小麥脫水素 TaDHN-2基因功能與結構分析
2018-12-06 01:59:38段寧振李夢園王瀑童常亞南袁曉波樊亞棟李永春孟凡榮
麥類作物學報 2018年11期
段寧振,張 寧,李夢園,王瀑童,常亞南,袁曉波,樊亞棟,李永春,孟凡榮
(1.河南農業大學生命科學學院,河南鄭州 450002; 2.河南農業大學農學院,河南鄭州450002)
小麥是我國最主要的糧食作物之一,其產量多少直接影響我國的糧食安全和社會穩定[1-2]。干旱、高鹽等逆境脅迫是影響作物高產的主要限制因素。植物受到逆境脅迫誘導后會產生一系列調控蛋白,并通過對基因表達網絡及生理過程的調控增強植株對逆境脅迫的耐受性[3]。LEA(late embryogenesis abundant proteins)是一類受水分脅迫誘導而大量表達的功能蛋白,其在植物適應逆境的過程中發揮非常重要的調控功能[4-6]。
脫水素(dehydrins,DHN)是LEA蛋白家族的重要成員之一,屬于LEA的D-Ⅱ家族,蛋白分子量大小在9 kDa到200 kDa之間[7];脫水素具有較強的親水性,大量表達的脫水素能夠束縛水分子,維持細胞液正常流動,從而具有保持細胞穩定的功能[8-9],在逆境脅迫條件下能夠起到維持細胞滲透平衡、替代水分子與細胞膜結合、增強膜結構穩定性等多種調節作用[10-11]。研究發現,脫水素在擬南芥、水稻、玉米、小麥等多種植物中廣泛存在。擬南芥脫水素AtHIRD11能夠通過與Cu2+結合,降低活性氧的生成,從而降低自由基的產生和過氧化毒害[12]。擬南芥脫水素COR15a對低溫脅迫比較敏感,能夠通過減弱底物的聚集而對酶的活性起到一定的保護作用[13-14]。大麥中的脫水素HvDHN5在干旱、低溫、鹽等逆境脅迫條件下均被誘導表達[15]。在小麥中,脫水素也是受水分脅迫誘導而上調表達,例如TaDHN-1、WDHN-1和WCOR410均為受脅迫誘導表達的脫水素,其可能通過依賴于ABA的信號通路在逆境響應過程中發揮重要功能[16-17];小麥WDHN1-2主要參與轉錄調控、生長因子和信號轉導[18]。目前,關于小麥中脫水素參與的逆境脅迫分子調控機制仍不十分清楚。
本課題組在前期研究中系統分析了小麥根系水分脅迫響應的基因差異表達譜[19],并鑒定出一個在水分脅迫過程中被大量誘導表達的脫水素基因(TaAffx.131747.1.S1_x_at)。在此基礎上,本研究擬克隆該脫水素基因,并系統分析其序列特征及生物學功能,以期為該基因進一步應用于小麥抗逆分子育種奠定基礎。
1 材料與方法
1.1 試驗材料及處理
選用洛旱2號和中國春作為試驗材料。在恒溫光照培養箱中,采用水培法育苗(26 ℃,光照16 h/8 h),幼苗培養至兩葉一心時分別進行干旱處理[20% PEG6000(w/v)]、鹽脅迫處理[NaCl濃度為100 mmol·L-1]和ABA處理(濃度為100 μmol·L-1),取材時間為處理0 h、0.5 h、1 h、2 h、6 h、12 h、24 h、48 h。所有材料經液氮速凍后置于-80 ℃條件下保存備用。
1.2 菌株、載體試劑
大腸桿菌(Escherichiacoli)DH5 、BL21購于大連TaKaRa公司。載體pMD19-T(TaKaRa,中國大連),pET-28a為本實驗室保存。TransZol Plant,熒光染料購于TransGen Biotech公司(北京),In-FusionTM Advantage PCR Cloning Kit,反轉錄試劑盒,La Taq、EcoR I、XhoI購于TaKaRa公司(中國大連),質粒提取試劑盒、HRP-DAB試劑盒購自天根生化科技(北京),HIS抗體購于Abcam公司(上海),山羊抗兔IgG二抗, HRP辣根過氧化物酶標記購于康為世紀。
