博亞生物技術有限公司合成。陽離子脂質體Lipofectamine 2000 購自Invitrogen 公司,TRIzolTM Reagent 購自Invitrogen公司,新生SD大鼠由同濟醫科大學實驗中心提供。
1.2 方法 (1)質粒構建:根據已知的大鼠TGFβ1基因序列合成目的片段引物:TGFβ1引物: P1:5'-GGAATTCGCCACCATGGGCATGCCGCCCTCGGGG CT-3',P2:5'-CCGCTCGAGTCAGCTGCACTTGCAGGA GCG-3'。TGF 1兩端有EcoRI和XhoI酶切位點。以大鼠cDNA為模板擴增TGFβ1基因中1.2kb的目的基因片段,PCR反應條件為:94℃變性5min進入循環過程,94℃變性20s,60℃下退火25s,72℃延伸75s,循環32次后72℃復性3min,凝膠電泳回收TGFβ1產物,限制性內切酶EcoRI和XhoI酶切TGFβ1產物回收1.2kb的酶切產物。(2)表達載體構建:TGFβ1酶切產物通過T4連接酶與EcoRI和XhoI酶切的質粒pcDNA3.1(+)連接,轉化感受態DH-5ɑ,新霉素抗性篩選重組子pcDNA3.1(+)-TGFβ1,酶切鑒定成功后,菌液由上海英俊公司進行測序分析。同時,用含綠色熒光蛋白的pcDNA3.1(+)-EGFP為陰性對照質粒。(3)pcDNA3.1(+)-TGFβ1轉染HSC及G418的篩選:大鼠肝星狀細胞株HSC-T6含15%胎牛血清(FCS)的DMEM液,置于37℃,體積分數為5%CO2孵育箱中培養。轉染前1d,將HSC-T6細胞接種于6孔培養板,每孔2×105個細胞,過夜培養后,將構建的質粒轉染細胞,操作按脂質體Lipofectamine 2000 轉染手冊進行。取一試管加入質粒2μg,用250μl Opti-MEM 培養基混合稀釋,另一試管加入脂質體Lipofectamine 2000 5μl,用250μl Opti-MEM培養基混合稀釋。室溫無菌條件下放置5min,兩試管混合后放置20min,在6孔板中每孔加入量為500μl混合液培養4h,更換培養液繼續培養,48h后計算轉染率,并改用含400μg/mlG418培養基,15%FCS的DMEM培養液2ml,6d后細胞大部分凋死,G418濃度減至200μg/ml維持篩選,殘存細胞約16d后形成陽性克隆,更換為正常培養液繼續培養和傳代。并將細胞分為pcDNA3.1(+)-TGFβ1轉染組,pcDNA3.1(+)-EGFP轉染組,未傳代及轉染細胞為空白組,正常培養傳代21d的空白組細胞為陰性對照組。(4)熒光定量PCR檢測TGFβ1mRNA的表達:篩選陽性克隆后,繼續培養48h,用Trizol試劑盒提取細胞總RNA,熒光定量PCR法分別擴增TGFβ1mRNA。按定量RT-PCR試劑盒說明取1μg總RNA逆轉錄合成cDNA,然后以Sybr Green I作為熒光標記物,在LightCycler熒光實時定量PCR儀(Roche公司,德國)上進行PCR反應。