森林生態(tài)系統(tǒng)性狀的空間格局與影響因素研究進(jìn)展
——基于中國東部樣帶的整合分析
2018-11-02 06:29:22何念鵬張佳慧劉聰聰王瑞麗徐志偉朱劍興侯繼華于貴瑞
生態(tài)學(xué)報(bào) 2018年18期
何念鵬, 張佳慧, 劉聰聰, 徐 麗, 陳 智, 劉 遠(yuǎn), 王瑞麗, 趙 寧, 徐志偉, 田 靜, 王 情, 朱劍興, 李 穎, 侯繼華, 于貴瑞,*
1 中國科學(xué)院地理科學(xué)與資源研究所 生態(tài)系統(tǒng)網(wǎng)絡(luò)觀測與模擬重點(diǎn)實(shí)驗(yàn)室, 北京 100101 2 中國科學(xué)院大學(xué) 資源與環(huán)境學(xué)院, 北京 100049 3 西北農(nóng)林科技大學(xué) 林學(xué)院, 楊陵 712100 4 中國科學(xué)院西北生態(tài)環(huán)境資源研究院 遙感與地理信息科學(xué)研究室, 蘭州 730000 5 東北師范大學(xué)地理科學(xué)學(xué)院, 長春 130024 6 北京林業(yè)大學(xué) 林學(xué)院, 北京 100083
1 性狀研究進(jìn)展簡述
性狀或功能性狀是生物(植物、動(dòng)物和微生物等)對外界環(huán)境長期適應(yīng)和進(jìn)化后所呈現(xiàn)出來的可量度的特征[1]。植物光合作用是生態(tài)系統(tǒng)初級生產(chǎn)力的重要來源,因此有關(guān)植物性狀的研究一直以來受到人們高度重視,甚至人們潛意識(shí)地將性狀研究等同于植物性狀或植物功能性狀研究[2]。在實(shí)際操作過程中,研究人員更多選擇能體現(xiàn)植物對環(huán)境適應(yīng)并(或)影響植物生產(chǎn)力的部分性狀,如葉片性狀(葉片大小、葉片厚度、比葉面積、氮(N)和磷(P)含量等)和根性狀(根大小、根長度、比根長、N和P含量等)[3]。20世紀(jì)80年代以來,植物性狀研究作為生物學(xué)、地學(xué)和環(huán)境科學(xué)交叉研究的紐帶,成為研究熱點(diǎn),尤其是涉及碳收支0、養(yǎng)分和水分利用的植物性狀更是倍受重視,重點(diǎn)探討了如下幾方面的科學(xué)問題:①植物性狀在不同時(shí)空尺度下的變異規(guī)律及其影響因素;②植物如何通過多種性狀協(xié)同(或權(quán)衡)來優(yōu)化生產(chǎn)力;③如何將器官水平測定的傳統(tǒng)性狀用于解釋自然群落構(gòu)建的機(jī)制;④如何從性狀角度探討陸地生態(tài)系統(tǒng)對全球變化(氣候變化、氮沉降和酸沉降等)的響應(yīng)與適應(yīng)。
近年來,植物性狀研究的豐碩成果主要體現(xiàn)在如下幾個(gè)方面:①種內(nèi)和種間性狀變異及其時(shí)空變異規(guī)律。研究人員從器官-物種-群落-生態(tài)系統(tǒng)、控制實(shí)驗(yàn)-野外調(diào)查-模型模擬等多個(gè)尺度和方法對植物性狀種內(nèi)和種間變異進(jìn)行了探討,具體指標(biāo)如植物個(gè)體大小、種子、葉和根形態(tài)性狀、解剖結(jié)構(gòu)、化學(xué)元素含量等[4]。②多種性狀相互關(guān)聯(lián)關(guān)系及其生產(chǎn)力優(yōu)化機(jī)制。經(jīng)典的研究成果是從資源優(yōu)化利用效率角度發(fā)展了葉經(jīng)濟(jì)學(xué)譜系(Leaf economic spectrum, LES)、根系經(jīng)濟(jì)學(xué)譜系(Root economic spectrum, RES)和木材經(jīng)濟(jì)學(xué)譜系(Wood economic spectrum, WES)[5- 8]。③性狀與種間競爭、自然群落結(jié)構(gòu)維持與功能優(yōu)化間的關(guān)系。在物種水平,研究人員較好地建立了植物性狀與生產(chǎn)力的定量關(guān)系,但如何將個(gè)體水平測定的性狀與天然群落的結(jié)構(gòu)和功能相結(jié)合,仍是該領(lǐng)域的研究熱點(diǎn)與難點(diǎn)[9, 10]。④從植物性狀角度探討植物群落或生態(tài)系統(tǒng)對全球變化的響應(yīng)與適應(yīng)。研究人員利用所測定的部分植物性狀數(shù)據(jù),直接或結(jié)合模型探討全球變化要素(氣候變化、氮沉降和酸沉降等)對生態(tài)系統(tǒng)結(jié)構(gòu)和功能的影響[11-12];其中,利用生態(tài)化學(xué)計(jì)量學(xué)(Ecological stoichiometry)的基本理論,可從N∶P平衡需求的角度揭示生態(tài)系統(tǒng)N和P元素限制狀況以及未來全球變化情景下生態(tài)系統(tǒng)生產(chǎn)力的變化規(guī)律[13- 16]。
整體而言,目前絕大多數(shù)植物性狀研究主要針對優(yōu)勢物種或個(gè)別模式物種,極少充分考慮天然群落物種組成與群落結(jié)構(gòu)的復(fù)雜性[8, 17],因此其結(jié)論是否適用于復(fù)雜的天然群落尚待驗(yàn)證。即使是著名的世界植物屬性數(shù)據(jù)庫(TRY-Plant trait database),也是以收集公開發(fā)表數(shù)據(jù)為基礎(chǔ)建立的,在大多數(shù)植物群落中僅有為數(shù)不多的物種數(shù)據(jù)。此外,TRY數(shù)據(jù)庫的數(shù)據(jù)多集中在植物葉片,非常缺乏葉-枝-干-根-種子等配套的植物性狀數(shù)據(jù)。因此,未來的植物性狀研究應(yīng)該向更系統(tǒng)的指標(biāo)體系、仿真自然群落的模式開展。近幾年,國外科學(xué)家大量提倡從物種推演到群落的觀點(diǎn),但都面臨缺乏一個(gè)系統(tǒng)的實(shí)測數(shù)據(jù)庫來支持研究方法和理論發(fā)展的難題[1, 12]。因此,非常有必要開展系統(tǒng)性調(diào)查與測試,并以此為基礎(chǔ)探討和發(fā)展從物種水平到群落水平的性狀研究的方法論和基本理論。然而,縱觀國內(nèi)外相關(guān)研究,一直缺少貼近天然生態(tài)系統(tǒng)(植物、動(dòng)物、微生物、土壤等)的性狀調(diào)查模式,來引導(dǎo)跨學(xué)科、系統(tǒng)性、集成式調(diào)查和測定;該調(diào)查模式一旦被確立并被廣泛采用,將會(huì)提供各種系統(tǒng)性的性狀調(diào)查數(shù)據(jù)。
本文介紹了在國家基金委重大項(xiàng)目資助下的中國東部南北樣帶(North-South Transect of Eastern China, NSTEC)森林生態(tài)系統(tǒng)性狀(植物、微生物、土壤)的綜合調(diào)查方案及其核心研究結(jié)論,從熱帶雨林至寒溫帶針葉林3700 km樣帶上,選擇了9個(gè)地帶性森林生態(tài)系統(tǒng)開展詳細(xì)的調(diào)查和測定工作,同時(shí)探討了森林生態(tài)系統(tǒng)的性狀研究——如何從物種水平拓展至群落水平?如何建立天然森林群落的性狀與功能的定量關(guān)系?對基于系統(tǒng)性調(diào)查數(shù)據(jù)的性狀研究也進(jìn)行了展望。
2 中國東部南北樣帶(NSTEC)典型森林生態(tài)系統(tǒng)在性狀研究中的潛在意義
根據(jù)國際地圈生物圈計(jì)劃 (International Geosphere-Biosphere Program, IGBP) 的要求,全球變化陸地樣帶都是由一系列沿著某種具有控制陸地生態(tài)系統(tǒng)結(jié)構(gòu)、功能和組成、生物圈-大氣圈間的痕量氣體交換和水分循環(huán)的全球變化驅(qū)動(dòng)力帶狀區(qū)域組成,其長度應(yīng)不小于1000 km,以確保能夠覆蓋氣候和大氣模式以及決策尺度,并有足夠?qū)挾?數(shù)百公里)以涵蓋遙感影像范圍[18-19]。中國東部南北樣帶(NSTEC)于2005年被IGBP確認(rèn)為第15 條標(biāo)準(zhǔn)樣帶;它的主體從中國東部108—118°E 沿經(jīng)線由海南島北上到40°N,然后向東錯(cuò)位10°,再由118—128°E 往北到國界(圖1)。NSTEC具有明顯的熱量梯度與水熱組合梯度,覆蓋了北半球主要森林類型,形成了世界上獨(dú)特且完整的以熱量驅(qū)動(dòng)的植被連續(xù)帶[20-21]。
