β-木糖苷酶的研究進展
2018-11-03 02:49:12湯勇,蔡俊*
中國釀造 2018年10期
湯 勇,蔡 俊*
(湖北工業大學 發酵工程教育部重點實驗室 工業發酵湖北省協同創新中心 工業微生物湖北省重點實驗室,湖北 武漢 430068)
植物細胞壁的橫向結構主要由半纖維素和木質素結合而成。β-1,4-木聚糖是木質纖維素生物質中含量最多的半纖維素組分。這種復雜的多糖的基本骨架由β-1,4糖苷鍵相連的D-吡喃木糖組成,并且含有不同的側鏈取代基,最常見的取代基為乙酰基、阿拉伯糖和葡萄糖醛酸。側鏈的數量和組成取決于木聚糖來源,例如硬木木聚糖的側鏈包括葡萄糖醛酸基和乙酰基,如圖1所示,而軟木木聚糖的側鏈包括葡萄糖醛酸基和阿拉伯呋喃糖基,如圖2所示。由于植物木聚糖聚合度和取代側鏈的不同,完全降解需要一套木聚糖水解酶系,酶系中各種酶的作用位點見圖3,主要包括β-1,4-木聚糖酶和β-木糖苷酶(β-xylosidase,β-xyl)[1]。其中內切β-1,4-木聚糖酶破壞木聚糖骨架成更短的可溶性低聚木糖;β-木糖苷酶水解可溶性低聚木糖和木二糖的非還原末端釋放木糖。該催化作用還消除了內切木聚糖酶的終產物抑制作用[2]。
1 β-木糖苷酶的來源
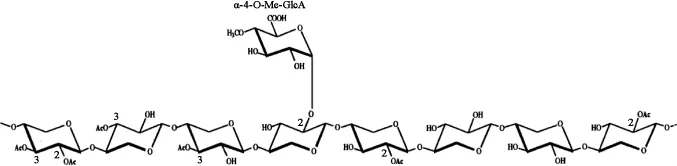
圖1 硬木木聚糖的結構Fig.1 Structure of the hardwood xylan
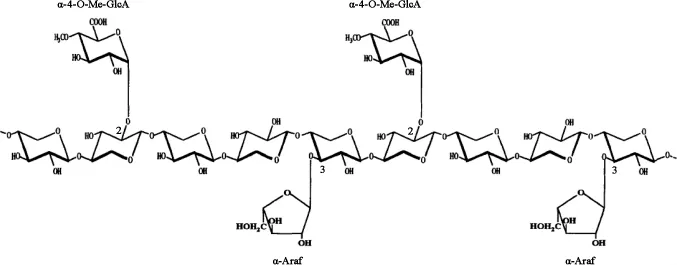
圖2 軟木木聚糖的結構Fig.2 Structure of the softwood xylan
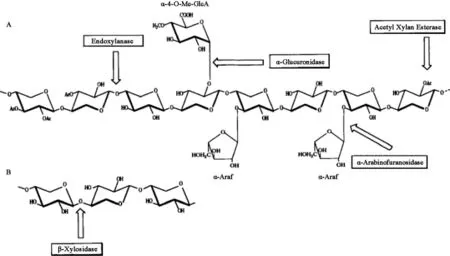
圖3 木聚糖降解酶系參與水解木聚糖(A)與β-木糖苷酶水解低聚木糖(B)Fig.3 The xylanolytic enzymes involved in the degradation of xylan(A)and hydrolysis of xylooligosaccharide by β-xylosidase(B)
β-木糖苷酶在自然界中廣泛存在,常見于細菌和真菌之中,真菌中又多為絲狀真菌,而極少見于酵母菌中。MHETRAS N等[3]篩選得到一株產β-木糖苷酶酵母菌Pseudozyma hubeiensis NCIM 3574。雖然自然界中存在著大量產β-木糖苷酶的微生物,但其所產β-木糖苷酶的酶活和酶學特性不夠理想,所以近十幾年來,人們致力于通過基因工程的手段來解決上述問題。例如SHI H等[4]成功將Thermotoga thermarum DSM 5069中編碼表達β-木糖苷酶的基因導入大腸桿菌(Escherichia coli)BL21中表達。該重組表達的β-木糖苷酶具有更高的活性、耐熱性和木糖耐受性。
