代謝工程改造釀酒酵母生產(chǎn)2,3-丁二醇的研究進展
2018-11-05 00:49:56劉德安王長麗葛菁萍
中國釀造 2018年9期
劉德安,王長麗,丁 昊,葛菁萍,2*
(1.黑龍江大學(xué)生命科學(xué)學(xué)院 微生物學(xué)黑龍江省高校重點實驗室,黑龍江 哈爾濱 150080;2.農(nóng)業(yè)微生物技術(shù)教育工程研究中心,黑龍江 哈爾濱 150500)
隨著當(dāng)今社會經(jīng)濟的蓬勃發(fā)展,能源短缺逐漸成為人類發(fā)展進程的絆腳石,所以開發(fā)綠色新能源,減緩石油危機迫在眉睫[1]。基于釀酒酵母(Saccharomyces cerevisiae)全基因組測序的完成,利用代謝工程手段對S.cerevisiae進行遺傳學(xué)改造,為實現(xiàn)生物燃料如乙醇、2,3-丁二醇(2,3-butanediol,2,3-BD)的工業(yè)化生產(chǎn)提供了保障[2]。此方法的興起不但能在保護生態(tài)環(huán)境的同時發(fā)展經(jīng)濟,實現(xiàn)能源結(jié)構(gòu)的綠色轉(zhuǎn)型,更是響應(yīng)“十一五”發(fā)展規(guī)劃、推動節(jié)能減排號召的一種有效手段。
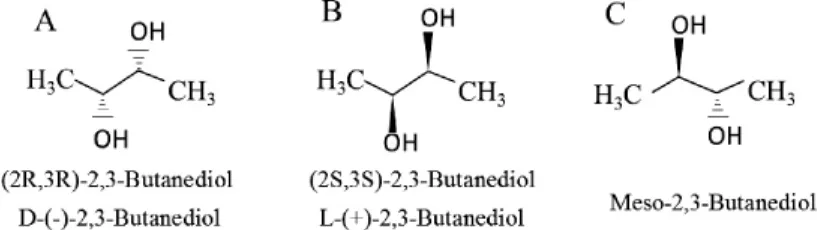
繼乙醇之后,2,3-BD是一種性價比較高的新興能源化合物,廣泛應(yīng)用于航空航天、食品、醫(yī)藥等領(lǐng)域。2,3-BD脫羧化反應(yīng)合成乙偶姻(acetoin)和雙乙酰(2,3-butanedione),可以作為食品風(fēng)味添加劑等[3]。20世紀(jì)中期,以石油為原料化學(xué)法合成2,3-BD一直占據(jù)主導(dǎo)地位。直到20世紀(jì)后期,特別是21世紀(jì)以來,石油危機、糧食短缺和環(huán)境污染等問題的日益加劇,使得生物法合成2,3-BD又重新吸引了國內(nèi)外學(xué)者的目光,對其的研究再次成為了關(guān)注的熱點[4]。2,3-BD的結(jié)構(gòu)式如下:
自然界存在許多天然合成2,3-BD的微生物(見表1),如肺炎克雷伯氏菌(Klebsiella pneumonia)[5]、產(chǎn)酸克雷伯氏菌(Klebsiellaoxytoca)[6]、產(chǎn)氣腸桿菌(Enterobacter aerogenes)[7]、多粘類芽孢桿菌(Paenibacillus polymyxa)[8]、S.cerevisiae[9],這些微生物能應(yīng)用代謝工程策略提高2,3-BD的生產(chǎn)性能。因此,有研究利用敲除副產(chǎn)物合成途徑的關(guān)鍵酶編碼基因的手段來提高2,3-BD的產(chǎn)量[10-12]。