NSTEC從南到北覆蓋了熱帶山地雨林、亞熱帶闊葉常綠林、溫帶闊葉落葉林、溫帶針闊混交林和寒溫帶針葉林[22- 24];其土壤理化特性隨著緯度升高發(fā)生顯著變化,從低有機(jī)質(zhì)含量的熱帶紅壤逐漸轉(zhuǎn)變?yōu)楦呔暥鹊貐^(qū)高有機(jī)質(zhì)含量的棕壤[25]。由于該樣帶具有明顯的水熱梯度特征,提供了一個(gè)理想的天然實(shí)驗(yàn)室,也有利于探討大尺度下各種生態(tài)系統(tǒng)性狀(植物、微生物和土壤)的變異規(guī)律和影響因素,并可采用空間替代環(huán)境要素變化的模式探討生態(tài)系統(tǒng)性狀的響應(yīng)與適應(yīng)機(jī)制。由于NSTEC覆蓋了北半球幾乎所有的主要森林類型,因此其研究結(jié)論具有全球意義[20]。研究于2013年沿NSTEC選擇了9個(gè)地帶性森林生態(tài)系統(tǒng)(均為原生林)開展系統(tǒng)性狀調(diào)查研究,從南到北分別是尖峰嶺(熱帶山地雨林)、鼎湖山(南亞熱帶常綠闊葉林)、九連山(亞熱帶常綠闊葉林)、神農(nóng)架(北亞熱帶常綠闊葉林)、太岳山(暖溫帶落葉闊葉林)、東靈山(暖溫帶落葉闊葉林)、長白山(溫帶紅松闊葉混交林)、涼水(溫帶紅松闊葉混交林)和呼中(寒溫帶針葉林)(圖1)。調(diào)查樣地跨越33個(gè)緯度(18.7°N—51.8°N)和14個(gè)經(jīng)度(108.9°E—123.0°E)。 研究區(qū)域年均溫(MAT)最低為-3.67 ℃,最高達(dá)到23.15 ℃;年均降水(MAP)最低為472.96 mm,最高為2265.80 mm(表1)。
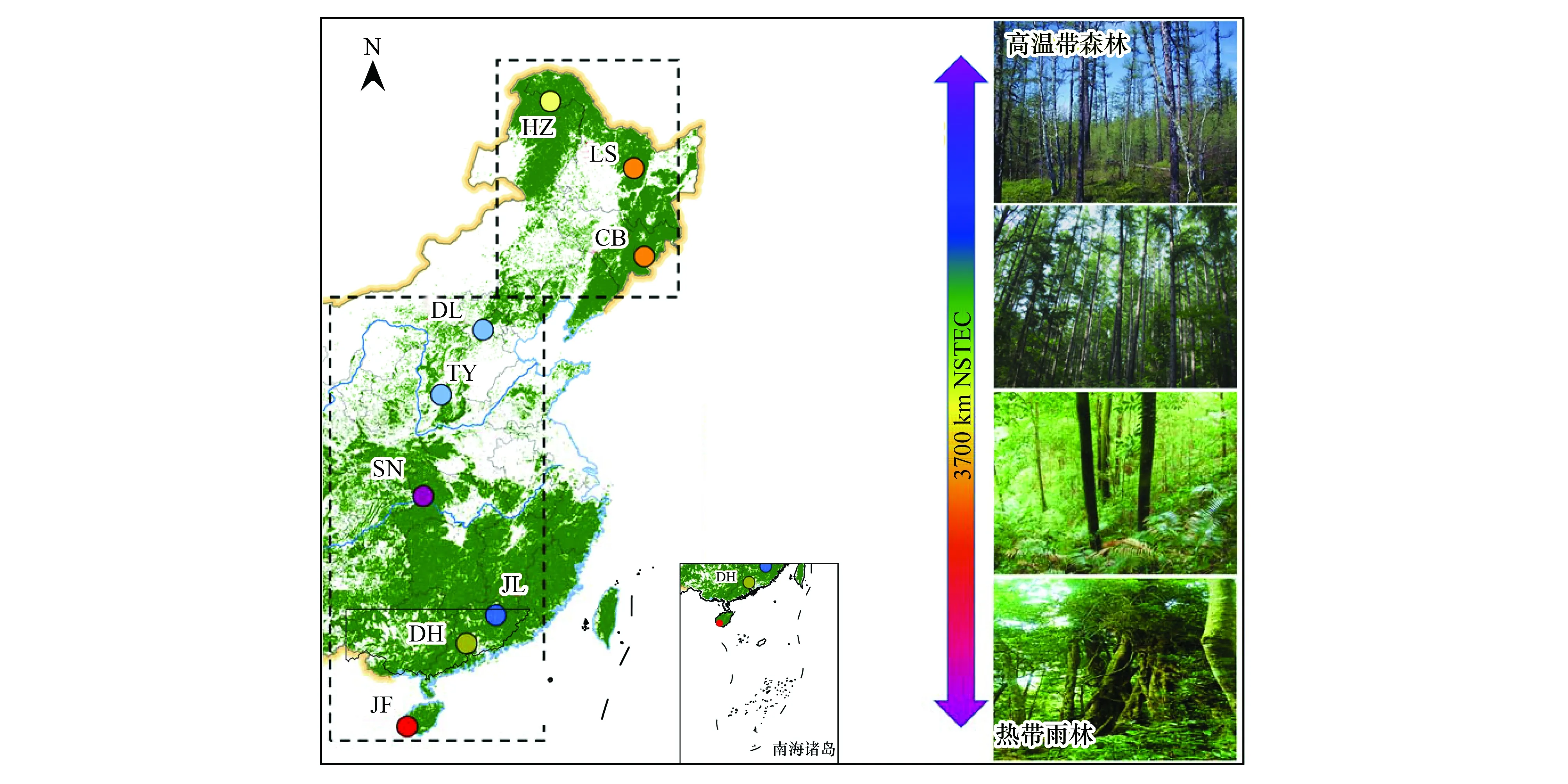
圖1 中國東部南北樣帶及9個(gè)地帶性森林生態(tài)系統(tǒng)空間分布Fig.1 Picture for the North-South Transect of Eastern China (NSTEC) and the nine typical forest ecosystems JF, 尖峰嶺; DH, 鼎湖山; JL, 九連山; SN, 神農(nóng)架; TY, 太岳山; DL, 東靈山; CB, 長白山; LS, 涼水; HZ, 呼中
3 如何基于NSTEC典型森林生態(tài)系統(tǒng)開展系統(tǒng)性的性狀調(diào)查
如何開展系統(tǒng)性的性狀調(diào)查,使其更加接近天然森林生態(tài)系統(tǒng)?為此,引入了圖2的模式。以便所測定的指標(biāo)體系能更接近復(fù)雜的天然森林生態(tài)系統(tǒng),實(shí)現(xiàn)從物種水平(或器官水平)尺度上推至群落水平,探討自然群落水平(或生態(tài)系統(tǒng)水平)性狀與功能的定量關(guān)系。
表1中國東部南北樣帶9個(gè)典型森林生態(tài)系統(tǒng)氣候、植被和土壤基本狀況
Table1Propertiesofclimate,vegetation,andsoilinthenineselectedtypicalforestecosystemsalongtheNorth-SouthTransectofEasternChina(NSTEC)
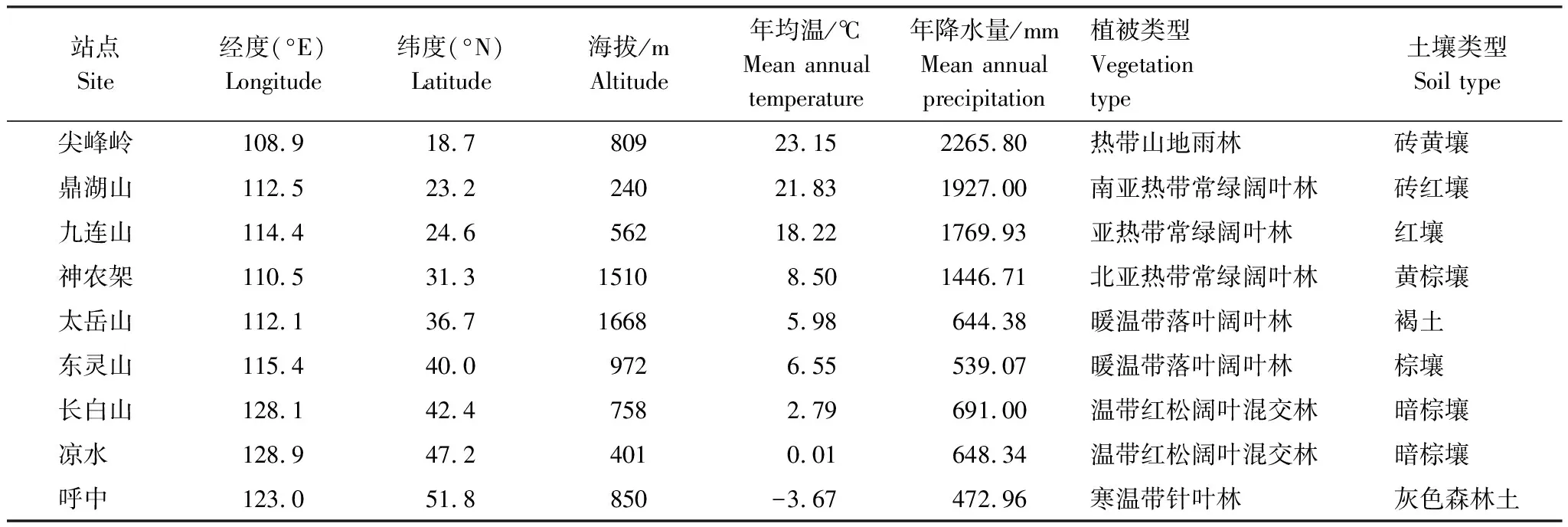
站點(diǎn)Site經(jīng)度(°E)Longitude緯度(°N)Latitude海拔/mAltitude年均溫/℃Mean annual temperature年降水量/mmMean annual precipitation植被類型Vegetation type土壤類型Soil type尖峰嶺108.918.780923.152265.80熱帶山地雨林磚黃壤 鼎湖山112.523.224021.831927.00南亞熱帶常綠闊葉林磚紅壤 九連山114.424.656218.221769.93亞熱帶常綠闊葉林紅壤 神農(nóng)架110.531.315108.501446.71北亞熱帶常綠闊葉林黃棕壤 太岳山112.136.716685.98644.38暖溫帶落葉闊葉林褐土 東靈山115.440.09726.55539.07暖溫帶落葉闊葉林棕壤 長白山128.142.47582.79691.00溫帶紅松闊葉混交林暗棕壤 涼水 128.947.24010.01648.34溫帶紅松闊葉混交林暗棕壤 呼中 123.051.8850-3.67472.96寒溫帶針葉林灰色森林土
土壤類型鑒別參照了中國土壤分類系統(tǒng)(1993)
2013年 7—8月生長旺季,沿NSTEC樣帶選擇了9個(gè)地帶性森林生態(tài)系統(tǒng)為調(diào)查對象(圖1和表1),為降低人為干擾,實(shí)驗(yàn)樣地均設(shè)置于自然保護(hù)區(qū)內(nèi)或野外生態(tài)站的長期監(jiān)測樣地內(nèi),在每類森林生態(tài)系統(tǒng)的典型地段,分別設(shè)置4個(gè)30 m × 40 m喬木調(diào)查樣方、每個(gè)喬木樣方內(nèi)分別設(shè)置2個(gè)5 m × 5 m灌木調(diào)查樣方和4個(gè)1 m × 1 m草本調(diào)查樣方(圖3),按中國生態(tài)系統(tǒng)研究網(wǎng)絡(luò)(CERN)調(diào)查規(guī)范,采用每木檢尺詳細(xì)調(diào)查喬、灌、草的群落結(jié)構(gòu)特征,為后續(xù)采用生物量加權(quán)實(shí)現(xiàn)物種-功能群-群落尺度性狀數(shù)據(jù)拓展提供數(shù)據(jù)支撐[26]。
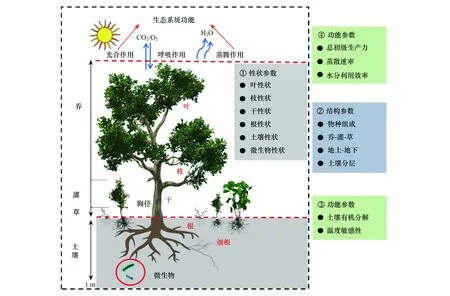
圖2 典型森林生態(tài)系統(tǒng)性狀調(diào)查的邏輯框架Fig.2 Logical frame of the investingated traits (plant, soil microbe, and soil) in typical forest ecosystems
植物樣品采集:①喬木樣品采集:選擇樣地內(nèi)(或周邊)長勢良好的喬木植株,采用吊塔+人工爬樹+高枝剪相結(jié)合模式獲得頂層向陽樹枝;隨后手工摘取無病蟲害的、完全展開的健康葉片,每個(gè)樣地分別形成葉或細(xì)枝混合樣品。在葉片樣品采集的同時(shí),采用生長錐采集樹芯樣品;同時(shí),沿側(cè)根挖掘出完成的細(xì)根根系(從1級到5級根[27])。②灌木樣品采集:對每個(gè)灌木物種,在每個(gè)樣地選擇1—3株健康植株,利用高枝剪或枝剪截取冠層頂端枝條,分別獲得葉和枝條的混合樣品;同時(shí)采用挖掘法獲得灌木根系。③草本樣品采集:在每個(gè)樣地內(nèi),每個(gè)草本物種選取5—30株,采集地上部分(葉、莖)和根系。此外,在草本樣方內(nèi)收集地表凋落物混合樣品。
土壤樣品采集:利用6 cm直徑的土鉆,分0—10 cm、10—30 cm、30—50 cm、50—100 cm四層進(jìn)行土壤取樣。在每個(gè)樣地取30—40鉆土壤樣品,獲得每層土壤樣品>10 kg;隨后,大約4 kg鮮土壤快速冷藏于-20 ℃冰柜,其他樣品遮蔭風(fēng)干。
此次性狀調(diào)查的系統(tǒng)性體現(xiàn)在:①協(xié)同調(diào)查了群落結(jié)構(gòu)、群落內(nèi)每個(gè)物種性狀、微生物屬性、土壤屬性、土壤有機(jī)質(zhì)周轉(zhuǎn)及其溫度敏感性等(表2)。②調(diào)查了樣地內(nèi)出現(xiàn)的所有高等植物種類(喬、灌、草)。比如在呼中寒溫帶針葉林調(diào)查了95個(gè)物種、在尖峰嶺熱帶山地雨林調(diào)查了208個(gè)物種,9個(gè)森林生態(tài)系統(tǒng)共調(diào)查了1177種植物。③開展了喬、灌、草的葉-枝-干-根性狀配套調(diào)查。④葉片性狀包括常規(guī)形態(tài)、氣孔[23, 28]、葉片結(jié)構(gòu)[29, 30]、化學(xué)元素[24, 26, 31, 32]、葉綠素含量[33, 34]、非結(jié)構(gòu)性碳水化合物[35- 37]、葉片熱值等[22, 38]。⑤采用多種方法系統(tǒng)地測定了土壤微生物組成和酶活性[39]。⑥測定了土壤有機(jī)質(zhì)含量及其組分[25];⑦開展了土壤有機(jī)質(zhì)碳氮礦化及其溫度敏感性研究[40- 42]。此次調(diào)查涵蓋了生態(tài)系統(tǒng)植物群落到土壤環(huán)境的整體系統(tǒng)性、植物群落喬木、灌木到草本的群落系統(tǒng)性以及植物個(gè)體葉、枝、干到根的個(gè)體系統(tǒng)性。然而,針對圖2中森林生態(tài)系統(tǒng)冠層界面的功能參數(shù),不同于傳統(tǒng)葉片水平測定的功能參數(shù),更多是指生態(tài)系統(tǒng)尺度的生產(chǎn)力和水分利用效率等,主要是通過渦度相關(guān)技術(shù)來進(jìn)行測量或模型模擬;它并非此次野外調(diào)查數(shù)據(jù)的主要內(nèi)容,該數(shù)據(jù)共享于中國通量網(wǎng)(ChinaFlux)。