2 β-木糖苷酶的分類
由于β-木糖苷酶對不同底物的活性是不同的,所以將這些酶根據底物特異性來分類是很困難的。迄今為止,根據碳水化合物活性酶數據庫(CAZy),β-木糖苷酶分為糖苷水解酶家族(glycoside hydrolases,GHs)3,30,39,43,52和54,如表1所示[5]。該家族的分類取決于酶所使用的底物、氨基酸序列相似性、三維結構和作用機制。已知絲狀真菌的木糖苷酶只屬于第3、第43和第54家族,如表1中第3家族中細菌木糖苷酶占89%,第43家族中細菌木糖苷酶占92%。
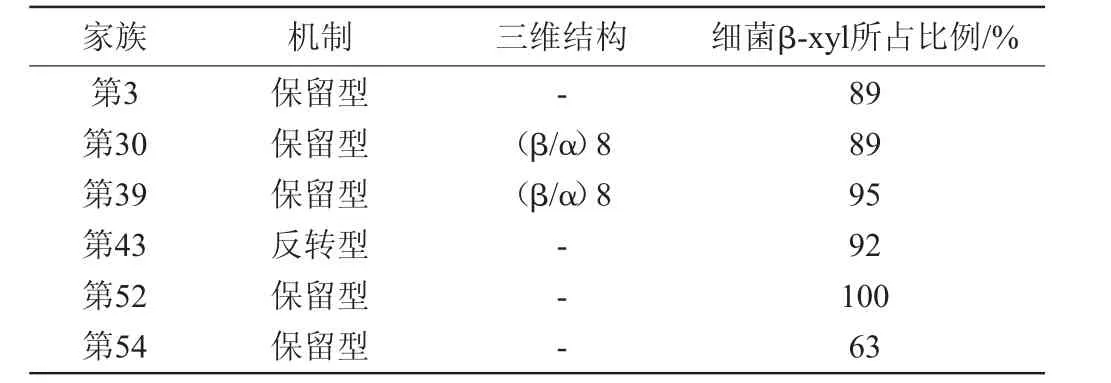
表1 CAZY數據庫的β-木糖苷酶家族分類Table 1 β-xylosidase family classification according to the CAZY database
3 β-木糖苷酶的分泌部位
與胞外分泌的木聚糖酶相反,不同微生物和不同培養條件下,β-木糖苷酶可以是胞外型或膜結合型。如絕大多數細菌和酵母所產β-木糖苷酶被認為是以可溶形式分泌于細胞質中。在淺白隱球酵母(Cryptococcusalbidus)中,木二糖和木三糖通過β-木糖苷通透酶系統進入細胞,然后由β-木糖苷酶轉化為木糖[6]。在這些微生物中,低聚木糖的水解必須在細胞內完成。相反,真菌所產β-木糖苷酶可分為胞內型(如Streptowycestherwoviolaceus OPC-520)、胞外型(如Aureoba sidiumpullulans)和膜結合型(如Streptomyces sp.EC 10)[7]。某些絲狀真菌β-木糖苷酶早期與菌絲體結合,然后通過分泌或細胞裂解釋放到胞外。大多數純化后的真菌β-木糖苷酶分子質量>100 kDa,少數分子質量較低。從這些微生物純化得到的β-木糖苷酶主要以單體和二聚體的形式存在。
4 β-木糖苷酶的特性
不同微生物來源的β-木糖苷酶的最適pH和最適溫度存在差異見表2。
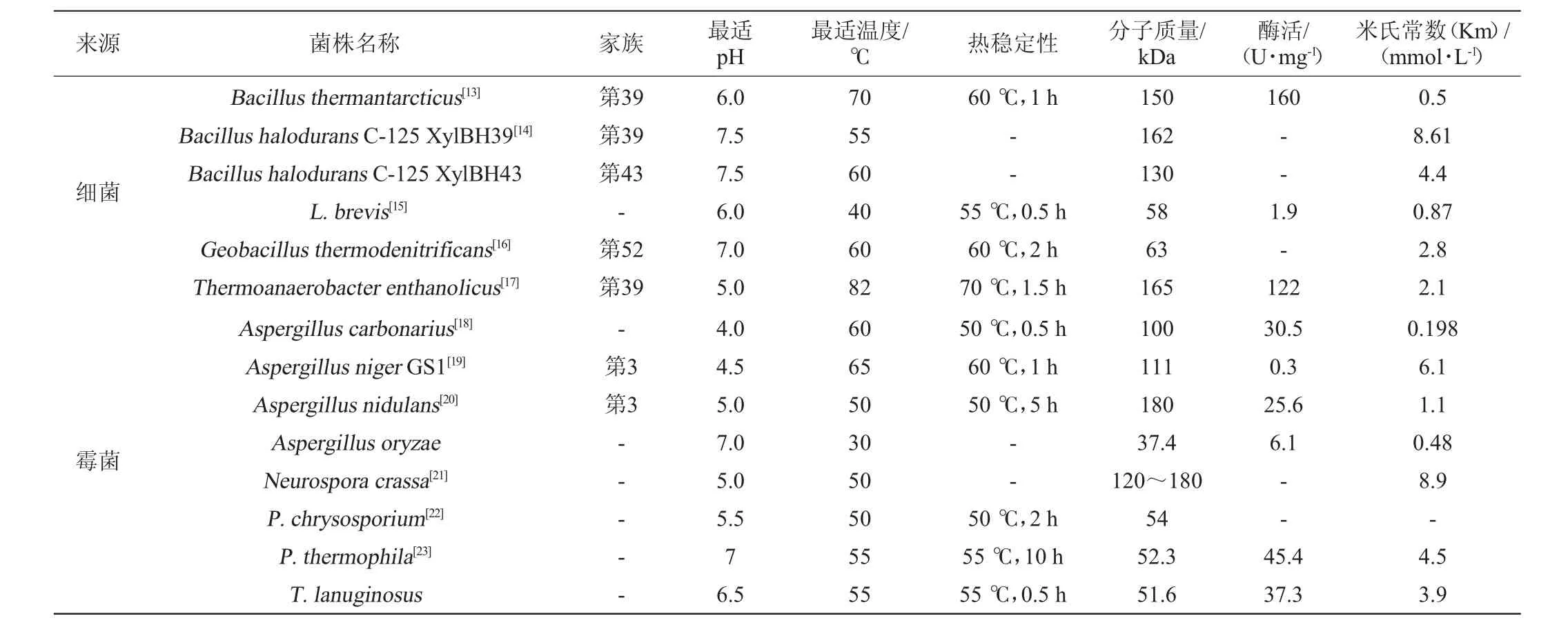
表2 不同微生物來源的β-木糖苷酶的物理化學性質Table 2 Physicochemical properties of β-xylosidase from different microorganism