SANGPROO M等[13]在K.oxytoca細(xì)胞內(nèi)敲除副產(chǎn)物合成途徑中的乙醛脫氫酶(acetaldehyde dehydrogenase,ALDA)、乙醇脫氫酶(alcohol dehydrogenase,ADH),最終使工程菌株的2,3-BD產(chǎn)量達到114 g/L。利用代謝工程手段雖然能有效地提高細(xì)菌2,3-BD的產(chǎn)量,但諸如克雷伯氏菌屬(Klebsiella sp.)和腸桿菌屬(Enterobacter sp.)等細(xì)菌大都是條件致病菌,違反了綠色安全工業(yè)化生產(chǎn)的標(biāo)準(zhǔn)[13]。因此,利用公認(rèn)安全無毒(generally regarded assafe,GRAS)微生物S.cerevisiae改造合成2,3-BD的代謝途徑成為新的研究熱點[14]。
近代以來隨著代謝工程原理與技術(shù)的不斷深入研究,關(guān)于S.cerevisiae產(chǎn)2,3-BD代謝途徑改造的深入探索,改造方法和手段的不斷創(chuàng)新,2,3-BD的生產(chǎn)得到了大幅度增加。本研究總結(jié)了近年來利用代謝工程原理和方法對S.cerevisiae代謝網(wǎng)絡(luò)進行重新設(shè)計和合理改造的三種策略。并展望未來S.cerevisiae的重點研究方向。

表1 產(chǎn)2,3-BD的微生物Table1 2,3-butanediol-producing microorganism
1 微生物中2,3-BD的代謝途徑
在細(xì)菌的2,3-BD代謝途徑中(見圖1A),糖類物質(zhì)經(jīng)糖酵解途徑(embden meyerhof pathway,EMP)變?yōu)楸?丙酮酸經(jīng)α-乙酰乳酸合成酶(α-acetolactatesynthase,ALS)、α-乙酰乳酸脫羧酶(α-acetolactatedecarboxylase,ALDC)和乙偶姻還原酶(acetoin reductase,AR)或稱2,3-BD脫氫酶(2,3-butanediol dehydrogenase,BDH)3個關(guān)鍵酶共同催化后合成α-乙酰乳酸、乙偶姻和2,3-BD[19]。這一系列反應(yīng)是酸性發(fā)酵過程,所以會伴隨著甲酸、丁二酸、乙酸、蘋果酸等酸性副產(chǎn)物的生成。
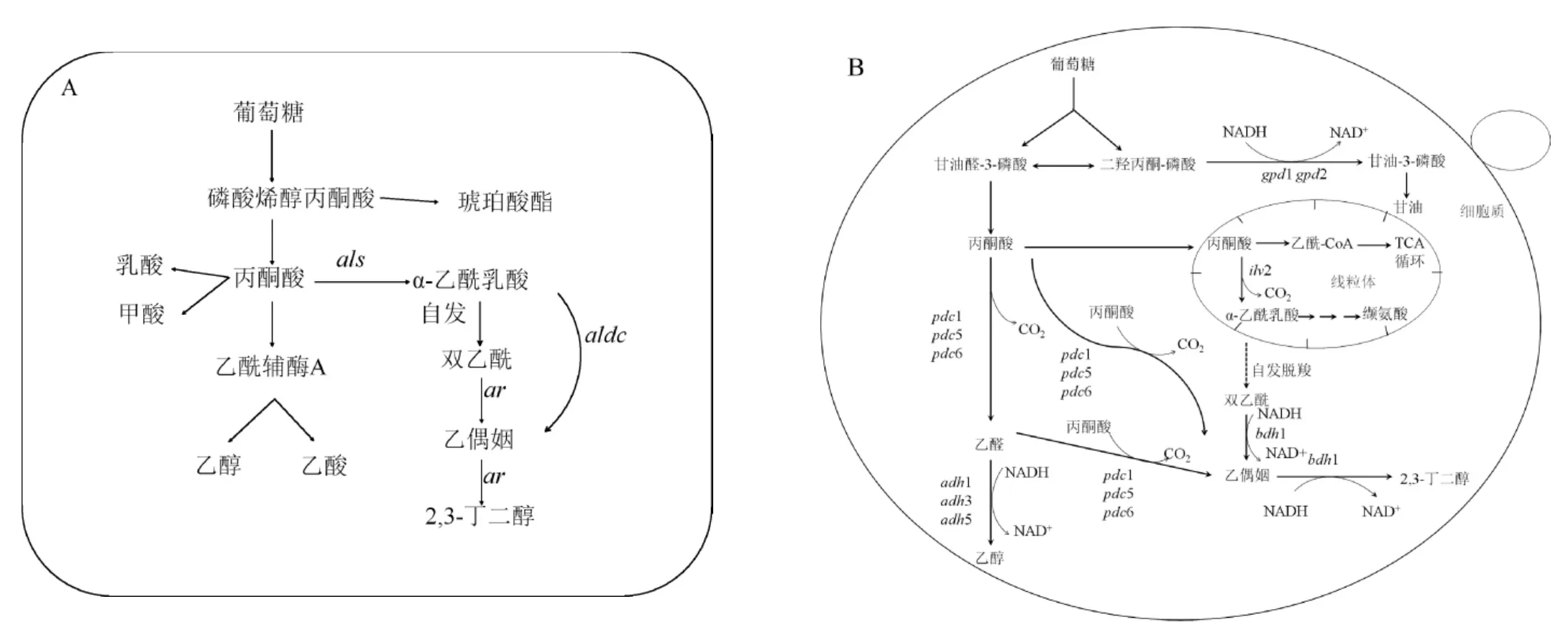
圖1 細(xì)菌(A)和釀酒酵母(B)中的2,3-BD代謝途徑Fig.1 Metabolic pathway of 2,3-butanediol in bacteria(A)and Saccharomyces cerevisiae(B)