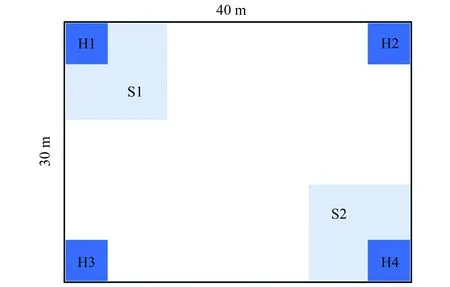
圖3 森林群落調(diào)查樣方的設(shè)置 Fig.3 Design of investigating plots for forest communities喬木樣方面積30 m × 40 m,其中還設(shè)置2個(gè)5 m × 5 m 灌木樣方(S1和S2)和4個(gè) 1 m × 1 m 草本樣方(H1—H4)
除揭示了多個(gè)性狀的空間變異規(guī)律及其影響因素外,基于上述系統(tǒng)性的調(diào)查數(shù)據(jù),發(fā)展了基于植物群落結(jié)構(gòu)和生物量加權(quán)將植物性狀從器官推導(dǎo)至群落的新方法(公式1和2),并率先探討了部分植物性狀與天然森林群落初級生產(chǎn)力(Gross Primary Productivity, GPP)、凈初級生產(chǎn)力(Net Primary Productivity, NPP)、凈生態(tài)系統(tǒng)生產(chǎn)力(Net Ecosystem Productivity, NEP)、水分利用效率(Water Use Efficiency, WUE)等的定量關(guān)系,推動(dòng)了天然森林群落性狀與功能關(guān)系的新認(rèn)識(shí),并提供了可借鑒的方法學(xué)。其中,公式1適于以質(zhì)量(或重量)為基本量綱的性狀指標(biāo),而公式2適用于以葉面積或厚度為量綱的指標(biāo)。通過公式1和2,能把絕大多數(shù)器官水平測定的植物性狀參數(shù)轉(zhuǎn)化為群落水平,并實(shí)現(xiàn)以單位面積為基礎(chǔ)轉(zhuǎn)化,可為探討性狀與天然森林群落功能的關(guān)系奠定堅(jiān)實(shí)的基礎(chǔ),因?yàn)榇蟪叨忍烊簧秩郝渖a(chǎn)力和水分利用效率等功能參數(shù),大多都是以單位土地面積為基礎(chǔ)(通量觀測、遙感觀測或模型模擬)。
(1)
(2)
式中,Traitcomm是群落水平性狀值;trait是物種水平性狀值;n是森林群落中的物種豐富度;j=1, 2, 3, 4分別代表了葉、枝、干和根;Biomassij是指每個(gè)器官生物量在整個(gè)群落生物量中所占的比例;LAIi是指植物群落中i物種的葉面積指數(shù)[23, 28]。
表2中國東部南北樣帶森林生態(tài)系統(tǒng)的性狀指標(biāo)體系與測定方法
Table2TheparametersandmeasuredmethodsfordifferenttraitsinforestecosystemsalongtheNorth-SouthTransectofEasternChina(NSTEC)
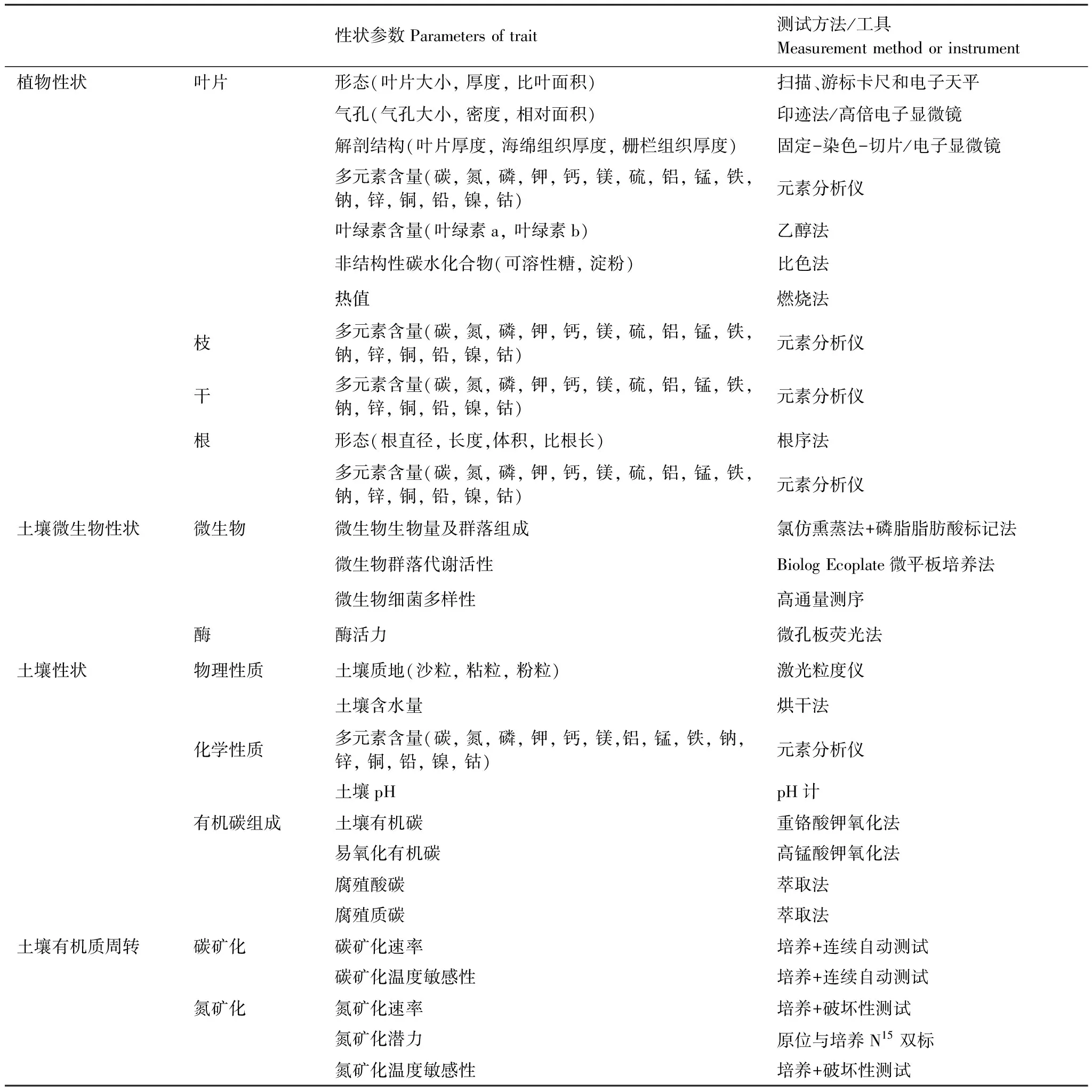
性狀參數(shù)Parameters of trait測試方法/工具M(jìn)easurement method or instrument 植物性狀葉片形態(tài)(葉片大小, 厚度, 比葉面積)掃描、游標(biāo)卡尺和電子天平氣孔(氣孔大小, 密度, 相對面積)印跡法/高倍電子顯微鏡解剖結(jié)構(gòu)(葉片厚度, 海綿組織厚度, 柵欄組織厚度)固定-染色-切片/電子顯微鏡 多元素含量(碳, 氮, 磷, 鉀, 鈣, 鎂, 硫, 鋁, 錳, 鐵, 鈉, 鋅, 銅, 鉛, 鎳, 鈷)元素分析儀葉綠素含量(葉綠素a, 葉綠素b)乙醇法非結(jié)構(gòu)性碳水化合物(可溶性糖, 淀粉)比色法熱值燃燒法枝多元素含量(碳, 氮, 磷, 鉀, 鈣, 鎂, 硫, 鋁, 錳, 鐵, 鈉, 鋅, 銅, 鉛, 鎳, 鈷)元素分析儀干多元素含量(碳, 氮, 磷, 鉀, 鈣, 鎂, 硫, 鋁, 錳, 鐵, 鈉, 鋅, 銅, 鉛, 鎳, 鈷)元素分析儀根形態(tài)(根直徑, 長度,體積, 比根長)根序法多元素含量(碳, 氮, 磷, 鉀, 鈣, 鎂, 硫, 鋁, 錳, 鐵, 鈉, 鋅, 銅, 鉛, 鎳, 鈷)元素分析儀土壤微生物性狀微生物微生物生物量及群落組成氯仿熏蒸法+磷脂脂肪酸標(biāo)記法微生物群落代謝活性Biolog Ecoplate微平板培養(yǎng)法微生物細(xì)菌多樣性高通量測序酶酶活力微孔板熒光法土壤性狀物理性質(zhì)土壤質(zhì)地(沙粒, 粘粒, 粉粒)激光粒度儀土壤含水量烘干法化學(xué)性質(zhì)多元素含量(碳, 氮, 磷, 鉀, 鈣, 鎂,鋁, 錳, 鐵, 鈉, 鋅, 銅, 鉛, 鎳, 鈷)元素分析儀土壤pHpH計(jì)有機(jī)碳組成土壤有機(jī)碳重鉻酸鉀氧化法易氧化有機(jī)碳高錳酸鉀氧化法腐殖酸碳萃取法腐殖質(zhì)碳萃取法土壤有機(jī)質(zhì)周轉(zhuǎn)碳礦化碳礦化速率培養(yǎng)+連續(xù)自動(dòng)測試碳礦化溫度敏感性培養(yǎng)+連續(xù)自動(dòng)測試氮礦化氮礦化速率培養(yǎng)+破壞性測試氮礦化潛力原位與培養(yǎng)N15雙標(biāo)氮礦化溫度敏感性培養(yǎng)+破壞性測試
4 森林植物性狀的空間變異及其影響因素
4.1 葉片性狀
4.1.1 葉片形態(tài)性狀的緯度格局及其影響因素
葉片形態(tài)性狀可以很好地表征植物對外界環(huán)境的響應(yīng)與適應(yīng),備受科研人員的廣泛關(guān)注[8, 17, 32]。通過對NSTEC樣帶9個(gè)典型森林生態(tài)系統(tǒng)1100多種植物葉片性狀數(shù)據(jù)的分析,發(fā)現(xiàn)葉片形態(tài)屬性的緯度變異性在物種水平和群落水平上存在差異[43]。在物種水平上,隨緯度增加,植物葉片大小(LA)、葉片厚度(LT) 和葉片干物質(zhì)含量(LDMC)均顯著降低,葉片比葉面積(SLA)增加;然而,LA和LT沿緯度梯度的變化趨勢卻較弱(R2=0.03-0.04)。然而,利用物種水平數(shù)據(jù)+群落結(jié)構(gòu)數(shù)據(jù)的尺度拓展方法(公式1或公式2),發(fā)現(xiàn)在群落水平上葉片大小(CWM-LA)和比葉面積(CWM-SLA)隨著緯度增加呈現(xiàn)先增加到最大值而后降低的趨勢。群落葉片厚度(CWM-LT)隨緯度增加而增加,群落葉片干物質(zhì)含量(CWM-LDMC)變化趨勢與之相反。相關(guān)研究結(jié)果表明:在探討植物性狀大尺度空間變異規(guī)律和影響因素時(shí),物種水平算術(shù)平均的傳統(tǒng)方法與結(jié)合群落結(jié)構(gòu)和相對生物量(或葉片面積)的權(quán)重加權(quán)法存在很大的差異,有些結(jié)果甚至截然相反;至于物種水平算術(shù)平均法vs.群落結(jié)構(gòu)和生物量加權(quán)法在探討植物性狀格局及其影響因素的優(yōu)缺點(diǎn),會(huì)因研究人員背景或研究內(nèi)容的變化有所變化。
利用巢式方差分析,將物種水平上葉片形態(tài)屬性的緯度變異性區(qū)分為樣地內(nèi)和樣地間的變異性。結(jié)果表明,物種水平的葉片形態(tài)屬性38.2%—50.9%的變異性存在于樣地內(nèi)部共存物種之間,只有16.0%—37.2%的變異性能夠被樣地之間的差異(包括氣候和土壤因子的差異)所解釋。此外,與SLA和LDMC相比,LA和LT中較多的變異性存在于樣地內(nèi)部(> 49%)。植物功能型是LA、LT、SLA和LDMC空間變化的主要影響因素,能夠解釋物種水平的葉片屬性19.43%—41.57%的變異性,而環(huán)境因素僅僅解釋了總變異的1.65%—9.13%。這和全球及區(qū)域尺度上植物功能型對葉片形態(tài)、養(yǎng)分和生理屬性空間變異的解釋程度大于環(huán)境因素的結(jié)論一致[44, 45]。氣候和土壤N含量是影響群落水平的葉片形態(tài)屬性沿緯度變化的主要因素,氣候和土壤養(yǎng)分條件作為主要的環(huán)境濾鏡,通過改變?nèi)郝鋬?nèi)的物種組成和群落結(jié)構(gòu),進(jìn)而決定了葉片形態(tài)屬性的群落加權(quán)平均值在不同環(huán)境條件下的差異[43]。
研究揭示了森林植物葉片形態(tài)性狀的緯度變異規(guī)律與影響因素,但在以下幾方面還需探討:①由于種間變異解釋了葉片功能性狀的大部分變異,物種自身系統(tǒng)發(fā)育是否對葉片功能性狀產(chǎn)生影響以及產(chǎn)生多大影響?②從熱帶雨林到寒溫帶針葉林群落的構(gòu)建機(jī)制是否存在差異?③葉片形態(tài)性狀如何影響天然森林生態(tài)系統(tǒng)的功能?