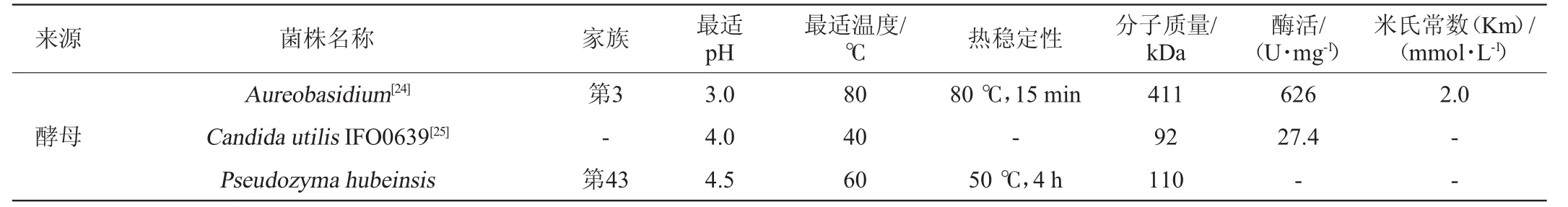
續表
由表2可知,大多β-木糖苷酶的最適pH在3.0~6.0。一般情況下,細菌所產β-木糖苷酶最適pH高于真菌β-木糖苷酶。當然,也有在更酸或中性pH下具有最佳活性的β-木糖苷酶。例如菌核青霉(Penicillium sclerotiorum)[8]所產β-木糖苷酶的最適pH為2.5;米曲霉(Aspergillus oryzae)所產β-木糖苷酶的最適pH為7.0[9];疏綿狀嗜熱絲孢菌(T.lanuginosus)[10]所產β-木糖苷酶的最適pH為6.5。
β-木糖苷酶所要求的反應溫度一般比較溫和,在40~60℃之間,而近年來人們越來越關注木糖苷酶的熱穩定性。褐色高溫單孢菌(Thermomonosporafusca)所產β-木糖苷酶在65℃的條件下保存8h以上仍有一定的活性,在70℃的條件下可保存1.5h[11]。糞堆梭菌(Clostridium stercorarium)所產β-木糖苷酶最適反應溫度高達80℃,并且在50~70℃的環境下仍然具有良好的穩定性[12]。由于缺乏對木糖苷酶三維立體結構的確定,導致絲狀真菌木糖苷酶熱穩定性的結構因素至今還不得而知。木糖苷酶結構中存在的二硫鍵、疏水相互作用、電子對之間的相互作用可能是影響其熱穩定性的原因。此外,在一些情況下,將硫醇和二價金屬離子通過固定化或者化學修飾加到β-木糖苷酶上可以增強其熱穩定性,從而防止溫度的升高所導致的催化活性的降低,同時降低微生物污染的風險。耐熱菌株的這一特性為酶的貯存和工業化利用提供了很大的便利。
真菌β-木糖苷酶的一個典型特征是其轉木糖基化活性。轉木糖基活性(transxylosylation activity)是通過形成新的糖苷鍵進而形成更長的低聚糖的一種能力(轉糖基:將糖基供體上活化的糖基或單糖轉移到糖基接受體的過程)[26]。由于其高選擇性和立體定向性,這種轉糖基作用也被用作酶法合成β-木聚糖酶底物和各類糖苷。SULISTYOJ等[27]報道了粉狀曲霉(A.pulverulentus)所產具有轉木糖基活性的β-木糖苷酶將低聚木糖的木糖殘基轉移到各種受體如醇和酚類化合物的過程。例如來自泡盛曲霉(A.awamori)X-100的β-木糖苷酶因其顯著的轉木糖基能力可以產生聚合度>7的低聚木糖[28]。
5 β-木糖苷酶的催化機制
半纖維素酶多為糖苷水解酶,一種廣泛的水解兩種或多種碳水化合物之間或碳水化合物和非碳水化合物部分之間的糖苷鍵的酶系[29]。在GHs分類中的酶,例如β-木糖苷酶,通過兩種不同的機制裂解糖苷鍵來起作用:保留端基異位碳構型(保留型)或反轉異位碳構型(反轉型)[30]。糖苷水解酶第3家族和第54家族的成員采用保留型,第43家族則含有反轉型糖苷水解酶。