PCR引物:TGFβ1:5'-GAGGCGGTGCTCGCTTTGTA-3',下游引物 5'-TTGTTGCGGTCCACCATTAGC-3'。反應條件如下:預變性94℃ 3min,50個循環中94℃ 30s,53℃30s,72℃ 30s,通過融解曲線分析和電泳確定目的條帶,CT法進行相對定量。(5)Western blot法測定TGF-β1的表達:收集各組細胞,消化后加入預冷的細胞裂解液,置冰上30min,4℃、12000g離心,取上清液進行蛋白分光光度儀定量(考馬斯亮蘭法),每組取20μg蛋白加熱變性后,10%SDS-PAGE電泳分離,電轉移至PVDF膜上,含5%脫脂奶粉的Tris鹽溶液封閉2h,加入1:200稀釋的一抗兔抗TGFβ1多克隆抗體,4℃孵育過夜。用含吐溫-20的Tris鹽溶液洗膜3次,加入1:5000稀釋的辣根過氧化物酶標記的羊抗兔二抗,室溫下孵育1h。化學發光法(ECL)檢測條帶。β-actin作為內參。上述實驗均重復3次,并計算機掃描電泳圖像。
1.3 統計學方法 采用SPSS17.0統計軟件。計量資料用(±s)表示,用方差分析,P<0.05為差異有統計學意義。
2 結果
2.1 重組質粒的鑒定 重組質粒pcDNA3.1(+)-TGFβ1用BamHI酶切,經2%瓊脂糖凝膠電泳后可見1.2kb的清晰條帶,進行測序結果與GenBank所提供的序列完全相同,提示含有TGFβ1基因的pcDNA3.1(+)構建成功。見圖1。
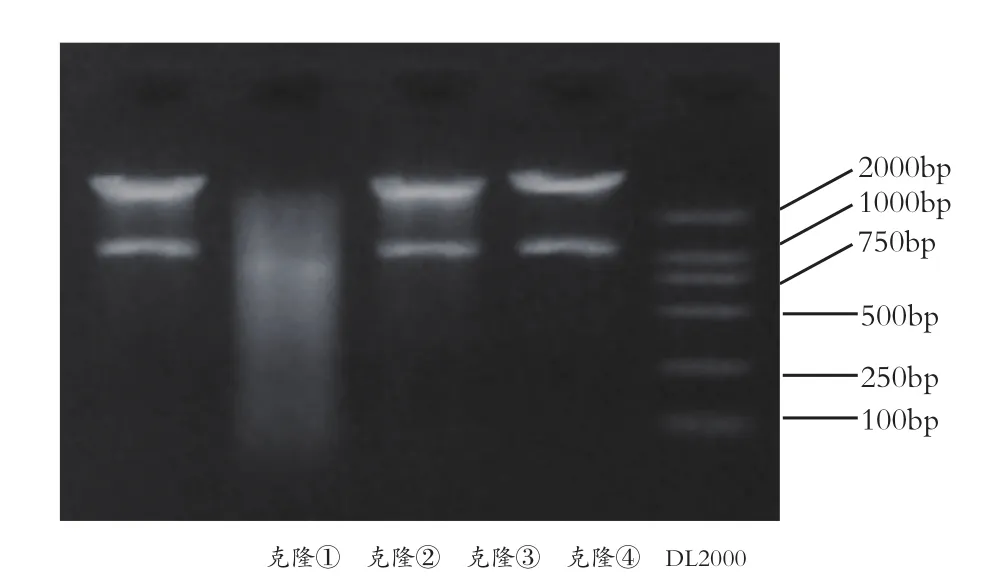
圖1 重組pcDNA3.1(+)-TGFβ1BamHI單酶切鑒定結果
2.2 脂質體對Lipofectamine 2000對細胞毒性作用的G418篩選 應用轉染試劑Lipofectamine 200 pcDNA3.1(+)-TGFβ10轉染HSC-T6細胞后鏡下觀察顯示,在5μl的用量下HSC-T6細胞轉染前、后的形態和數目與對照組及空白組無明顯差異,瞬時轉染48h后測得轉染率為28.2%。篩選結果顯示,G418篩選試劑的最佳濃度為400μg/ml,應用21d后可篩選出穩定表達的細胞克隆,并能進一步培養傳代。見圖2。
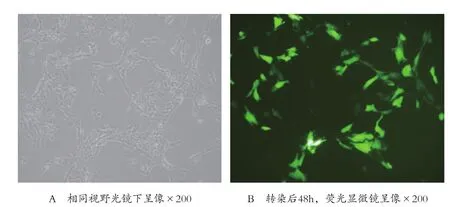
圖2 轉染48h EGFP的表達
2.3 TGFβ1mRNA的表達情況 熒光定量PCR法檢測TGFβ1的mRNA表達,結果顯示:pcDNA3.1(+)-TGFβ1轉 染 組 TGFβ1的 mRNA 表 達 較 pcDNA3.1(+)-EGFP質粒轉染組、空白組及對照組明顯增高[(20.220±0.879)vs(9.609±1.607)vs(5.122±3.512)vs(6.213±0.259),P<0.05]。

圖3 Wester blot 法檢測各組TGFβ1蛋白的表達