4.1.2 葉片氣孔性狀的緯度格局及其影響因素
氣孔是植物體內(nèi)水分和二氧化碳與外界環(huán)境進(jìn)行交換的主要通道,其形態(tài)和行為不僅可體現(xiàn)植物自身對環(huán)境的適應(yīng)對策,對全球大氣碳循環(huán)和水循環(huán)也具有深遠(yuǎn)影響[46]。迄今為止,國內(nèi)外科學(xué)家有關(guān)氣孔性狀的研究大多集中在特定區(qū)域的少數(shù)物種[46- 48];在區(qū)域尺度上,天然森林群落的氣孔形態(tài)及其對環(huán)境變化的響應(yīng)和適應(yīng)依然模糊不清。
采用印跡法和高倍電鏡對NSTEC樣帶9個(gè)地帶性森林1100多種植物氣孔性狀進(jìn)行了調(diào)查,具體指標(biāo)包括氣孔密度(SD)和長度(SL)。結(jié)果表明:SD和SL在物種水平上的緯度格局較弱,但在群落水平上隨著緯度的升高而明顯降低,這種變異是由溫度和降水驅(qū)動(dòng)的。在物種水平上,SD與SL呈現(xiàn)負(fù)相關(guān)關(guān)系;在群落水平上,SD與SL卻表現(xiàn)出顯著的正相關(guān)。表明了群落氣孔屬性和物種個(gè)體氣孔屬性兩者對環(huán)境變化的適應(yīng)機(jī)制存在著差異。物種水平上SD與SL之間的權(quán)衡關(guān)系是植物個(gè)體對環(huán)境變化的適應(yīng)進(jìn)化的結(jié)果,而群落水平上的氣孔特征則更多受到群落構(gòu)建過程的影響。在群落水平上的SD能夠解釋NPP空間變異的 51%[23]。為了探究氣孔性狀的影響因素及其與生態(tài)系統(tǒng)水分利用效率的聯(lián)系,提出了氣孔面積指數(shù)(Stomatal area fraction,f,%)[28],并利用群落結(jié)構(gòu)和生物量加權(quán)法計(jì)算了每個(gè)植物群落的氣孔面積指數(shù)(f),發(fā)現(xiàn)f隨著緯度的增加表現(xiàn)出先升高后降低的趨勢,干旱度指數(shù)是主要的驅(qū)動(dòng)因素。同時(shí)還發(fā)現(xiàn)f與生態(tài)系統(tǒng)WUE顯著正相關(guān),表明植物群落通過增大f來優(yōu)化生態(tài)系統(tǒng)的水分利用效率。相關(guān)研究在區(qū)域尺度上為氣孔性狀影響生態(tài)系統(tǒng)水分利用效率提供了直接證據(jù),提高了人們對氣孔性狀適應(yīng)環(huán)境的認(rèn)識(shí)。
通過對NSTEC植物氣孔性狀的系統(tǒng)性調(diào)查,填補(bǔ)了大尺度氣孔數(shù)據(jù)的空白[28]。然而,目前有關(guān)葉片氣孔性狀的分析仍然主要集中在個(gè)體、功能群和群落的格局及影響因素;氣孔作為最重要的植物葉片功能性狀之一,其如何在群落維持和構(gòu)建機(jī)制上發(fā)揮重要的作用仍需進(jìn)一步探討。
4.1.3 葉片解剖結(jié)構(gòu)的緯度格局及其影響因素
葉片解剖結(jié)構(gòu)(上下表皮細(xì)胞、柵欄組織、海綿組織等)具有較大的可塑性,在不同的選擇壓力下會(huì)形成各種適應(yīng)類型[49, 50]。植物同化二氧化碳的同時(shí)也伴隨著蒸騰作用帶來的水分喪失,一方面植物要減少水分喪失,達(dá)到高的水分利用效率,另一方面實(shí)現(xiàn)碳的最大化獲取,充分發(fā)揮光合作用而大量損失水分,導(dǎo)致了低水分利用效率[51]。葉綠素主要分布在柵欄組織和海綿組織中,因此,解剖結(jié)構(gòu)是葉片進(jìn)行光合作用過程中影響光合效率和氣體交換的主要部分[52]。隨著環(huán)境梯度的變化,植物會(huì)如何調(diào)整葉片各個(gè)解剖結(jié)構(gòu)之間的比例關(guān)系,葉片解剖結(jié)構(gòu)是否影響著自然生態(tài)系統(tǒng)的生產(chǎn)力和水分利用效率,值得深入探討。
利用固定-染色-切片技術(shù),對NSTEC的1100多種植物葉片的解剖結(jié)構(gòu)進(jìn)行了測定[29, 30],具體指標(biāo)包括上表皮厚度(AD)、下表皮厚度(AB)、葉厚度(LT)、柵欄組織厚度/海綿組織厚度(PT/ST)、柵欄組織厚度/葉片厚度(CTR)以及海綿組織厚度/葉片厚度(SR)。實(shí)驗(yàn)結(jié)果表明:在物種水平上AD、AB、LT、PT/ST、CTR、SR的均值分別是11.59 μm、8.65 μm、119.66 μm、64.84%、26.82%、48.81%。在功能群水平上,喬木、灌木和草本的6個(gè)解剖結(jié)構(gòu)指標(biāo)之間具有顯著的差異 (P<0.05)。利用群落結(jié)構(gòu)和生物量加權(quán)推導(dǎo)至群落水平后,AD、AB、LT、PT/ST、CTR、SR 均值分別是8.68 μm、3.83 μm、111.55 μm、83.43%、20.89%、28.89%。從物種、功能群到群落水平,解剖結(jié)構(gòu)指標(biāo)均具有顯著的緯度格局,但是不同研究層次,解剖結(jié)構(gòu)指標(biāo)隨緯度的變化趨勢有較大差異。解剖結(jié)構(gòu)受到溫度、降水共同影響,并與干旱指數(shù)顯著相關(guān)。在群落水平,解剖結(jié)構(gòu)與GPP和WUE具有顯著相關(guān)性,其中GPP隨著SR的增加而增加,隨著PT/ST的增加而降低;WUE隨著PT/ST的增加而增加,而隨著SR的增加而降低[30]。
已有研究報(bào)道了從熱帶至寒溫帶森林的植物葉片解剖結(jié)構(gòu)數(shù)據(jù)[30],探究了葉片解剖結(jié)構(gòu)在物種、功能群和群落水平上的緯度分布格局及其影響因素,在區(qū)域尺度上建立了葉片解剖結(jié)構(gòu)與生態(tài)系統(tǒng)生產(chǎn)力和水分利用效率的聯(lián)系,為揭示天然群落中性狀與生態(tài)功能的聯(lián)系提供了新證據(jù)。未來應(yīng)加強(qiáng)系統(tǒng)進(jìn)化對葉片解剖結(jié)構(gòu)的影響研究,從長期適應(yīng)與進(jìn)化角度探討葉片解剖結(jié)構(gòu)的生態(tài)學(xué)意義。
4.1.4 葉片葉綠素含量的緯度格局及其影響因素
光合作用是植物生長的重要能量來源和物質(zhì)基礎(chǔ)。植物光合反應(yīng)可分為原初反應(yīng)、電子傳遞和光合磷酸化、碳同化三個(gè)過程。葉片中的葉綠素a(Chlorophyll a, Chl a)和葉綠素b(Chlorophyll b, Chl b)是原初反應(yīng)的必要條件;然而二者功能卻又有所不同。大多數(shù)Chl a和所有Chl b都能夠吸收太陽光(Chl a主要吸收紅橙光,Chl b主要吸收藍(lán)紫光),但是只有少量處于激發(fā)態(tài)的Chl a能在原初反應(yīng)中傳遞電子,并將光能轉(zhuǎn)化成電能,從而為電子傳遞和光合磷酸化以及碳同化做準(zhǔn)備。據(jù)此推測:葉綠素總量(Chlorophyll content, Chl a+b)以及Chl a 與Chl b 比值(Chl a/b)直接或間接地影響植物的光合效率,是反映植物光合能力的重要指標(biāo)[53]。鑒于葉綠素對植物光合作用的重要意義,科學(xué)家們對葉綠素?zé)晒鈁54]、葉綠素的合成分解[55, 56]等進(jìn)行了大量研究。然而,目前對于葉片葉綠素含量的研究大多停留在區(qū)域優(yōu)勢物種或某物種葉綠素的影響因素[57, 58]等,對于大尺度群落葉綠素的研究鮮有報(bào)道。
利用經(jīng)典的乙醇法提取,測定了NSTEC上9個(gè)地帶性森林群落內(nèi)1100多種植物葉片葉綠素含量(Chl a、Chl b、Chl a+b、Chl a/b),分析發(fā)現(xiàn)葉綠素在物種間和不同生活型間都存在顯著差異;在物種水平時(shí),葉片葉綠素含量僅存在較弱的緯度格局。進(jìn)一步分析發(fā)現(xiàn),葉綠素幾乎不受系統(tǒng)發(fā)育、氣候、土壤的影響,較大的種間變異是導(dǎo)致物種水平葉綠素微弱緯度格局的重要原因。然而,當(dāng)利用群落結(jié)構(gòu)和生物量數(shù)據(jù)加權(quán)法拓展后,發(fā)現(xiàn)群落水平的葉片葉綠素含量隨緯度的升高而降低;進(jìn)一步分析發(fā)現(xiàn)Chl與GPP顯著正相關(guān)(R2=0.32),即Chl一定程度上能夠反映生態(tài)系統(tǒng)GPP,但其作為單一性狀對GPP空間變異的解釋度遠(yuǎn)低于先前人們的預(yù)期。相關(guān)研究從群落的角度揭示了自然群落葉綠素緯度變異規(guī)律、影響因素及其與GPP的定量關(guān)系,為植物功能性狀與功能的研究提供了一個(gè)范例[33-34],也為生態(tài)模型構(gòu)建與優(yōu)化提供了理論依據(jù)和重要參數(shù)。
近年來,許多學(xué)者用葉綠素含量建立了很好的光合模型,并且發(fā)現(xiàn)用葉綠素代替光合效率比傳統(tǒng)上用N含量代替光合效率具有更好的效果[53]。然而,類似模型都是將對少數(shù)物種的實(shí)驗(yàn)結(jié)果直接應(yīng)用于復(fù)雜的自然群落,需要來自群落水平的性狀與功能數(shù)據(jù)的實(shí)驗(yàn)驗(yàn)證;此外,還應(yīng)考慮不同區(qū)域環(huán)境異質(zhì)情況下,葉綠素與GPP定量關(guān)系的變異規(guī)律等。
4.1.5 葉片非結(jié)構(gòu)性碳水化合物的緯度格局與影響因素
碳水化合物是植物光合作用的重要產(chǎn)物,常分為結(jié)構(gòu)性碳水化合物和非結(jié)構(gòu)性碳水化合物(Non-structural carbohydrates, NSC)。NSC主要用于植物生理代謝活動(dòng),其含量的變化很大程度的影響著植株代謝和生長過程、植物碳吸收與碳消耗之間的平衡關(guān)系和植物對外界環(huán)境變化的重要適應(yīng)策略[59]。在垂直地帶性上,溫度驅(qū)動(dòng)機(jī)制下的林線形成(溫度對植物葉片非結(jié)構(gòu)性碳水化合物的影響)有兩種對立的生物生理假說——碳限制假說[60-61]和生長限制假說[62-63]。然而,在大尺度的自然森林生態(tài)系統(tǒng)中,如沿著不同氣候帶(從熱帶到寒溫帶、隨著溫度的降低),植物葉片NSC的變化趨勢會(huì)支持何種假說,不得而知。
以NSTEC樣帶上9個(gè)地帶性森林生態(tài)系統(tǒng)1100多種植物葉片為對象,測定了其可溶性糖和淀粉含量 (NSC=可溶性糖+淀粉)。在物種水平,植物葉片可溶性糖、淀粉和NSC的變化范圍分別為 6.48—218.45 mg/g、7.56—200.41 mg/g 和19.47—379.72 mg/g,平均值分別為45.70、47.50 和93.20 mg/g。此外,不同生活型間葉片NSC差異顯著,草本葉片可溶性糖、淀粉和NSC最高,分別為49.89、60.12和110.00 mg/g;落葉喬木葉片可溶性糖、淀粉和NSC含量顯著高于常綠喬木葉片(P<0.05);針葉喬木的葉片可溶性糖、淀粉和NSC含量顯著高于闊葉喬木葉片。植物葉片可溶性糖、淀粉和NSC含量具有明顯的緯度地帶性分布規(guī)律,沿緯度帶從南到北呈遞增趨勢。然而, 葉片可溶性糖和淀粉的分配策略(比值)沿緯度沒有明顯的變化趨勢,與溫度和降水之間也無顯著相關(guān)性[35- 37]。