目前人們已經比較清楚的是β-木糖苷酶的水解過程為酸堿催化反應,活性位點處的兩個關鍵催化氨基酸(如谷氨酸或天冬氨酸)一個作為一般酸(質子供體),另一個作為親核試劑[31]。其中,保留型酶采用雙取代反應機制,包括糖基化反應和轉糖基化反應;反轉型則采用單取代機制[32]。青霉菌(Penicillium herquei)和炭色孢腔菌(Cochliobolus carbonum)所產β-木糖苷酶是由絲狀真菌所產使用保留型機制的β-木糖苷酶。谷氨酸和天冬氨酸被普遍認為是所有絲狀真菌β-木糖苷的催化殘基[33]。
6 β-木糖苷酶的誘導和阻遏
木聚糖降解酶系中最重要的兩種酶:木聚糖酶和β-木糖苷酶均為典型的誘導酶。大多數研究發現木聚糖水解酶的表達受到木聚糖或木糖的特異性誘導和CreA介導的碳分解代謝物阻遏。即碳源調節xlnR基因的轉錄,CreA介導阻遏xlnR基因的轉錄。因此,木聚糖降解酶基因的轉錄受到xlnR因子誘導和CreA介導的碳分解代謝物阻遏的共同影響。
木糖可以是木聚糖降解酶基因表達的誘導劑,有時也會抑制木聚糖降解酶基因的表達[34]。木糖觸發CreA介導的碳分解代謝物阻遏取決于環境中木糖的含量。例如在含有1%燕麥木聚糖和0.1%木糖的培養基中誘導康氏木霉(Trichoderma koningii)中β-木糖苷酶基因的表達。關于T.koningii的研究表明,提供少量的木糖可以促進β-木糖苷酶的誘導表達[35]。相反地,D-木糖則會阻遏T.emersonni和T.reesei中β-木糖苷酶基因的表達。對于特定的微生物,木糖苷酶的表達由不同的誘導劑誘導。例如,誘導黑曲霉(A.niger)木聚糖降解酶系的碳源是木聚糖,木二糖和D-木糖[36]。埃默森籃狀菌(T.emersonii)的木糖苷酶基因(bxl1)可以由較低濃度的木聚糖——甲基-β-D-吡喃木糖苷和木糖誘導[37]。
微生物的碳分解代謝物阻遏其實是一種節能機制,一旦環境存在著大量葡萄糖或者木糖,木聚糖降解酶就不必要合成。如在葡萄糖存在下,A.niger的β-木糖苷酶基因(xlnD)的表達被抑制[38]。此外,RAJOKA M I[39]發現,除誘導外,當微生物在生長于易于代謝的底物和無機氮源時還存在另外一種β-木糖苷酶合成調節機制。
7 β-木糖甘酶的分離純化的研究現狀