1.3 RNA和gDNA提取
使用Trizol(Invitrotgen)法[20]提取總RNA,用1%瓊脂糖凝膠檢測RNA質量,利用Nanodrop2000核酸分析儀測定總RNA的濃度和純度。基因組DNA提取采用CTAB法。
1.4 小麥 TaDHN-2>基因的克隆和序列分析
根據課題組的前期研究結果[19],進行NCBI的序列比對和生物信息學分析,依據分析結果設計了特異性引物TaDHN-2-1(5′-CAAGTGAGCAAGACAACACACCATA-3′)和TaDHN-2-2(5′-CACCTCAAACTTTCACAAGTAGCGG-3′)用于cDNA片段的克隆,PCR擴增結果經1%的瓊脂糖凝膠電泳檢測后,進行DNA片段凝膠回收、純化,并連接到pMD19-T載體進行測序分析。蛋白質二級結構分析采用CSOPMA, CCD、Expasy在網站進行,用ProtScale進行疏水性分析并使用GraphPad prism軟件處理數據,用SignalP-4.0檢測信號肽區域,以PredictProtein預測跨膜區域,進行亞細胞內定位。使用MEGA5.0軟件對擬南芥的AtRED14(AT1G76180.1)、AtLTI29(CAA62448.1)、AtHIRD11(OAP19672.1) 和AtLTI30(CAA54704.1);水稻的OsRAB21(CAA68765.1) 和OsWSI724(BAA05539.1);玉米ZmRAB17(CAM56275.1);小麥TaWZY1-2(ABV24865)、TaWCS120(AAA34261.2)、TaDHN-1(AB272228.1);大麥的HvDHN3(AAD02255.1)和 HvDHN13(AAT81473.1)進行蛋白序列聚類分析。
1.5 小麥 TaDHN-2>基因的表達模式分析
依據克隆后測序的結果設計實時定量特異引物TaDHN-2-3(5′-GGCATTTCCAGCCCTCG-3′)和TaDHN-2-4(5′-ATGACGCCCTTCTTC TCG-3′);內參基因為β-actin(Accession No.AB181991),引物為actin-F(5′-GTTCCAATCTATGAGGGATACACGC-3′)和actin-R(5′-GAACCTCCACTGAGAACAACATTACC-3′)。表達特性分析采用實時定量RT-PCR的方法,使用Bio-Rad公司的IQ5 PCR 儀,程序為95 ℃預變性1 min;95 ℃變性10 s,60 ℃延伸35 s,35個循環。依據實時定量PCR獲得的內標基因β-actin和目的基因的Ct值,按照2-ΔΔCt法[21]計算目的基因的相對表達量,進行3次生物學重復。
1.6 原核表達載體構建和誘導表達
依據前期保存的pET-28a質粒,設計引入酶切位點EcoR I、XhoI的PCR引物,TaDHN-2-5(5′-CGCGAATTCCACCTGTGCAAGATGG-3′;圖1下劃線部分為EcoRI酶切位點)和 TaDHN-2-6(5′-TGCTCGAGTCTTTCACAAGTAGCGG-3′;圖1下劃線部分為XhoI酶切位點)。對PCR回收產物和原核表達載體pET-28a的質粒做EcoRI 和XhoI雙酶切,經連接后將重組載體轉化大腸桿菌DH5 ,陽性克隆進行PCR驗證后提取質粒再次酶切驗證并測序。將重組載體質粒轉入大腸桿菌BL21中,挑取陽性克隆,接種于1 mL含有卡那霉素(50 mg·L-1)的LB液體培養基中過夜培養,次日按1∶100的比例擴大培養,當OD600到0.6時,分別加IPTG至終濃度1 mmol·L-1過夜培養。