相關(guān)研究探討了植物葉片非結(jié)構(gòu)性碳水化合物及其組分的空間格局及控制因素,對了解氣候變化對樹木生理生態(tài)過程的響應(yīng)和適應(yīng)機(jī)制具有一定的意義。未來還應(yīng)加強(qiáng)NSC在不同器官間的分配特征研究,應(yīng)考慮多種因素如氣候、植被和土壤對植物葉片NSC的影響機(jī)制;此外,在復(fù)雜的天然森林群落中,系統(tǒng)發(fā)育、光合途徑對植物葉片NSC的影響也需要進(jìn)一步闡述。
4.2 細(xì)根性狀的緯度格局及其影響因素
細(xì)根在植物水分和養(yǎng)分吸收中具有重要作用,由于野外取樣和測定難度較大,有關(guān)細(xì)根的研究仍然不足[27];目前關(guān)于細(xì)根的功能性狀之間是否存在類似于葉經(jīng)濟(jì)學(xué)譜的規(guī)律仍存在激烈爭論(圖4)。根系經(jīng)濟(jì)學(xué)譜理論(RES)認(rèn)為:細(xì)根中與資源獲取和資源貯存相關(guān)的性狀之間存在權(quán)衡關(guān)系。在RES的一端是快速回報(bào)的物種,具有低的根直徑(RD)、高的比根長(SRL)和根氮含量(RN),同時(shí)具有高的根系呼吸速率和短的壽命;相反的特征存在于慢速回報(bào)的物種內(nèi)。然而,根的多維度假說認(rèn)為,根特征之間不存在唯一的權(quán)衡關(guān)系,(與葉片相比)根系特征受到更為復(fù)雜的環(huán)境限制、具有更高的系統(tǒng)發(fā)育保守性,同時(shí)還會(huì)受到菌根侵染的影響。此外,由于細(xì)根的劃分標(biāo)準(zhǔn)存在差異,也阻礙了對細(xì)根性狀變異及其相互關(guān)系的深入研究;近年來研究提出和發(fā)展了“根序法”和“功能劃分法”,極大推動(dòng)了細(xì)根的研究工作[27, 64]。
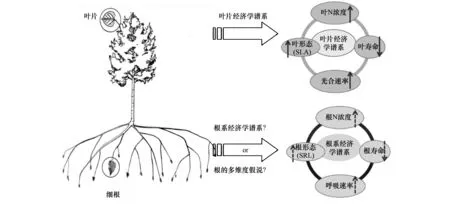
圖4 細(xì)根功能性狀之間關(guān)系的相關(guān)假說Fig.4 Main hypotheses on the traits of fine foot
在NSTEC的9個(gè)地帶性森林中選擇了群落內(nèi)的常見物種(包括喬木、灌木和草本植物),開展了較系統(tǒng)的一級根的功能性狀測定[65]。根據(jù)主成分分析(PCA),在物種和群落水平上均發(fā)現(xiàn)了2個(gè)主成分軸(PC1和PC2)。其中,PC1主要與群落水平比根長(CWM_SRL)和根直徑(CWM_RD)相關(guān),代表了根厚度的變化,PC2主要與根養(yǎng)分性狀相關(guān)。此外,不同氣候區(qū)域的細(xì)根功能性狀存在差異。亞熱帶地區(qū)的植物細(xì)根傾向于具有細(xì)且長的根,而北方針葉林地區(qū)的植物根系具有相反的功能性狀。一些溫帶森林(如長白山)中的物種具有高的氮含量和低的碳氮比。
細(xì)根形態(tài)和化學(xué)指標(biāo)之間的獨(dú)立性支持了細(xì)根的多維度性假說。PC1主要代表了根形態(tài)性狀(如RD和SRL)的變異性;PC2與根的養(yǎng)分性狀相關(guān),這反映了細(xì)根從土壤中吸收養(yǎng)分的重要性。同時(shí),在物種水平上與根形態(tài)性狀相比,根的養(yǎng)分性狀表現(xiàn)了較低的系統(tǒng)發(fā)育保守性和對土壤環(huán)境更高的可塑性[66]。進(jìn)一步分析發(fā)現(xiàn):系統(tǒng)發(fā)育作用能夠解釋木本和草本植物51.4%和49.4%的變異性;并且大多數(shù)變異性發(fā)生在分支水平上(clade),反映了不同進(jìn)化時(shí)期系統(tǒng)發(fā)育分支的根性狀的巨大差異。與物種水平不同,群落水平的細(xì)根性狀變異性受到了環(huán)境濾鏡和生物之間相互作用的雙重影響,這些因素最終決定了群落內(nèi)的物種組成和功能性狀分布[67]。例如,在群落水平上,隨著年均溫增加,PC1先降低,而后略微增加;相反的格局表現(xiàn)在PC2和土壤磷含量的關(guān)系中。
目前,大多數(shù)研究只涉及根的形態(tài)和養(yǎng)分性狀,而對于更能準(zhǔn)確地刻畫根系功能的解剖結(jié)構(gòu),尤其是涉及真菌侵染方面的性狀,以及和根系構(gòu)建特征相關(guān)的性狀(如分支強(qiáng)度)還有待加強(qiáng)。部分研究已經(jīng)表明細(xì)根的菌根侵染率和分支強(qiáng)度是細(xì)根資源吸收策略的重要組成部分[68, 69],因此,未來需要將菌根特征和根的構(gòu)建特征考慮在內(nèi),擴(kuò)展根性狀的研究維度,深入探究根系對環(huán)境的適應(yīng)策略。
4.3 根-葉性狀關(guān)聯(lián)性
葉片與細(xì)根分別是植物體地上和地下最活躍的器官,理論上其性狀應(yīng)存在著密切聯(lián)系[70];因此,理解葉片和細(xì)根功能性狀之間的關(guān)系,能夠?yàn)橹参镔Y源利用策略和生態(tài)系統(tǒng)過程模型優(yōu)化提供重要依據(jù)。然而,目前對于植物地上/地下器官功能性狀之間的關(guān)聯(lián)關(guān)系存在較大爭議。一種假說認(rèn)為,植物各器官功能性狀之間的關(guān)系是一維的。外界環(huán)境的選擇壓力和生物物理限制共同導(dǎo)致了植物功能性狀聚集在從“快”到“慢”的策略軸上(即資源獲取-保守策略)[70]。另一種假說則認(rèn)為,植物不同的器官由于所受到的環(huán)境限制和行使的功能均存在差異,因而會(huì)獨(dú)立進(jìn)化來適應(yīng)不同的環(huán)境條件。因此,植物不同器官功能性狀之間的關(guān)系是多維度的[66, 71],這有利于提高群落內(nèi)物種共存和生態(tài)系統(tǒng)結(jié)構(gòu)與功能的穩(wěn)定性。
利用在NSTEC獲取的木本和草本植物的葉片和細(xì)根性狀,根據(jù)根和葉養(yǎng)分獲取和形態(tài)特征對應(yīng)角度,選擇了5組性狀來探討根-葉性狀關(guān)系。5組分別為:葉片厚度 vs 細(xì)根直徑、比葉面積vs 比根長、葉片組織密度vs細(xì)根組織密度、葉片碳含量vs 細(xì)根碳含量、葉片氮含量vs 細(xì)根氮含量。實(shí)驗(yàn)結(jié)果表明:形態(tài)特征之間的關(guān)系取決于是否考慮系統(tǒng)發(fā)育因素,而葉片氮和細(xì)根氮含量之間存在緊密的正相關(guān)關(guān)系[72]。因此,根養(yǎng)分特征可以作為連接不同植物器官以及植物與環(huán)境之間關(guān)系的橋梁。葉片和細(xì)根之間形態(tài)特征的解耦可能與地上和地下不同的選擇壓力有關(guān)。葉片特征受到最大化光合速率且最小化水分損失的驅(qū)動(dòng),因此葉片特征存在一維的權(quán)衡關(guān)系。與葉片不同,根性狀受到了更為復(fù)雜的環(huán)境因子(土壤養(yǎng)分和物理?xiàng)l件)和生物因素(菌根侵染和其他根系微生物)的影響[71],導(dǎo)致了葉片和細(xì)根的形態(tài)特征的解耦。
同時(shí),利用NSTEC地帶性森林生態(tài)系統(tǒng)中植物葉片和細(xì)根的16種元素含量碳(C)、氮(N)、鉀(K)、鈣(Ca)、鎂(Mg)、硫(S)、磷(P)、鋁(Al)、錳(Mn)、鐵(Fe)、鈉(Na)、鋅(Zn)、銅(Cu)、鉛(Pb)、鎳(Ni)、鈷(Co),探討了植物葉片和細(xì)根多元素協(xié)同變異規(guī)律。與細(xì)根相比,葉片中具有較高含量的大量元素(N,K,Ca,Mg,S和P,除C以外),而細(xì)根中具有較高含量的微量金屬元素(Al,Fe,Na,Zn,Cu,Pb,Ni和 Co,除了Mn)[32]。進(jìn)一步分析發(fā)現(xiàn),植物譜系是影響植物葉片和根系元素含量變異的重要因素。植物分類(種科目水平的疊加)對葉片和細(xì)根元素含量變異的解釋率均值分別為48.9%和42.6%。植物分類對葉片和細(xì)根中大量元素(C,N,K,Ca,Mg,S和P)的解釋率分別達(dá)到63.7% 和 61.4%,而對微量元素(Al,Mn,Fe,Na,Zn,Cu,Pb,Ni和Co)的解釋率分別為37.6%和28.0%。此外,植物分類對元素變異的解釋率隨著元素含量的升高而升高,并且在葉片和細(xì)根中存在相同的趨勢。整體而言,與微量元素相比,大量元素的變異受到環(huán)境的影響較小。葉片N,K,Fe,Na和細(xì)根Pb,Ni變異較大地受到降水的影響,而葉片P和細(xì)根P,Al含量變化更多的受溫度的影響。其中,葉片部分與之前對于葉片生物地理格局受到降水和溫度影響的研究結(jié)果相類似[73, 74]。
由于葉片和地下根系的功能性狀均存在較大的系統(tǒng)發(fā)育保守性,加之不同研究區(qū)域或物種選擇的差異,導(dǎo)致根葉性狀關(guān)系的相關(guān)研究結(jié)論存在較大爭議。此外,葉片和細(xì)根性狀之間的關(guān)聯(lián)關(guān)系將在不同植物科或分支上存在差異,還有待進(jìn)一步驗(yàn)證;此外,未來還應(yīng)利用植物譜系結(jié)構(gòu)更深入地探討植物營養(yǎng)獲取策略的進(jìn)化格局及其驅(qū)動(dòng)機(jī)制[31]。
4.4 植物C、N、P化學(xué)計(jì)量特征及其分配策略
生態(tài)化學(xué)計(jì)量學(xué)是研究生物多種化學(xué)元素平衡的科學(xué),它通過元素比例在各個(gè)生命過程及生物地球化學(xué)循環(huán)中的數(shù)量關(guān)系,來認(rèn)識(shí)和解決不同層次(基因,細(xì)胞,器官,有機(jī)體,群落,生態(tài)系統(tǒng))的生態(tài)問題[75]。目前,已經(jīng)從物種水平到區(qū)域尺度上對葉片C、N、P元素分配規(guī)律進(jìn)行了大量研究[8, 74, 76]。但受測試難度和經(jīng)費(fèi)的限制,真正基于植物群落水平系統(tǒng)測試數(shù)據(jù)來開展相關(guān)研究的報(bào)道非常少,尤其是區(qū)域到全球尺度。在復(fù)雜的天然森林群落中,三個(gè)假設(shè)仍待系統(tǒng)的實(shí)測數(shù)據(jù)來證實(shí);它們分別是:①元素分配假說(Element Allocation Hypothesis),即越活躍器官被分配更多的營養(yǎng)元素以實(shí)現(xiàn)功能最大化;②元素可塑性假說(Element Plasticity Hypothesis),即地下器官因其更復(fù)雜的環(huán)境因子和生物因素的影響而具有更高的元素可塑性;③ N-P異速分配假說 (N-P Allometry Hypothesis),即N-P異速分配指數(shù)在不同植物器官或不同功能群間存在一定的保守性。
利用NSTEC地帶性森林群落中1100多種植物葉-枝-干-根的測定數(shù)據(jù),發(fā)現(xiàn)葉片(植物器官中普遍認(rèn)為是最活躍的)的N、P含量以及N∶P的值均高于枝(或莖)和根,一定程度上驗(yàn)證元素分配假說[77]。對不同功能群植物而言,N和P含量表現(xiàn)為草本高于木本,N∶P則表現(xiàn)為相反的結(jié)果,在一定程度上反映草本和木本不同的生存策略。如果用變異系數(shù)來表征元素可塑性,物種間N和P的變異系數(shù)均表現(xiàn)為根高于葉和枝,即根元素可塑性更高,與元素可塑性假說一致。對于N-P異速分配策略研究,尺度選擇是一個(gè)非常重要的影響因素;在大尺度上,N-P異速分配斜率均小于1且在不同器官間或不同功能群間沒有顯著差異,證明了其較強(qiáng)的保守性(N-P異速分配假說);然而,在站點(diǎn)尺度,異速分配斜率有呈現(xiàn)大于1的情況[77]。基于長白山垂直樣帶的相關(guān)數(shù)據(jù),葉片、莖、細(xì)根的N-P異速分配關(guān)系在不同生長型植物中存在差異[24]。在垂直梯度上,葉片、莖和細(xì)根的N、P分配對環(huán)境變化的響應(yīng)一致,反映了植物各器官共存的基本機(jī)理。