S.cerevisiae自身的2,3-BD代謝途徑(見圖1B)不完整,S.cerevisiae胞內(nèi)有兩條2,3-BD合成途徑,第一條途徑是丙酮酸被α-乙酰乳酸合成酶催化生成為α-乙酰乳酸,α-乙酰乳酸通過自身氧化脫羧生成雙乙酰,在BDH1的催化下生成乙偶姻;第二條途徑是丙酮酸經(jīng)丙酮酸脫羧酶(pyruvatedecarboxylase,PDC)催化生成乙醛,乙醛在乙醇脫氫酶(alcohol dehydrogenase,ADH)的作用下生成乙醇,或者丙酮酸在PDC的作用下直接生成乙偶姻;最后,乙偶姻在BDH1的催化作用下生成終產(chǎn)物2,3-BD。與細(xì)菌合成2,3-BD的合成途徑相比,S.cerevisiae中沒有ALDC,不能通過酶促反應(yīng)合成乙偶姻,但是在氧氣存在的條件下,S.cerevisiae中α-乙酰乳酸自發(fā)脫羧生成雙乙酰,在BDH1的催化作用下將雙乙酰還原為2,3-BD。
2 釀酒酵母合成2,3-BD的代謝途徑改造策略
之前,國內(nèi)外學(xué)者一直利用傳統(tǒng)的誘變育種技術(shù)(如物理誘變、化學(xué)誘變)來提高菌種的產(chǎn)量和性能,但這種方式具有隨機性強、可塑性弱且工作量大等弊端。因此,基于全基因組的系統(tǒng)代謝工程和輔酶調(diào)控策略對關(guān)鍵代謝節(jié)點進行合理的遺傳改造逐漸吸引了學(xué)者眼球。通過優(yōu)化S.cerevisiae合成2,3-BD的途徑,阻斷乙醇、甘油兩條主要的競爭代謝支路進而強化生物合成2,3-BD,有望快速獲得遺傳穩(wěn)定性好、發(fā)酵水平顯著提高的S.cerevisiae。
2.1 過表達關(guān)鍵酶基因
GONZáLEZE等[20]研究發(fā)現(xiàn),在酵母細(xì)胞代謝過程中丙酮酸脫羧酶可以將乙醛和丙酮酸轉(zhuǎn)化成乙偶姻,但碳代謝流向微弱,乃有痕量的2,3-BD產(chǎn)生。因此,有必要對2,3-BD生物合成途徑中關(guān)鍵酶進行過表達,并減少S.cerevisiae中分支代謝產(chǎn)物,用于提高2,3-BD的產(chǎn)量。
S.cerevisiae中存在兩條代謝通路,丙酮酸可以通過乙醛或α-乙酰乳酸兩條代謝路徑生成乙偶姻。因此,可以參考細(xì)菌2,3-BD的改造策略,如圖2A所不。