1.7 重組目的蛋白的純化和對 α-淀粉酶活性影響分析
上述過夜菌液12 000 rpm離心收集菌體,1 g菌體加入4 mL結合緩沖液(10 mmol·L-1咪唑、50 mmol ·L-1NaH2PO4、300 mmol·L-1NaCl、pH 8.0),懸浮菌液。冰上10 s間隔超聲破碎裂解菌體,4 ℃、12 000 r·min-1離心45 min收集上清液,上清液用0.45 μm的濾膜過濾,經鎳柱純化目的蛋白。以Bradford 法[22]測定蛋白濃度。使用Western blot法[23]鑒定目的蛋白,經SDS-PAGE后,將目的蛋白所在的分離膠轉移到硝酸纖維素膜,在含有0.5%脫脂奶粉的TBST下浸泡過夜。用0.5% SMP的TBST稀釋HIS抗體(購于Abcam公司)孵育1 h,TBST洗滌干凈后,使用0.5% SMP稀釋酶標記的第二抗體繼續孵育1 h。洗滌后使用天根公司的HRP-DAB試劑盒顯色,記錄試驗結果。
取α-淀粉酶提取液1 mL,加入目的蛋白,終濃度分別為0、10、20、60、100 μg·mL-1,每個濃度兩個重復,然后加入1 mL pH5.6檸檬酸。60 ℃和4 ℃分別水浴15 min后,加入等溫的淀粉溶液5 mL。水浴5 min后試驗組加入4 mL 0.4 mol·L-1NaOH終止反應,取1 mL反應液加入1 mL 3,5-二硝基水楊酸煮沸5 min,稀釋至15 mL,測定OD510,計算。對照組用等體積500 mmol·L-1咪唑洗脫緩沖液代替目的蛋白溶液,并在水浴前加入4 mL 0.4 mol·L-1NaOH鈍化酶活性。
2 結果與分析
2.1 小麥脫水素基因 TaDHN-2>的克隆及其序列分析
分別以PEG處理6 h后洛旱2號三葉期葉片的cDNA和gDNA為模板,用特異引物(TaDHN-2-1和TaDHN-2-2)進行PCR擴增(圖1A,1B)。測序結果表明,以cDNA為模板的克隆基因大小為556 bp,其中5′ UTR 59 bp,3′UTR 38 bp,ORF大小為 459 bp。與cDNA序列比對,其gDNA包含一個109 bp的內含子,符合GC/AG的剪切機制。進一步分析顯示,克隆基因可編碼152個氨基酸,分子量約15.52 kDa,等電點是7.17。氨基酸組成分析發現,其富含親水性氨基酸Gly(G)、Thr(T)、Gln(Q)、Lys(K)(圖1C),不含有疏水性氨基酸Cys(C)、Trp(W)。經NCBI數據庫Blastp檢索,其與傘穗山羊草(Aegilopsumbellulata)中的脫水素基因CAJ56057.1序列同源性為81%,表明克隆的基因為脫水素基因,將其命名為 TaDHN-2>。

A: TaDHN-2> 的cDNA擴增結果;B: TaDHN-2>的gDNA擴增結果;A、B圖中的1和M分別指 TaDHN-2>目的片段和DL2000 DNA Marker;C: TaDHN-2>氨基酸組成及含量分析;D: TaDHN-2>序列特性。A:cDNA products of the TaDHN-2>; B:gDNA amplification of the TaDHN-2>; 1 and M in chart A and B refer to target fragment of TaDHN-2 and DL2000 DNA marker, respectively; C:Analysis of amino acid composition and content of TaDHN-2>; D:The DNA and protein sequences of TaDHN-2>.