在探討植物元素化學(xué)計(jì)量學(xué)和分配策略時(shí),還應(yīng)在更大的范圍對這三個(gè)假說進(jìn)行驗(yàn)證,并探討植物多元素的分配策略。此外,鑒于尺度選擇對研究結(jié)果的巨大影響,未來在進(jìn)行尺度擴(kuò)展時(shí)需要考慮其合理性并選擇適當(dāng)?shù)姆椒ā?/p>
4.5 森林生態(tài)系統(tǒng)C、N、P化學(xué)計(jì)量特征及其緯度格局
現(xiàn)階段,生態(tài)化學(xué)計(jì)量學(xué)的研究大多聚焦于物種或植物器官,對土壤和微生物也有一定研究;但由于天然森林群落結(jié)構(gòu)的復(fù)雜性以及葉-枝-干-根的變異性,對植物群落C、N、P化學(xué)計(jì)量特征的研究十分缺乏。一方面造成生態(tài)化學(xué)計(jì)量學(xué)從分子到全球的聯(lián)系難以完整,另一方面使相關(guān)生態(tài)模型因難以獲得不同區(qū)域的相關(guān)數(shù)據(jù)而不能得到優(yōu)化。此外,目前對群落化學(xué)計(jì)量特征的研究大多是以優(yōu)勢種為測定對象(或收集公開數(shù)據(jù)),再通過物種水平直接算數(shù)平均法推導(dǎo),這種做法是否能真實(shí)反映復(fù)雜的天然森林群落還有待實(shí)驗(yàn)數(shù)據(jù)驗(yàn)證。
已有研究獲取了NSTEC樣帶9個(gè)典型森林所有物種葉-枝-干-根、土壤等的C、N、P含量數(shù)據(jù),采用群落結(jié)構(gòu)和生物量加權(quán)法分析中國森林生態(tài)系統(tǒng)不同組分的C∶N∶P 化學(xué)計(jì)量特征[26]。該研究基于實(shí)測數(shù)據(jù)在區(qū)域尺度給出了生態(tài)系統(tǒng)不同組分以及植物不同器官的C∶N∶P化學(xué)計(jì)量特征(圖5),并發(fā)現(xiàn)C∶P與N∶P有隨緯度增加而減小的趨勢。經(jīng)過對氣候(溫度和降水)以及土壤養(yǎng)分(N、P含量)冗余分析發(fā)現(xiàn),氣候和土壤共同影響N∶P的變異,其中氣候是主要影響因素。通過對N∶P比值分析,結(jié)合N∶P >16為磷限制的假設(shè)[78],發(fā)現(xiàn)中國森林在南方受到磷元素限制更為突出。此外,利用內(nèi)穩(wěn)性模型,計(jì)算了植物不同器官N∶P的內(nèi)穩(wěn)性指數(shù),發(fā)現(xiàn)葉片的元素內(nèi)穩(wěn)性高于其他器官,說明越活躍的器官元素穩(wěn)定性越高。需要指出的是:群落結(jié)構(gòu)和生物量加權(quán)所獲得的植物C∶N∶P化學(xué)計(jì)量特征,與物種水平直接算術(shù)平均值差異很大;此外,與國際上通過收集數(shù)據(jù)評估的結(jié)論相比,也存在很大差異[26]。
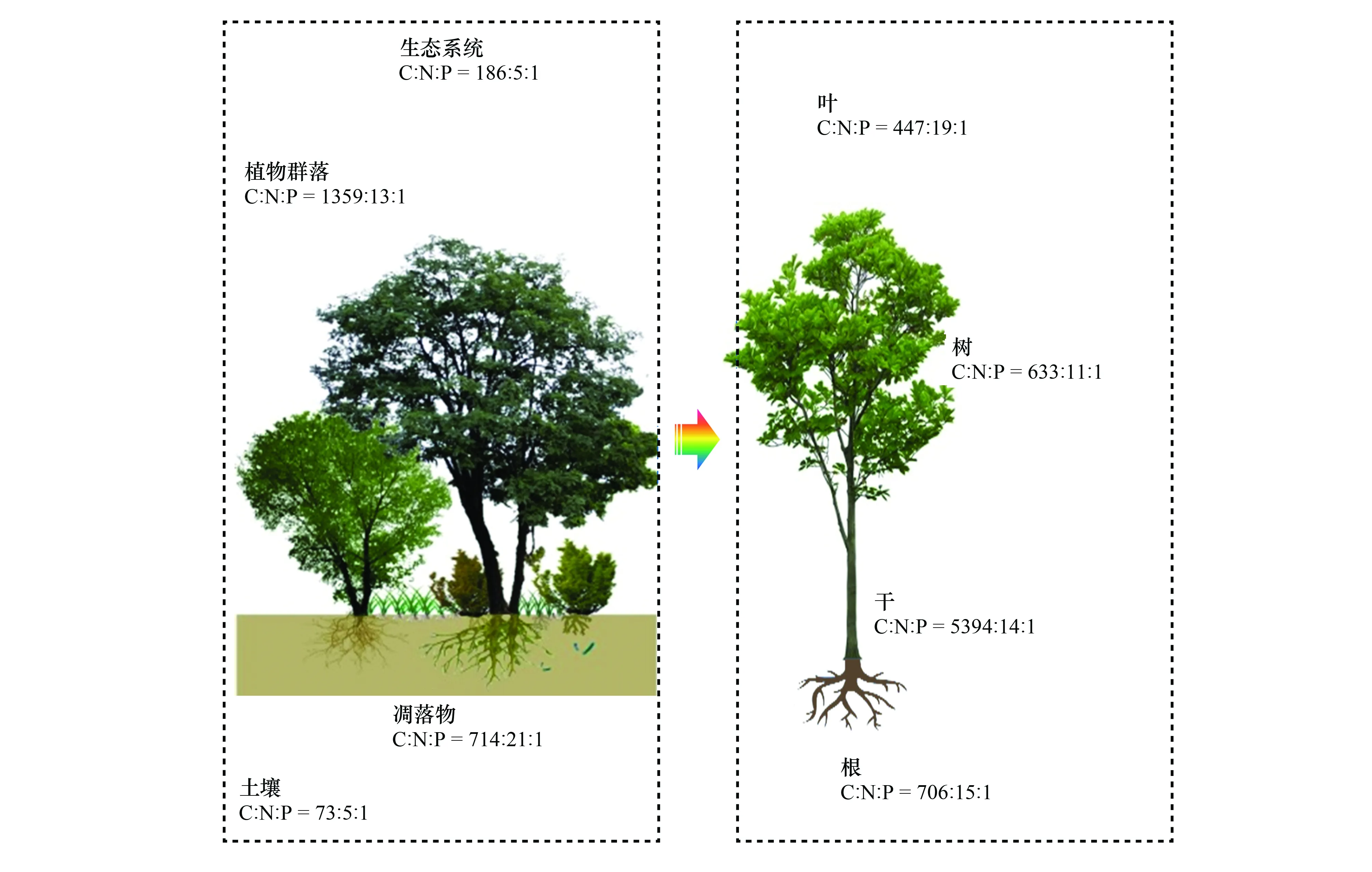
圖5 中國森林生態(tài)系統(tǒng)各組分及植物各器官C∶N∶P化學(xué)計(jì)量特征Fig.5 Variation of C∶N∶P stoichiometry in different plant organs and different ecosystem components in Chinese forests
相關(guān)研究填補(bǔ)了生態(tài)系統(tǒng)及不同組分、植物不同器官C∶N∶P化學(xué)計(jì)量學(xué)特征的實(shí)測數(shù)據(jù)的空白,并定量揭示了從熱帶雨林至寒溫帶針葉林的緯度變異規(guī)律與影響因素。群落尺度的元素化學(xué)計(jì)量特征及其緯度變異規(guī)律,可為全球C、N、P耦合模型提供基礎(chǔ)數(shù)據(jù)和理論支持。
5 土壤微生物和酶活性的緯度格局及其影響因素
5.1 土壤微生物群落的生物地理格局及其影響因子
傳統(tǒng)上,土壤微生物的地理分布被認(rèn)為是隨機(jī)的,可在條件允許的任何環(huán)境中擴(kuò)散和生長[79];同時(shí),土壤微生物的空間分布特征與大型動(dòng)植物存在明顯區(qū)別[80, 81]。近年來,隨著土壤微生物群落結(jié)構(gòu)的大尺度空間分布格局研究的增多,越來越多結(jié)果表明土壤微生物具有地帶性的分布規(guī)律,并將其歸因于生態(tài)系統(tǒng)地上與地下聯(lián)系、地理分隔和微生物的限制性擴(kuò)散[82]。
采用磷脂脂肪酸法測定了NSTEC樣帶上9個(gè)地帶性森林土壤微生物群落。實(shí)驗(yàn)結(jié)果表明:溫帶森林的土壤總磷脂脂肪酸(PLFA)要高于熱帶及亞熱帶。土壤細(xì)菌生物量的緯度格局與土壤總 PLFA的緯度格局一致。土壤真菌生物量無明顯的緯度變異。熱帶及亞熱帶土壤真菌/細(xì)菌(F/B)、革蘭氏陽性菌/革蘭氏陰性菌(G+/G-)值要高于北方溫帶及暖溫帶森林土壤。NSTEC具有明顯的水熱組合梯度,其氣候因子是土壤微生物生物量表現(xiàn)出緯度變異的重要原因。
在空間格局上,土壤微生物的群落結(jié)構(gòu)可能受各種生物與非生物因素的影響[83]。NSTEC數(shù)據(jù)也表明溫度、降水、土壤溫度對土壤微生物群落結(jié)構(gòu)變異的影響最大[84],在區(qū)域尺度,氣候可以通過影響土壤溫度及含水量來影響土壤微生物群落結(jié)構(gòu)[85]。每種土壤微生物都有最適溫度,當(dāng)環(huán)境溫度改變時(shí),高于或低于其最適溫度,微生物必定會(huì)被環(huán)境淘汰,土壤微生物群落結(jié)構(gòu)進(jìn)而發(fā)生改變。土壤水分增加將會(huì)導(dǎo)致某些細(xì)胞溶解,土壤含水量降低則導(dǎo)致土壤微生物胞質(zhì)皺縮或質(zhì)壁分離,從而改變土壤微生物群落結(jié)構(gòu)。與其他環(huán)境因子相比,土壤酸堿度(pH)及植物功能群特征對NSTEC土壤微生物群落結(jié)構(gòu)的緯度變異影響較小。植物功能群特征中,只有喬木比葉面積的群落權(quán)重值與土壤總PLFA、細(xì)菌、放線菌及G+呈顯著或極顯著負(fù)相關(guān),而其他植物性狀與土壤微生物生物量無顯著相關(guān)關(guān)系。
5.2 土壤微生物功能多樣性的生物地理格局及其影響因子
土壤微生物的代謝多樣性是分析微生物對環(huán)境適應(yīng)能力的有效指標(biāo)[86]。有關(guān)環(huán)境因素及人類活動(dòng)對土壤微生物的結(jié)構(gòu)、多樣性及酶活性的影響的研究已有很多[84, 87],并發(fā)現(xiàn)土壤微生物群落及酶活性具有生物地理分布格局。微生物的功能多樣性還會(huì)隨著植被類型及氣候條件的變化而變化[88];不同氣候區(qū)土壤微生物對碳源的利用能力不同,如北方森林土壤微生物更多的利用羧酸類碳源,而對氨基酸類的碳源的利用能力較弱[89]。
采用Biolog EcoPlate微平板培養(yǎng)法,測定了NSTEC樣帶的土壤微生物群落代謝活性。其中Biolog EcoPlate微孔板的總體顏色變化可用平均孔顏色變化率表示(Average well color development,AWCD),它可反映土壤微生物活性。實(shí)驗(yàn)結(jié)果表明:培養(yǎng)起始24 h內(nèi),AWCD變化不明顯,而培養(yǎng)24—96 h間 AWCD快速增長,此時(shí)微生物活性旺盛,隨后緩慢增長直至趨于穩(wěn)定。土壤微生物群落的碳源代謝強(qiáng)度變化趨勢與AWCD一致,呼中和東靈山森林土壤微生物群落的碳源代謝強(qiáng)度最低,分別為65.7和59.7。尖峰嶺森林土壤最高為157.6;而其他7個(gè)站點(diǎn)土壤微生物群落碳源代謝強(qiáng)度無顯著差異,變化范圍為130—140。整體而言,在NSTEC樣帶土壤微生物碳源代謝強(qiáng)度無顯著的緯度變異規(guī)律。
利用96 h的微生物吸光值對土壤微生物群落功能多樣性指數(shù)進(jìn)行了計(jì)算,結(jié)果顯示不同的多樣性指數(shù)均呈現(xiàn)相似的空間變化規(guī)律,從熱帶雨林到寒溫帶針葉林,土壤微生物群落功能多樣性具有空間變異性。土壤微生物對碳源的利用能力隨著緯度變化而變化,并可聚為4類,相同氣候帶森林土壤微生物的碳源代謝能力相似。通過對土壤微生物對6種碳源類型的利用程度的分析,發(fā)現(xiàn)土壤微生物對碳源的利用主要集中在糖類、羧酸、氨基酸和聚合物等4類物質(zhì),四者相對吸光值約為85%。土壤微生物對胺類及芳香類物質(zhì)的利用程度較低,其相對吸光值之和約為15%。
在所有影響因素中,植物功能群特征是影響土壤微生物功能多樣性緯度變異的主要因子。通常,含氮量高的植物生長速率更快,其凋落物也更有利于細(xì)菌生長[90],而含氮量低的植物凋落物則更利于真菌生長[84,90]。在NSTEC樣帶上,較高的葉片氮含量一定程度上限制土壤微生物的碳源利用能力[91]。
5.3 土壤細(xì)菌多樣性的緯度格局及其影響因子
土壤微生物多樣性的生物地理學(xué)研究,有助于深入挖掘土壤中的未知生物資源,深刻理解土壤中微生物多樣性的產(chǎn)生、維持機(jī)制,并預(yù)測陸地生態(tài)系統(tǒng)結(jié)構(gòu)和功能的演變方向[82, 92]。隨著分子生物學(xué)的快速發(fā)展,越來越多研究表明土壤微生物群落組成和多樣性在大尺度地理空間上呈現(xiàn)規(guī)律性分布[87, 93]。環(huán)境因素、歷史進(jìn)化因素(距離分隔、物理屏障、擴(kuò)散限制和環(huán)境異質(zhì)性等)或者二者共同作用是造成土壤微生物群落空間分布格局的主要原因[87, 93, 94]。
以NSTEC樣帶9個(gè)地帶性森林土壤為研究對象,借助高通量測序手段,分析了不同森林土壤細(xì)菌α多樣性(OTU 豐富度和Faith’s 譜系多樣性)和群落結(jié)構(gòu)的變化[95]。實(shí)驗(yàn)結(jié)果表明:亞熱帶和熱帶森林生態(tài)系統(tǒng)中細(xì)菌OTU 豐富度和Faith’s 譜系多樣性顯著低于溫帶森林生態(tài)系統(tǒng),而土壤有機(jī)質(zhì)的可利用性是引起細(xì)菌OTU 豐富度和Faith’s 譜系多樣性變異的主要因素。此外,不同森林土壤細(xì)菌群落結(jié)構(gòu)也存在顯著變化。環(huán)境指標(biāo)和地理距離可以解釋80.5%的不同森林土壤微生物群落結(jié)構(gòu)的空間變異;其中,氣候因子、土壤和植物因素分別貢獻(xiàn)了11.9%、14.2% 和7.8%。通過偏蒙特爾分析發(fā)現(xiàn),溫度、降水、土壤酸堿度和土壤有機(jī)質(zhì)的可利用性是引起細(xì)菌群落結(jié)構(gòu)變異的最主要的驅(qū)動(dòng)因子。雖然植物葉片碳氮比會(huì)一定程度影響到細(xì)菌豐富度,但植物功能性狀對土壤細(xì)菌多樣性的貢獻(xiàn),遠(yuǎn)不如氣候和土壤。
5.4 土壤酶活性的生物地理格局及其影響因子