原核生物所產β-木糖苷酶是胞內酶,真菌所產β-木糖苷酶可分為胞內型、胞外型和膜結合型。通過對發酵上清液進行一系列分離純化可以得到純的β-木糖苷酶。其分離純化方法多為硫酸銨沉淀、超濾濃縮、離子交換層析、凝膠過濾層析等,不同微生物來源的β-木糖苷酶的常見分離純化步驟如表3。除上述常規方法,PAN I等[40]開發出了一種使用雙水相萃取對T.koningii所產β-木糖苷酶同時進行濃縮和純化的新方法。該方法快速、高效、回收率高且目標蛋白的純度與柱層析的純度相當。

表3 不同微生物來源的β-木糖苷酶的常見分離純化步驟Table 3 Common isolation and purification steps of β-xylosidase from different microorganisms
8 β-木糖苷酶的應用
β-木糖苷酶在工農業殘渣加工成燃料、化學和制藥、醫療用品、食品和飲料、纖維飼料、造紙工業等方面都是非常重要的[41]。β-木糖苷酶的主要應用如下:
(1)自然界中大量存在的木聚糖,使得其生物轉化后的產物低聚木糖和木糖有可能成為未來的能源。這種多糖轉化為單糖可以通過兩種方法進行:酸水解和酶水解。然而,酸水解過程會形成污染環境的副產物且成本高昂。因此,可再生型的生物轉化具有重要意義,即先通過酶水解木聚糖得到木糖,再由如酵母和休哈塔假絲酵母(Candida shehatae)等微生物通過發酵將木糖轉化為乙醇[42]。
(2)木寡糖是由木糖單元組成的低聚糖,具有不同的聚合度,被廣泛用作天然食品甜味劑,具有良好的降血糖和血脂的作用。木聚糖酶和木糖苷酶復合酶是生產含有2~4個殘基的木寡糖所必需的。另一方面,木寡糖作為益生元,可以選擇性地促進益生菌的生長,從而緩解便秘,促進消化和營養吸收等。
(3)由于傳統原料成本的增加,飼料中酶的添加量在過去幾年中大大增加。木聚糖降解酶能夠水解小麥,玉米和其他谷物中存在的半纖維素,促進營養物質的消化率,并減少氮磷的排放。單胃動物不產生植物細胞壁降解酶,如木聚糖和纖維素降解酶,導致谷物食品的利用率降低。這些酶的添加促進了能量代謝,降低了食物黏度,提高能量和蛋白質利用率,從而增加動物體質量[43]。
(4)木聚糖水解酶復合物也用于紙漿和造紙工業中,主要用于在增白之前加工纖維素紙漿。用木聚糖酶復合物進行預處理可以降低化學品,特別是氯和氧化氯的消耗。據報道,紙漿漂白中的酶的運用可以減少30%氯化合物使用量,減少環境中15%~20%的高毒性有機氯化合物的釋放[44]。
(5)常用的釀造技術中的添加劑如乳化劑,氧化劑可以被酶所替代。在啤酒釀造中,長鏈阿拉伯木聚糖被釋放,增加了酒體黏度和啤酒濁度。木聚糖酶和β-木糖苷酶的加入可以促進阿拉伯木聚糖溶解,通過將其降解為較小的寡糖,降低其黏度,消除啤酒濁度[45]。
9 結論與展望