2.2 小麥脫水素TaDHN-2的結構特性分析
二級結構預測顯示,小麥脫水素TaDHN-2可形成 α-螺旋、β-折疊、β-轉角、無規則卷曲,占氨基酸比例分別為16.45%(25)、9.21%(14)、12.50%(19)、61.84%(94)(圖2A)。小麥脫水素TaDHN-2的親水性強(圖2B),沒有信號肽區域(SignalP-4.0)和跨膜區域,亞細胞定位到細胞質中。系統進化分析表明,小麥脫水素TaDHN-2與擬南芥AtRAB18和水稻OsRAB21同屬脫水素的YnSkn亞類(圖2C)。經進一步分析,小麥脫水素TaDHN-2的37~151氨基酸位點為脫水素保守結構域(圖1D),具有YnSKn類脫水素典型保守結構,包括N端的Y片段(16~22,VDEYGNP)、S片段(59~74,SSSSSEDDGMGGRRKK)以及C端的兩個K片段保守結構域(79~86和143~150; KIKEKLPG)。
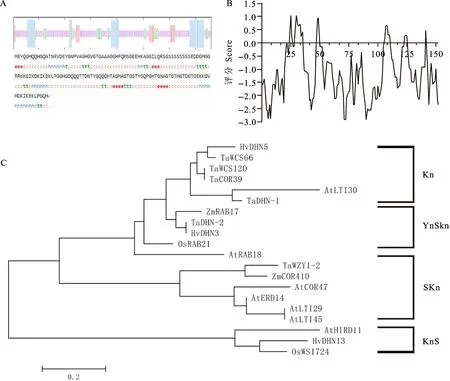
A:TaDHN-2的二級結構預測,h: α-螺旋,t:β-轉角,e:伸展鏈,c:無規則卷曲;B:TaDHN-2蛋白的氨基酸序列親/疏水性預測;C:TaDHN-2與其他脫水素蛋白的系統進化分析。A:Secondary structure of TaDHN-2, h:Alpha-helix, t:Beta-turn, e:Extended strand, c:Random coil; B:hydrophily/hydrophobicity of TaDHN-2; C:Phylogentic tree of TaDHN-2 and other dehydrins.
2.3 小麥脫水素基因 TaDHN-2>對脫落酸ABA的響應
在100 μmol·L-1ABA處理1 h后, TaDHN-2>在中國春根系中的表達量最高,是對照的5.3倍。ABA處理之后的2~48 h,其表達量與對照沒有明顯差異。在洛旱2號根系中,在ABA處理后,隨時間的延長, TaDHN-2>表達量逐漸增高,在處理6 h時達到最高,是對照的6.3倍,之后其表達量逐漸下降(圖3A)。在中國春葉中的表達模式與根系相似,而洛旱2號葉片中 TaDHN-2>基因在在ABA誘導后2 h增強表達,在ABA處理12 h時其表達量快速增高到4.9倍,達到最高(圖3B)。總體上看來,在ABA誘導下根系的 TaDHN-2>表達模式與葉片相近,但是其在洛旱2號中對ABA脅迫響應速度慢于中國春,但整體持續增強表達時間較長,這可能與洛旱2號具有較強耐旱能力有關。
2.4 小麥脫水素基因 TaDHN-2>對滲透脅迫的響應
在20% PEG6000處理6 h后,中國春根系 TaDHN-2>的表達量達到最高,是對照的10倍。處理12~48 h表達量緩慢下降到對照水平。而洛旱2號根系的 TaDHN-2>表達量在20% PEG6000處理0.5 h時達到最高,為對照2.3倍,之后其表達量與對照沒有顯著差異(圖4A)。在洛旱2號和中國春的葉中, TaDHN-2>具有相似的表達模式,但其在洛旱2號中的相對表達量變化較中國春更為明顯,在20%PEG6000處理1 h時, TaDHN-2>在兩個品種中的表達量均最高,洛旱2號和中國春中的表達量分別為對照的5.3和3.8倍(圖4B)。