LIANJ等[21]通過過表達丙酮酸甲酸裂解酶(pyruvate formate-lyase,PLFA)基因(pfl A,pfl B)、乙醛脫氫酶基因(mhp F)來增強乙醛的代謝途徑,同時敲除adh1、adh3和adh5基因抑制或減弱通往乙醇的代謝流,敲除自身2,3-BD生物合成的基因(ilv2),防止受自身代謝影響,引導(dǎo)代謝通路最終由產(chǎn)乙醇代謝轉(zhuǎn)向產(chǎn)2,3-BD,進而提高S.cerevisiae產(chǎn)2,3-BD的水平,通過分批補料發(fā)酵,2,3-BD的產(chǎn)量達到0.307 g/L,是野生型S.cerevisiae 2,3-BD產(chǎn)量的7.3倍,乙醇產(chǎn)量降低7.7%;BRACKMAN G等[22]通過異源過表達來自枯草芽孢桿菌(Bacillussubtilis)的乙酰脫氫酶基因E1組分的aco A、aco B來提高乙酰-CoA與乙醛合成乙偶姻的能力(見圖2B),在敲除adh1、adh3、adh5和ilv2基因的基礎(chǔ)上,過表達pfl A、pfl B、aco A、aco B及內(nèi)源性的bdh1基因,構(gòu)建工程菌株進行分批補料發(fā)酵,2,3-BD產(chǎn)量達到0.341 g/L。由此說明乙醛途徑基因的過表達對增強S.cerevisiae的2,3-BD產(chǎn)量無顯著效果。
因此,最近的研究集中在α-乙酰乳酸合成途徑上。由于α-乙酰乳酸的自發(fā)氧化脫羧是S.cerevisiae 2,3-BD合成途徑中的限速步驟。為解決這一問題,在S.cerevisiae中異源表達胞質(zhì)als和aldc基因,可使α-乙酰乳酸直接將α-乙酰乳酸轉(zhuǎn)化為乙偶姻,最終合成2,3-BD[23]。由圖2C可知,KIM SR等[24]在S.cerevisiae中異源表達B.subtilis中的als和aldc基因并且過表達bdh1基因,通過分批補料發(fā)酵,2,3-BD產(chǎn)量達到0.69 g/L,是野生型S.cerevisiae 2,3-BD產(chǎn)量的115倍。綜上所述,異源過表達α-乙酰乳酸合成途徑中的2,3-BD關(guān)鍵酶基因更能直接的提高2,3-BD產(chǎn)量。
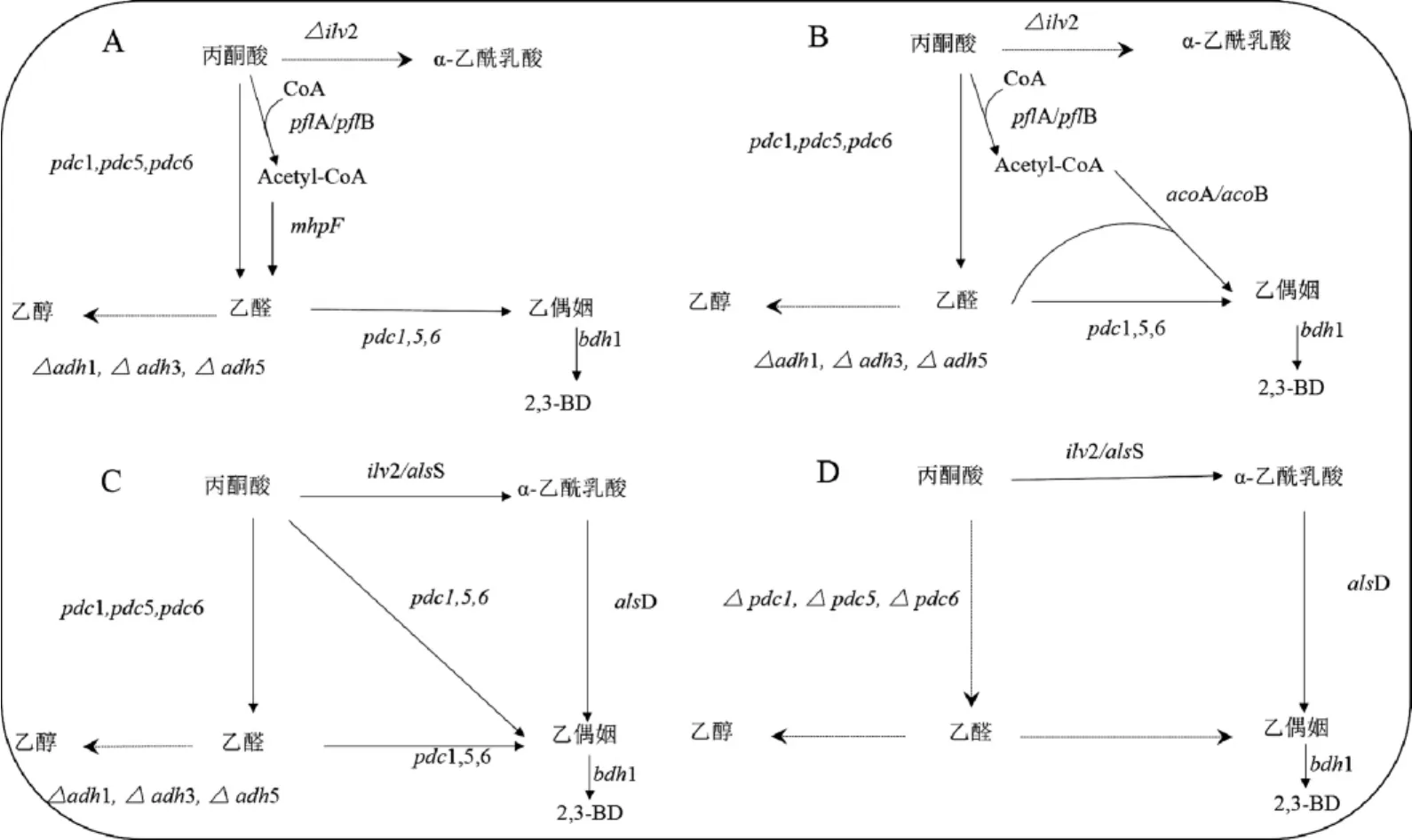
圖2 釀酒酵母中基因過表達與基因敲除策略Fig.2 Strategies of gene overexpression and gene knockout in Saccharomyces cerevisiae
2.2 敲除adh和pdc基因減少副產(chǎn)物形成
研究者發(fā)現(xiàn),雖然過表達各種2,3-BD生物合成途徑中的關(guān)鍵酶基因能提高2,3-BD的產(chǎn)量,但2,3-BD產(chǎn)量的提高程度并不理想[24]。大量乙醇的積累嚴(yán)重制約著代謝流向2,3-BD,因此,在這種情況下,阻斷或弱化乙醇的代謝流進而加大2,3-BD產(chǎn)量是至關(guān)重要的。由圖1B可知,S.cerevisiae代謝途徑中的PDC能將丙酮酸轉(zhuǎn)化為乙醛,并在ADH作用下進一步還原成乙醇。因此,采用敲除adh基因(圖2C)或pdc基因(圖2D)兩種代謝工程策略,將乙醇代謝通路阻斷是切實可行的。