隨著人們對木聚糖降解酶系的深入研究,近年來有關β-木糖苷酶的生物化學和分子生物學上的研究取得了重大進展,但迄今為止進行的大多數研究集中在β-木糖苷酶的篩選、純化、表征和重組表達。
首先,關于產β-木糖苷酶微生物的篩選,還沒有直接簡便的篩選方法。已知產β-木糖苷酶微生物多集中在細菌和霉菌,關于產β-木糖苷酶的酵母菌和具有完整木聚糖降解酶系統的酵母的研究還很欠缺。另一方面,植物性和動物性來源的β-木糖苷酶還有待進一步開發和研究。
其次,由于β-木糖苷酶在工業上的廣泛應用使得其必須具有更高的產量、更高的活性和更好的生化特性,如熱穩定性、pH穩定性等。為了提高β-木糖苷酶的產量和活性,比較常用的手段為誘變育種、基因工程育種、酶的固定化和調節酶的誘導和阻遏效應。其中誘變育種原理簡單,但獲得有益突變株的占比不多;相對基因工程育種,誘變育種方法的操作步驟更少。沒有高通量篩選方法的條件下誘變育種的工作量較大,要處理大量突變株。然而基因工程育種目的性強,是目前用的較多、效果較好的一種手段,同時基因工程育種可以有效的改良β-木糖苷酶的生化特性,已經有過很多關于通過β-木糖苷酶基因的重組表達大大提高了其熱穩定性,使其更高的應用于工業生產的報道。
由于缺乏三維立體結構測定的研究,β-木糖苷酶熱穩定性和其結構之間的關系至今還不清楚。目前已知β-木糖苷酶的糖基化及其熱穩定性,此外,將硫醇和二價金屬離子通過固定化或者化學修飾加到β-木糖苷酶上可以提高其熱穩定性。所以,β-木糖苷酶結構是未來值得研究的一個方向。
由于β-木糖苷酶是誘導酶又受到分解代謝物反饋抑制,目前比較清楚的是在一些情況下,少量木糖和乙醇可以促進β-木糖苷酶的表達,但關于低分子質量化合物誘導木聚糖水解酶系表達的分子機制還知之甚少;木聚糖和高分子量底物是β-木糖苷酶有效的誘導劑,一般情況下,β-木糖苷酶表達會受到木糖和葡萄糖引起的碳分解代謝物阻遏。所以,如何提高β-木糖苷酶木糖/葡萄糖耐受性或解除其阻遏效應或者探索β-木糖苷酶酶合成調節機制是亟待解決的問題。
最后,由于木聚糖的異質性,需要木聚糖水解酶系統:β-1,4-內切木聚糖酶、β-木糖苷酶、α-L-阿拉伯呋喃糖苷酶、α-D-葡萄糖醛酸酶、α-D-半乳糖苷酶、乙酰木聚糖酯酶、對香豆酸酯酶和阿魏酸酯酶這一系列酶的協同作用將其降解至木糖。目前研究較多的是β-1,4-內切木聚糖酶和β-木糖苷酶間的協同作用,關于其他酶之間的協同作用和實用性研究還很欠缺。然而,已有報道,某些微生物所產β-木糖苷酶還同時具有α-L-阿拉伯呋喃糖苷酶活性,對于含有兩種或多種酶活性的β-木糖苷酶的來源和原理仍然值得進一步探究。