土壤酶是土壤生物過程的主要調(diào)節(jié)者[96],與土壤有機(jī)質(zhì)分解、礦質(zhì)營養(yǎng)元素循環(huán)、能量轉(zhuǎn)移等密切相關(guān)[97],在生態(tài)系統(tǒng)生物地球化學(xué)循環(huán)中具有重要作用。開展大尺度土壤酶活性的生態(tài)化學(xué)計(jì)量關(guān)系及影響因素的研究,能夠?yàn)榻沂緟^(qū)域尺度土壤碳循環(huán)的生物調(diào)控機(jī)制提供依據(jù)[98]。
利用微孔板熒光法,測定了NSTEC樣帶9個(gè)地帶性森林土壤的酶活性,主要是與土壤碳循環(huán)相關(guān)的土壤β- 1,4-葡萄糖苷酶(BG)、與土壤氮循環(huán)相關(guān)的土壤β- 1,4-N-乙酰葡糖氨糖苷酶(NAG)和L-亮氨酸氨基肽(LAP)以及與土壤磷循環(huán)相關(guān)的酸性磷酸酶(AP)。實(shí)驗(yàn)結(jié)果表明:不同氣候帶的森林土壤BG、NAG、LAP、AP酶活性具有較大差異。在樣帶尺度,BG 和NAG 酶活性從寒溫帶森林到暖溫帶森林逐漸升高,隨后又逐漸降低。總體上,北方森林土壤BG和NAG酶活性高于南方森林土壤。相反,土壤AP酶活性則表現(xiàn)為南方森林高于北方森林。根據(jù)熱帶雨林土壤較低的BG∶AP、NAG∶AP酶活性比值(0.21、0.13),可推斷磷是該地區(qū)養(yǎng)分物質(zhì)循環(huán)過程的主要限制元素。熱帶、亞熱帶森林土壤AP酶活性較高,而BG∶AP和(NAG+LAP)∶AP酶活性比值較低,表明亞熱帶及熱帶森林土壤磷有效性較低,生態(tài)系統(tǒng)更受磷元素的限制。對全球土壤AP酶活性的整合分析發(fā)現(xiàn),隨著緯度降低、氣候逐漸趨于濕熱,土壤AP酶活性逐漸升高,而BG∶AP、(NAG+LAP)∶AP酶活性比值逐漸減小[99]。
森林土壤BG、NAG酶活性的空間分布與年均降水量格局呈顯著負(fù)相關(guān),并隨著年平均溫度的升高而顯著降低,但沒有發(fā)現(xiàn)土壤LAP和AP酶活性空間分布與年均降水量、年均溫顯著相關(guān),年均溫可能是影響全球尺度土壤酶活性的一個(gè)關(guān)鍵因子。年均降水量通過影響土壤含水量來影響底物的擴(kuò)散速率[100],從而影響土壤酶活性。在NSTEC樣帶上,土壤BG、NAG酶活性與土壤酸堿度呈顯著正相關(guān),而土壤AP酶活性與土壤酸堿度呈負(fù)相關(guān)[90, 101]。
6 土壤有機(jī)質(zhì)的緯度格局與影響因素
6.1 土壤有機(jī)質(zhì)組分的緯度格局與影響因素
土壤有機(jī)質(zhì)十分復(fù)雜,根據(jù)其周轉(zhuǎn)時(shí)間可分為活性有機(jī)碳和穩(wěn)定有機(jī)碳[102]。土壤活性有機(jī)碳易被土壤微生物分解利用,且易氧化分解并對環(huán)境和人為干擾的響應(yīng)更敏感[103, 104];土壤活性碳組分空間格局及其影響因素的分析,有助于闡明土壤有機(jī)碳(SOC)在人為干擾或氣候變化情景下的早期變化和軌跡[105]。
以NSTEC樣帶9個(gè)典型森林土壤為研究對象,分析測試了其表層SOC、易氧化有機(jī)碳(EOC)、土壤微生物碳(MBC)和可溶性有機(jī)碳(DOC)含量[106]。實(shí)驗(yàn)結(jié)果表明:SOC含量隨緯度增加呈增加趨勢,在涼水最大,尖峰嶺最小。土壤EOC含量隨緯度增加也呈增長趨勢,9個(gè)森林站點(diǎn)EOC含量差異顯著并在涼水最大。MBC隨緯度增加整體呈增加趨勢,在呼中最大而東靈山最小;不同地區(qū)間差異很大,最大值與最小值間可達(dá)7.85倍。DOC含量的緯度變化格局與其他碳組分完全相反,其含量隨緯度增加呈降低趨勢。從氣候區(qū)來看,SOC和EOC整體表現(xiàn)為熱帶森林<亞熱帶森林<溫帶森林;DOC則表現(xiàn)為熱帶森林>亞熱帶森林>溫帶森林。
基于結(jié)構(gòu)方程的分析,氣候、植被生物量、土壤質(zhì)地和土壤微生物分別解釋了SOC、EOC、MBC和DOC緯度變異的74%、65%、51%和76%。其中,SOC和EOC緯度格局主要受氣候和土壤質(zhì)地的影響,植被生物量和土壤微生物的影響相對較弱。MBC緯度格局的主要控制因素是氣候,土壤質(zhì)地對其影響十分微弱。對于DOC緯度格局,氣候和植被生物量表現(xiàn)為正效應(yīng),土壤質(zhì)地則表現(xiàn)為負(fù)效應(yīng)。
6.2 土壤腐殖碳組分緯度格局及其影響因素
土壤腐殖質(zhì)是土壤穩(wěn)定性碳庫的主要來源,長期而言土壤腐殖質(zhì)的損耗將造成大量二氧化碳從土壤向大氣輸入[107];同時(shí),土壤腐殖質(zhì)的化學(xué)抵制力是控制土壤和大氣間碳循環(huán)的重要過程[108]。因此,大尺度的土壤腐殖碳組分及其格局形成機(jī)制,將為碳動(dòng)力模型的發(fā)展及驗(yàn)證提供數(shù)據(jù)支撐。
以NSTEC樣帶9個(gè)地帶性森林土壤為研究對象,分析測試了其表層土壤可提取腐殖質(zhì)碳(HEC)、胡敏酸碳(HAC)、富里酸碳(FAC)和胡敏素碳(HUC)含量。實(shí)驗(yàn)結(jié)果表明:森林土壤腐殖碳組分HAC、FAC、HUC和HEC含量范圍分別為:0.44—1.62 g kg-1、7.80 —18.30 g kg-1、11.37—49.98 g kg-1和8.53—19.94 g kg-1;它們在9個(gè)森林類型間都存在顯著性差異。整體而言,森林土壤腐殖碳組分HAC、FAC、HUC和HEC均隨緯度的增加呈增長趨勢。按氣候帶類型劃分,森林土壤腐殖質(zhì)碳組分都表現(xiàn)為熱帶<亞熱帶<溫帶的特點(diǎn),HAC和HEC在熱帶、亞熱帶和溫帶之間都差異顯著[109]。
氣候、土壤質(zhì)地和土壤微生物對森林土壤腐殖質(zhì)碳組分HAC、FAC、HUC和HEC的解釋率分別為91.32%、90.94%、94.22%和94.13%。氣候因素對HAC、FAC和HEC的影響最大,分別為78.72%、88.61%和89.8%;土壤質(zhì)地對HUC空間格局的影響最大,可解釋其85.92%空間變異。
6.3 土壤和植被碳貯量空間分布格局
科學(xué)家圍繞森林碳貯量的空間格局及其控制因素開展了大量的研究,但絕大多數(shù)區(qū)域尺度的研究都是單獨(dú)探討植被碳貯量和土壤碳貯量的空間格局及其影響因素[110- 113],而同時(shí)考慮森林植被和土壤碳貯量的空間格局的研究卻鮮有報(bào)道,限制了人們對整個(gè)森林生態(tài)系統(tǒng)碳貯量的空間分布格局和控制機(jī)制的認(rèn)識(shí)。
以中國東部森林為研究對象,收集和整理了2004—2014年間中國東部森林公開發(fā)表科研論文中的植被(地上生物量+根生物量)和土壤碳貯量數(shù)據(jù),共包括1471個(gè)樣地的植被數(shù)據(jù)和1999個(gè)樣地的土壤數(shù)據(jù)。在隨機(jī)取樣下,植被和土壤碳貯量存在緯度格局;如果僅考慮成熟林,則植被和土壤碳貯量表現(xiàn)出更強(qiáng)的緯度格局。此外,隨著統(tǒng)計(jì)尺度的增大(緯度2°和5°),森林植被和土壤碳貯量的緯度分布格局越發(fā)明顯[25]。在2°緯度下統(tǒng)計(jì),成熟林植被碳貯量從南向北由196.31 Mg C/hm2降低到51.71 Mg C/hm2,并隨緯度增加呈顯著的負(fù)指數(shù)關(guān)系;然而,0—10 cm土壤碳貯量隨著緯度的增加卻呈指數(shù)增加,由南向北土壤平均碳貯量從35.29 Mg C/hm2增加到123.28 Mg C/hm2;出人意料,森林生態(tài)系統(tǒng)(植被+土壤)的碳貯量沒有明顯緯度格局。植被和土壤碳貯量的分配比例(植被:土壤)隨緯度增加呈顯著的負(fù)對數(shù)關(guān)系,該比例系數(shù)能一定程度地為森林碳密度估算模型的建立提供科學(xué)指導(dǎo)。
氣候因素不僅決定著森林植被和土壤碳貯量的緯度分布格局,而且還控制著森林植被和土壤碳貯量分配比例的變化規(guī)律;然而,氣候?qū)χ脖惶假A量和土壤碳貯量的控制機(jī)制完全相反,對中國東部植被碳貯量而言降雨量更重要,而對土壤碳貯量而言溫度更重要[25]。隨著緯度的增加,地表凋落物呈逐漸增加的變化趨勢[114]。利用多源數(shù)據(jù)(收集文獻(xiàn)數(shù)據(jù)和中國科學(xué)院碳專項(xiàng)實(shí)測數(shù)據(jù)),何念鵬等還發(fā)展了基于經(jīng)典Logstic 生長方程的中國森林植被碳固持模型(Forest Carbon Sequestration Model, FCS model),并成功地對2010—2050植被固碳潛力及其空間格局進(jìn)行了預(yù)測[115]。相關(guān)研究結(jié)論揭示了全球氣候變化下森林的固碳效應(yīng),同時(shí)建議在理論研究或模型優(yōu)化時(shí)應(yīng)將植被和土壤分開考慮。
7 土壤有機(jī)質(zhì)周轉(zhuǎn)速率及其溫度敏感性
7.1 土壤碳礦化溫度敏感性的空間變異及其影響因素
土壤有機(jī)質(zhì)分解所釋放的二氧化碳是陸地生態(tài)系統(tǒng)與大氣間最大的氣體交換通量之一[116],土壤有機(jī)質(zhì)分解對溫度變化非常敏感;因此準(zhǔn)確地評估溫度變化對土壤有機(jī)質(zhì)分解的影響,對預(yù)測土壤碳庫變化及其對全球氣候變化的反饋具有重要意義。溫度敏感性(Q10)被用來表示土壤有機(jī)質(zhì)分解對溫度變化的響應(yīng),它為不同生態(tài)系統(tǒng)類型或研究間提供了一個(gè)規(guī)范的和可比較的參數(shù),同時(shí)也是許多碳循環(huán)模型的重要參數(shù)[117]。然而,由于各個(gè)獨(dú)立研究中傳統(tǒng)培養(yǎng)模式與測試方法的差異(且集成在單點(diǎn)或單個(gè)區(qū)域),導(dǎo)致不同研究結(jié)果間可比性較差,使許多科學(xué)家通常在模型中將Q10設(shè)定為常數(shù)(2.0或1.5),是模型預(yù)測不確定性的重要來源[118, 119]。如何革新土壤有機(jī)質(zhì)分解溫度敏感性的研究方法,對于降低不同區(qū)域研究間的不確定性、準(zhǔn)確評估土壤呼吸對氣候變化的響應(yīng)及其反饋均具有重要意義。
2013年以來,何念鵬等自主研發(fā)了土壤有機(jī)質(zhì)溫度敏感性的測定系統(tǒng)[120, 121],發(fā)展了“自動(dòng)變溫培養(yǎng)+連續(xù)自動(dòng)測試”新模式。新模式除了較好地克服了傳統(tǒng)模式(恒溫培養(yǎng)+間斷測試)存在的主要問題(微生物適應(yīng)性、底物消耗不均、操作過程繁瑣、樣品測試量大),還被成功應(yīng)用于研究不同生態(tài)系統(tǒng)類型或情景下土壤有機(jī)質(zhì)溫度敏感性[41, 122- 125]。基于NSTEC樣帶的研究,發(fā)現(xiàn)土壤有機(jī)質(zhì)基礎(chǔ)分解速率(20 ℃)在不同森林類型間差異顯著,且隨著緯度的升高而顯著升高。而且土壤有機(jī)質(zhì)分解的溫度敏感性(Q10)在不同森林類型間差異顯著,隨著緯度的升高顯著升高,為低溫區(qū)更敏感假設(shè)提供了直接的證據(jù)[41]。在此基礎(chǔ)上,對中國東部森林土壤以及內(nèi)蒙和西藏草地土壤溫度敏感性進(jìn)行了測定,發(fā)現(xiàn)Q10變化范圍從1.16到3.19(均值1.63),且在不同生態(tài)系統(tǒng)間差異顯著:高寒草地(2.01)> 溫帶草地(1.81)> 熱帶森林(1.59)> 溫帶森林(1.55)> 亞熱帶森林(1.52)[42]。