LEEJ等[25]敲除S.cerevisiae中的adh1、adh3、adh5基因后進行分批發(fā)酵,2,3-BD的產(chǎn)量達到2.29 g/L,與野生型S.cerevisiae相比,乙醇的產(chǎn)量降低31.25%,而2,3-BD的葡萄糖轉(zhuǎn)化率顯著提高,為0.113 g/g;KIM S等[26]在Adh-S.cerevisiae菌株基礎(chǔ)上,異源過表達B.subtilis中的als S和als D基因和內(nèi)源bdh1基因后進行分批補料發(fā)酵,2,3-BD產(chǎn)量達到了29.1 g/L。理論上講,在抑制乙醇代謝流向并過表達2,3-BD合成途徑后,菌株可以利用葡萄糖經(jīng)丙酮酸、α-乙酰乳酸、乙偶姻合成2,3-BD,或經(jīng)丙酮酸和乙醛轉(zhuǎn)化成乙偶姻,最終合成2,3-BD(見圖2C)。但弊端是Δadh1、Δadh3、Δadh5會積累大量的毒性物質(zhì)乙醛,且還會生成一定量的乙醇[27]。
為了完全抑制乙醇的積累并高效地積累丙酮酸,科研工作者再次嘗試以Pdc-S.cerevisiae菌株作為出發(fā)菌株(見圖2D),從根本上解決乙醇大量積累的問題。由于Pdc-S.cerevisiae菌株不能合成乙醛,進而不能生成乙酰-CoA,而乙酰-CoA是三羧酸循環(huán)(tricarboxylic acid cyle,TCA)循環(huán)的關(guān)鍵物質(zhì),因此,菌株代謝生長受到嚴(yán)重抑制。在這種情況下,通過補充C2化合物(如乙醇、乙酸)可以緩解由乙酰-CoA缺失導(dǎo)致的細(xì)胞生長受抑制的情況,并顯著增加2,3-BD的產(chǎn)量。此外,通過表達mth1A轉(zhuǎn)錄調(diào)節(jié)因子也可以達到緩解細(xì)胞抑制現(xiàn)象,KIM SJ等[16]敲除Pdc-S.cerevisiae菌株中的pdc、ilv2基因和過表達細(xì)菌的2,3-BD基因(als S、als D、bdh1)和mth1A基因后,在微需氧條件下補料分批發(fā)酵,2,3-BD產(chǎn)量為96.2 g/L,并且沒有檢測到乙醇和丙酮酸積累。
2.3 減緩釀酒酵母合成2,3-BD過程中輔因子平衡問題