2.4 TGFβ1蛋白的表達情況 Western blot印跡法檢測TGFβ1蛋白表達。TGFβ1在SDS-PAGE凝膠電泳中表現為46KD的條帶,實驗顯示:pcDNA3.1(+)-TGFβ1轉染組TGFβ1蛋白的表達較pcDNA3.1(+)-EGFP質粒轉染組、空白組及對照組明顯增高[(0.968±0.822)vs(0.319±0.083)vs(0.211±0.024)vs(0.250±0.086),P<0.05]。見圖 3。
3 討論
肝纖維化指肝臟纖維結締組織的過度沉積,是ECM合成和降解不平衡的結果,是各種慢性肝病向肝硬化發展所共有的病理改變和必經途徑。早期肝纖維化經治療可以逆轉[2],因此深入開展肝纖維化形成機制的研究有十分重要的意義。目前認為HSC的激活是肝纖維化形成機制的中心環節。慢性肝損害過程中,大量細胞因子通過介導細胞-細胞間,基質-細胞間相互作用影響HSC,激活并促使HSC大量合成ECM,導致ECM過分沉積出現肝纖維化。在HSC的激活、致纖維化以及與其他細胞的相互作用的過程中,多種因素參與該過程的調節,細胞因子起重要作用,而TGF-β1是激活 HSC 的最重要促進因子[3]。
TGFβ是一種多功能的多肽類細胞因子,幾乎體內所有細胞都能分泌TGFβ并存在其受體,在細胞的生長調節中起重要作用。自從1983年TGFβ1及隨后其他兩個成員TGFβ2、TGFβ3在哺乳類動物中發現,關于TGFβ的研究也越來越引起人們的重視。TGFβ家族是已知與纖維化形成關系最密切的細胞因子[4],其中,TGFβ1是最重要的致纖維化因子[5],在肝纖維化發展過程中起重要作用。TGFβ1是啟動鄰近靜息態HSC激活和轉化的初始信號之一,Gressner[6]等在肝纖維化形成的3步激活模式中表明TGFβ1的這種初始激活作用,并已被大量研究證實。Kupper細胞、單核細胞、血小板的旁分泌及HSC的自分泌是TGFβ1早期持續增高并使肝纖維化持續發展的重要原因。TGFβ1所介導的信號通道被認為在HSC的活化及ECM的產生起重要作用[7]。
HSC 作為肝纖維化發病的關鍵環節倍受關注,以HSC 為靶標的治療措施逐漸成為抗纖維化的重要方法[8]。因此,構建pcDNA3.1(+)-TGFβ1真核表達載體并轉染HSC,建立高表達TGFβ1的HSC,模擬肝纖維化過程中TGFβ1高表達的體內環境,并以此為細胞為基礎進行下一步研究。本資料表明,pcDNA3.1(+)-TGFβ1真核表達載體構建成功,并可成功轉染HSC,轉染率為28.2%,48h后加入G418篩選,約21d可形成陽性克隆,經熒光定量PCR及Western blot鑒定,克隆細胞高表達TGFβ1mRNA及蛋白,表明高表達TGFβ1的HSC篩選成功。鑒于HSC在肝纖維化發病機制中的主導作用,可應用高表達TGFβ1的HSC開展肝臟疾病,尤其是肝纖維化、肝硬化的研究,可得到更為可信且直接的實驗結果,為肝纖維化的防治提供理論依據。