在未來氣候變暖的背景下,需要特別關(guān)注高緯度森林和高寒草地生態(tài)系統(tǒng)。在不同的生態(tài)系統(tǒng)中,Q10與底物質(zhì)量之間顯著的負(fù)相關(guān)關(guān)系均支持碳-質(zhì)量溫度假說,表明該假說也適用于較大空間尺度的陸地生態(tài)系統(tǒng)。環(huán)境因子和土壤因子共解釋了52%的Q10空間變異;其中,土壤pH值和電導(dǎo)率是最主要因素。在中國東部森林,這兩個(gè)因素又主要受到氣候因素所影響,因此,未來應(yīng)當(dāng)考慮將這兩個(gè)因素加入模型,更準(zhǔn)確的模擬及預(yù)測有機(jī)質(zhì)分解對氣候變化的響應(yīng)與反饋。
7.2 土壤氮礦化速率及其溫度敏感性的空間格局與影響因素
7.2.1 土壤總氮礦化速率的空間格局與影響因素
氮素是植物生長的重要營養(yǎng)元素,土壤氮素有效性一定程度上決定了陸地生態(tài)系統(tǒng)的凈初級生產(chǎn)力大小[126]。土壤總銨化速率和總硝化速率是土壤供氮能力的重要指標(biāo),硝化作用指由微生物將有機(jī)氮或銨態(tài)氮轉(zhuǎn)化為硝態(tài)氮的過程,是植物直接吸收利用的僅次于銨態(tài)氮的另一種無機(jī)氮形式[40, 127]。 盡管一些研究在單站點(diǎn)尺度上分析了土壤的銨化或者硝化作用,但是在較大的區(qū)域尺度上,森林土壤總銨化和總硝化作用對于不同的環(huán)境因素變化的響應(yīng)及其機(jī)制仍不清楚。
采用15N庫稀釋法進(jìn)行野外原位標(biāo)記和室內(nèi)標(biāo)記的培養(yǎng)實(shí)驗(yàn),探討了從熱帶到寒溫帶9種配對森林土壤(原生林vs.次生林)的潛在銨化和硝化速率[40]。實(shí)驗(yàn)結(jié)果表明:野外平均銨化速率是(4.9 ± 0.5) mg N kg-1d-1,而平均潛在銨化速率是(32.0 ± 8.6) mg N kg-1d-1。野外平均硝化速率為(1.7 ± 0.3) mg N kg-1d-1,潛在硝化速率為(3.2 ± 0.6) mg N kg-1d-1。 在樣帶尺度上,銨化速率顯著高于硝化速率,并且在不同森林中銨化速率和硝化速率具有較大的變異性。銨化速率和硝化速率與氣象因素(溫度和降水)之間存在顯著的相關(guān)關(guān)系。結(jié)構(gòu)方程模型表明:土壤氮含量,微生物氮庫以及細(xì)菌的豐度是影響中國東部森林土壤銨化和硝化速率的主要因素。土壤因素對銨化和硝化速率較強(qiáng)的控制,意味著只有準(zhǔn)確地定量土壤因素及其對土壤氮轉(zhuǎn)化過程影響,才能更好地改進(jìn)土壤氮模型。
7.2.2 土壤凈氮礦化及其溫度敏感性的空間格局及其影響因素
在全球變暖的背景下,土壤凈氮礦化速率對溫度變化響應(yīng)的微小變化,都會(huì)改變土壤的氮素有效性,并一定程度上影響陸地生態(tài)系統(tǒng)的初級生產(chǎn)力[128]。凈氮礦化作用對溫度變化的響應(yīng)也可用溫度敏感性(Q10)表示,這一參數(shù)被廣泛用在土壤N循環(huán)模型中[129]。過去關(guān)于土壤凈氮礦化溫度敏感性的研究主要集中在單點(diǎn)上,而在大尺度上土壤凈氮礦化及其溫度敏感性的研究極少[130]。
基于1990—2014年間公開發(fā)表論文的土壤凈氮礦化及其溫度數(shù)據(jù),探討了凈氮礦化及其溫度敏感性的空間格局及調(diào)控機(jī)制[131]。結(jié)果顯示:在中國陸地生態(tài)系統(tǒng)中,土壤凈氮礦化速率的均值為(2.78 ± 0.18) mg N kg-1d-1,它隨著緯度的升高顯著降低。另外,土壤凈氮礦化速率隨著海拔高度的增加呈顯著的對數(shù)降低趨勢。在不同森林植被類型中,常綠針葉林和常綠闊葉林土壤凈氮礦化速率高于落葉針葉林和落葉闊葉林。土壤凈氮礦化的Q10均值為1.58,并在不同的生態(tài)系統(tǒng)間差異顯著。在森林生態(tài)系統(tǒng)中,常綠針葉林的Q10顯著高于落葉闊葉林和常綠闊葉林。土壤凈氮礦化的Q10隨著緯度和海拔的增加均呈顯著增加的趨勢,支持越寒冷的區(qū)域其溫度敏感性越高的理論假設(shè)。通徑分析結(jié)果顯示:土壤凈氮礦化速率主要受到年均溫和土壤有機(jī)質(zhì)含量的影響,而Q10主要受到土壤總氮含量的影響。此外,關(guān)于全球土壤氮礦化的整合研究也表明,土壤凈氮礦化的Q10存在明顯的緯度格局且在生態(tài)系統(tǒng)類型間差異顯著[130]。這些結(jié)果表明在氣候變暖的背景下,寒冷區(qū)域土壤的氮素有效性可能會(huì)增加,進(jìn)而促進(jìn)生態(tài)系統(tǒng)初級生產(chǎn)力的增加。
8 挑戰(zhàn)與展望
通過對近年來NSTEC典型森林樣帶性狀研究的簡要總結(jié),可以清楚地看出在如下幾個(gè)方面取得較好的進(jìn)展:①生態(tài)系統(tǒng)性狀(植物、微生物、土壤)的系統(tǒng)化測試,構(gòu)建了一個(gè)系統(tǒng)的、獨(dú)特的性狀數(shù)據(jù)庫,并探討基于典型森林生態(tài)系統(tǒng)性狀研究的新調(diào)查模式,為更好地基于性狀數(shù)據(jù)探討生態(tài)系統(tǒng)結(jié)構(gòu)與功能及其對環(huán)境適應(yīng)奠定了基礎(chǔ)。該模式的提出和成功的應(yīng)用,將會(huì)一定程度上引領(lǐng)未來性狀研究的調(diào)查方式,并促進(jìn)系統(tǒng)性調(diào)查數(shù)據(jù)的快速積累。②基于所測定的大量數(shù)據(jù),不僅按傳統(tǒng)途徑探討了植物、微生物和土壤性狀的緯度變異規(guī)律與影響因素,其中如氣孔性狀、解剖結(jié)構(gòu)性狀、葉綠素含量、非結(jié)構(gòu)性碳水化合物含量、土壤碳礦化、土壤凈氮礦化、溫度敏感性、生態(tài)系統(tǒng)各組分C∶N∶P化學(xué)計(jì)量特征等的大尺度分析,填補(bǔ)了相關(guān)研究的空白。更為重要的是以系統(tǒng)性的調(diào)查數(shù)據(jù)為基礎(chǔ),發(fā)展了基于植物群落結(jié)構(gòu)和生物量(葉片面積)加權(quán)法的從器官至群落的拓展方法,實(shí)現(xiàn)了單位土地面積(或單位器官)基礎(chǔ)的群落性狀推導(dǎo),較好地解決了國內(nèi)外性狀研究所面臨的巨大挑戰(zhàn)和難題。③定量揭示了天然森林生態(tài)系統(tǒng)中(部分)性狀與初級生產(chǎn)力、凈初級生產(chǎn)力、水分利用效率等的定量關(guān)系,為今后進(jìn)一步探討天然森林群落性狀與功能的關(guān)系提供了新的方法學(xué)依據(jù)和范例。
由于新調(diào)查模式所獲得的系統(tǒng)性數(shù)據(jù),更接近自然群落或生態(tài)系統(tǒng),因此,除了現(xiàn)有的分析角度或思路,還應(yīng)該具有更多的潛在用途。首先,從傳統(tǒng)生態(tài)學(xué)角度可以深入探討如下幾方面的科學(xué)問題:①綜合分析葉片或根系性狀的相互關(guān)系,甚至考慮分級化的“葉片性狀網(wǎng)絡(luò)”“根系性狀網(wǎng)絡(luò)”,再從點(diǎn)-樣帶-區(qū)域多層次探討性狀網(wǎng)絡(luò)變異及其機(jī)制。如上所述,簡單地從幾個(gè)指標(biāo)探討的關(guān)系有其科學(xué)價(jià)值和進(jìn)步意義,但還難以真實(shí)地揭示自然群落中植物性狀間的關(guān)系,其研究類似于“瞎子摸象”,難以揭示其整體關(guān)系與影響機(jī)制;類似的思路還適用于土壤微生物,是否可以建立 “微生物性狀網(wǎng)絡(luò)”。②利用系統(tǒng)化的調(diào)查數(shù)據(jù)(植物群落結(jié)構(gòu)-土壤微生物群落結(jié)構(gòu)-土壤理化性質(zhì)-氣候因子、群落內(nèi)每個(gè)植物物種葉-枝-干-根性狀、葉片形態(tài)-氣孔-解剖結(jié)構(gòu)-化學(xué)元素-葉綠素含量-非結(jié)構(gòu)性碳水化合物等),深入探討森林生態(tài)系統(tǒng)群落結(jié)構(gòu)在點(diǎn)-樣帶-區(qū)域尺度的維持機(jī)制,探討經(jīng)典生態(tài)學(xué)的核心科學(xué)問題“進(jìn)化vs.適應(yīng)、環(huán)境過濾效應(yīng)vs.競爭效應(yīng)、生態(tài)位分化理論vs.中性理論”等,推動(dòng)生態(tài)學(xué)理論研究的發(fā)展;③ 結(jié)合點(diǎn)-樣帶-區(qū)域尺度的生態(tài)系統(tǒng)生產(chǎn)力參數(shù),從葉片性狀協(xié)同優(yōu)化(或權(quán)衡)角度,深入探討森林生態(tài)系統(tǒng)生產(chǎn)力優(yōu)化機(jī)制。
同時(shí),能否為傳統(tǒng)性狀研究切實(shí)服務(wù)于宏觀生態(tài)學(xué)或宏觀地理學(xué)提供技術(shù)探討的平臺(tái)?如果能實(shí)現(xiàn)該目標(biāo),既可極大拓展性狀研究的范疇,又能使傳統(tǒng)性發(fā)展數(shù)據(jù)更好地服務(wù)于當(dāng)前區(qū)域甚至全球生態(tài)環(huán)境問題。然而,傳統(tǒng)性狀數(shù)據(jù)與遙感、模型和能量數(shù)據(jù)間存在巨大的裂隙,使它們之間很難相互驗(yàn)證或支持;具體表現(xiàn)為:①傳統(tǒng)性狀數(shù)據(jù)大多是在個(gè)體或群落類少數(shù)物種測試,缺乏相配套的群落結(jié)構(gòu)數(shù)據(jù),使其數(shù)據(jù)很難推導(dǎo)到群落尺度;然而,宏觀生態(tài)學(xué)的主要觀測尺度都是群落尺度;②傳統(tǒng)性狀數(shù)據(jù)的測試指標(biāo)的量綱更多是根據(jù)自身要求,集中在器官水平的含量或密度,而宏觀生態(tài)學(xué)的量綱大多在單位面積或體積,因此二者間如何轉(zhuǎn)化也是一個(gè)非常棘手的難題;③傳統(tǒng)的性狀與功能的關(guān)系,大多是在器官或物種水平建立的(如葉片氮含量與光合速率的關(guān)系、葉片葉綠素含量與光合速率的關(guān)系等),其是否適用于復(fù)雜的群落還有待商榷。上述三個(gè)難點(diǎn)是相互關(guān)聯(lián)的,只有通過系統(tǒng)性的性狀調(diào)查數(shù)據(jù)才能逐一解決。此外,為了更好地構(gòu)建性狀研究(植物、動(dòng)物、微生物等)與宏觀生態(tài)學(xué)研究的橋梁,需要從更高層次提出新的概念,如生態(tài)系統(tǒng)性狀(Ecosystem trait)。在此,將生態(tài)系統(tǒng)性狀定義為以單位土地面積為基礎(chǔ),在群落尺度能體現(xiàn)生物(植物、動(dòng)物、微生物)對環(huán)境適應(yīng)性和生產(chǎn)力優(yōu)化的性狀;即生態(tài)系統(tǒng)性狀是由植物群落性狀、動(dòng)物群落性狀、土壤微生物群落性狀等組成。它可以在相對統(tǒng)一的空間尺度和量綱上將植物群落性狀、動(dòng)物群落性狀、土壤微生物群落性狀、土壤屬性和氣候要素等緊密聯(lián)系起來,為深入研究不同尺度生物-非生物性狀間的關(guān)系奠定堅(jiān)實(shí)基礎(chǔ),并以群落性狀為核心連接宏觀生態(tài)學(xué)的主要研究途徑(遙感觀測、通量觀測和模型模擬),推動(dòng)傳統(tǒng)性狀指標(biāo)與宏觀生態(tài)學(xué)的應(yīng)用相結(jié)合。
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