在酵母細(xì)胞的生物轉(zhuǎn)化過程中,常常包含了復(fù)雜的生化反應(yīng),且需要大量的輔因子參與調(diào)控,其中輔因子供給不足常常是影響代謝的關(guān)鍵因素。IDAY等[28]在Adh-S.cerevisiae或Pdc-S.cerevisiae菌株中引入2,3-BD代謝途徑后,會導(dǎo)致葡萄糖流向2,3-BD的代謝過程的煙酰胺腺嘌吟二核甘酸(reduced form of nicotinamide-adenine dinucleotid,NADH)/NAD+比例不平衡。原因是S.cerevisiae在糖酵解途徑中產(chǎn)生了2 mol NADH,但乃有1 mol NADH在2,3-BD途徑中被氧化成NAD+。一般來說,S.cerevisiae可以通過乙醇和甘油的合成途徑消耗掉多余的1mol NADH,從而維持胞內(nèi)輔酶NADH/NAD+的平衡[29]。但工程菌株Adh-S.cerevisiae或Pdc-S.cerevisiae阻斷了代謝過程中乙醇的合成支路。因此,若要維持輔酶NADH/NAD+平衡,細(xì)胞乃有通過大量積累甘油來實現(xiàn)胞內(nèi)NAD+的再生。為了保證2,3-BD合成的同時,最大程度的減少甘油的積累,將碳通量引流向2,3-BD合成支路是接下來需要考慮的問題。EHSANIM等[30]提出了增加供氧的策略,在敲除ilv2基因并過表達als、aldc和bdh1基因的S.cerevisiae中增加溶氧量,甘油積累減少的同時,葡萄糖合成2,3-BD的理論產(chǎn)量比厭O2條件下高20%。這說明通過改變?nèi)苎跽{(diào)節(jié)氧化還原平衡對于生產(chǎn)2,3-BD具有重要意義。
此外,科研工作者們還提出了另一種辦法,通過外源添加替代電子受體的策略來緩解甘油的積累。如KAWAIS等[23]利用2-丁酮(2-butanone)作為外源添加電子受體用以緩解過量的NADH,甘油的產(chǎn)量降低了56%,2,3-BD的產(chǎn)率提高了1.5倍,達到了2,3-BD理論產(chǎn)率的90%。出于此目的,KIM S等[15]發(fā)現(xiàn)NADH氧化酶(NADH oxidase,NOX)可以將NADH氧化成NAD+,為了降低NADH的濃度,在過表達als、aldc、bdh1基因的Adh-S.cerevisiae工程菌株中外源表達乳酸乳球菌(Lactococcus lactis)中的NADH氧化酶,從而將C代謝甘油流向2,3-BD。通過分批補料發(fā)酵,2,3-BD產(chǎn)量達到73 g/L,與出發(fā)菌株相比,甘油的產(chǎn)量降低了65.3%,2,3-BD的產(chǎn)量提高了23.8%。
2.4 利用廉價原料合成2,3-BD
在S.cerevisiae中除過表達2,3-BD合成途徑、敲除副產(chǎn)物途徑提高2,3-BD產(chǎn)量外,也可以通過代謝工程手段使工程菌株具備利用廉價原料(木質(zhì)纖維素(lignocellulose)和藻類(algae))生產(chǎn)2,3-BD的能力,進而提高2,3-BD產(chǎn)量。木質(zhì)纖維素水解成葡萄糖和木糖后,葡萄糖能被S.cerevisiae利用,但木糖不能被利用[31]。因此,為了使S.cerevisiae能夠利用木糖,首先要解決的問題是關(guān)于代謝過程中存在的糖抑制現(xiàn)象,即當(dāng)環(huán)境中存在葡萄糖時葡萄糖會抑制木糖的代謝。BAEY H等[32]發(fā)現(xiàn),在纖維素中存在一種兩個葡萄糖通過β-1,4-糖苷鍵組成的纖維二糖不會受其他糖類代謝抑制。為了利用纖維二糖產(chǎn)2,3-BD,NAN H等[17]在Pdc-S.cerevisiae菌株中外源導(dǎo)入纖維糊精轉(zhuǎn)運蛋白(cellodextrin transporter,CDT)基因和粗糙脈孢菌(Neurospora crassa)中的β-葡糖苷酶(β-glucosidase)基因(見圖3),通過補料分批發(fā)酵,工程菌株的2,3-BD產(chǎn)量達到5.29 g/L,葡萄糖轉(zhuǎn)化率為0.29 g/g,這表明了纖維素水解產(chǎn)物可以持續(xù)有效的進行2,3-BD的生產(chǎn)。其次要解決的問題是如何利用木糖生產(chǎn)2,3-BD,JO SE等[33]在過表達als、aldc和bdh1基因的Pdc-S.cerevisiae菌株中過表達木糖發(fā)酵酵母(Scheffersomyces stipitis)的木糖醇脫氫酶(xylitol dehydrogenase,XDH)基因和內(nèi)源性木酮糖激酶(xylulose kinase,XK)基因,構(gòu)成異源木糖同化途徑(見圖3),構(gòu)建完成的工程菌株進行補料分批發(fā)酵,2,3-BD產(chǎn)量達到44 g/L。綜上,S.cerevisiae可以通過代謝工程的技術(shù)利用木質(zhì)纖維素產(chǎn)2,3-BD。
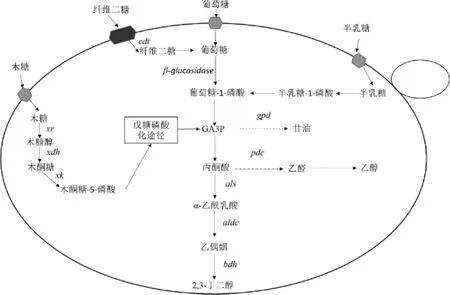
圖3 釀酒酵母中纖維二糖和藻類生物質(zhì)單糖的2,3-BD生物合成途徑Fig.3 2,3-BD biosynthetic pathway of cellobiose and algal biomass monosaccharides in Saccharomyces cerevisiae
藻類作為另外一種可再生的生物質(zhì)能源與纖維素生物質(zhì)相比具有許多優(yōu)點,如藻類的生產(chǎn)不會占用耕地,所需水資源少[34]。在海洋大型藻類中,紅藻中的天竺葵(Pelargonium hortorum Bailey)最為豐富,其水解產(chǎn)物由葡萄糖和半乳糖組成。為了能夠同時利用葡萄糖和半乳糖產(chǎn)2,3-BD,MITSUI K等[35]在過表達als、aldc和bdh1基因的Pdc-S.cerevisiae菌株中表達Mth1p基因后,菌株可以同時消耗葡萄糖和半乳糖,通過補料分批發(fā)酵,優(yōu)化葡萄糖與半乳糖的比例,菌株的2,3-BD產(chǎn)量達到100 g/L,葡萄糖轉(zhuǎn)化率為0.353 g/g。
3 總結(jié)與展望
21世紀(jì)以來,通過引用新型的代謝工程技術(shù),工程菌株S.cerevisiae產(chǎn)2,3-BD的能力顯著提升。此外,研究者建立了利用可再生能源來生產(chǎn)2,3-BD的方法[36]。S.cerevisiae利用可再生能源生產(chǎn)2,3-BD是有一定經(jīng)濟效益和具有可持續(xù)發(fā)展的前景[37]。展望未來,應(yīng)從以下幾個方面進行后續(xù)的研究。
(1)深入了解S.cerevisiae產(chǎn)2,3-丁二醇的代謝途徑和分支代謝途徑,利用基于全基因組的系統(tǒng)代謝工程和輔酶調(diào)控策略對關(guān)鍵代謝基因進行合理的遺傳改造,例如,人工合成基因線路技術(shù)、啟動子工程技術(shù)以及基因組編輯技術(shù)等優(yōu)化2,3-BD的代謝途徑。
(2)開發(fā)利用可再生能源產(chǎn)2,3-丁二醇,有些可再生能源木質(zhì)纖維素(如菊芋(Helianthustuberosus(L.1753)))水解液中的毒性物質(zhì)含量相對較少,并且水解液中含有豐富的營養(yǎng)物質(zhì),這有利于S.cerevisiae產(chǎn)2,3-丁二醇,通過優(yōu)化木質(zhì)纖維素的種類與水解方式以期得到更加適合S.cerevisiae生長和產(chǎn)2,3-BD的培養(yǎng)條件。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農(nóng)業(yè)(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(zhì)(2021年1期)2021-02-22 14:14:44
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年10期)2019-11-25 07:33:58
中國果業(yè)信息(2019年10期)2019-11-13 01:21:34
中學(xué)生數(shù)理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
聚氯乙烯(2018年9期)2018-02-18 01:11:34
中國衛(wèi)生(2016年3期)2016-11-12 13:23:26