鹽脅迫過程中,中國春根系的 TaDHN-2>表達量呈波動性變化,在處理2 h、48 h時表達量出現兩個峰值,分別是對照的5和4.8倍,但在洛旱2號根系中該基因表達變化不明顯(圖4C)。在葉片中,洛旱2號的 TaDHN-2>表達量在鹽處理后6 h時最高,是對照的5倍;而中國春的 TaDHN-2>表達量在不同處理時間與對照無明顯差異(圖4D)。
綜上所述, TaDHN-2>在2個品種中具有不同的誘導表達模式,在根系中在中國春中的誘導表達量顯著高于洛旱2號,而在洛旱2號葉片中的誘導表達量更高。
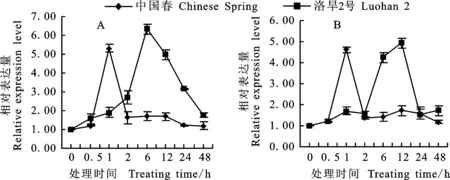
圖3 ABA處理下小麥 TaDHN-2>在小麥根(A)和葉片(B)中的表達模式Fig.3 Expression pattern of TaDHN-2 in wheat root(A) and leaf(B) under ABA treatment
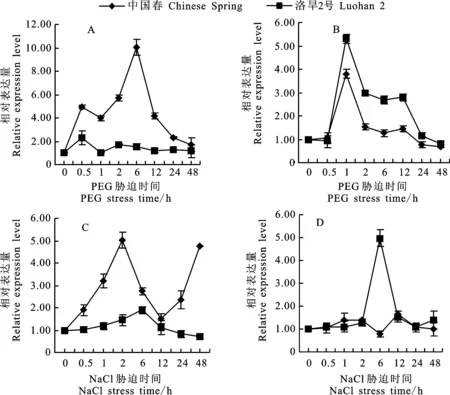
圖4 滲透脅迫下小麥 TaDHN-2>在根(左)和葉片(右)中的表達模式Fig.4 Expression profile of TaDHN-2 in wheat root(leaf) and leaf(right) under osmotic stress
2.5 小麥TaDHN-2對酶活性的保護功能分析
將包含完整ORF的目的基因插入pET-28a表達載體MCS區域的3′末端,構建了原核表達載體,PCR鑒定篩選陽性克隆(圖5A)。IPTG誘導表達HIS-TaDHN-2融合蛋白,鎳柱純化后,Western blot結果顯示,在15 kDa有雜交條帶(圖5B),表明HIS-TaDHN-2純化成功。
同α-淀粉酶提取液中添加融合蛋白,結果表明,在60 ℃下,α-淀粉酶活性隨著HIS-TaDHN-2濃度的增加而提高;在融合蛋白的濃度為100 μg·mL-1時, α-淀粉酶的活性提高到對照的2倍(圖6A)。在4 ℃條件下,融合蛋白濃度低于20 μg·mL-1時,HIS-TaDHN-2對α-淀粉酶活性沒有影響;當融合蛋白濃度達到60 μg·mL-1時, α-淀粉酶活性顯著增強,并隨加入融合蛋白濃度的增大而提高,當加入濃度為100 μg·mL-1, α-淀粉酶活性提高為對照的1.6倍(圖6B)。綜合上述結果說明,TaDHN-2在60 ℃和4 ℃可以提高α-淀粉酶的活性,對溫度引起的酶活性下降具有一定的保護作用。
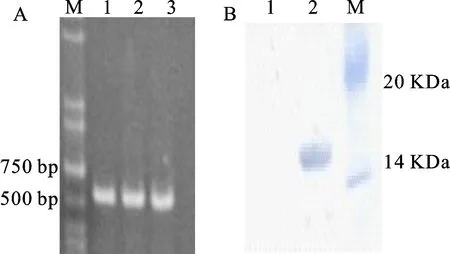
A圖中,M表示DL2000 DNA marker;1~3:單克隆菌液。B圖中,M:94 kDa;2:目的蛋白。In chart A,M:DNA marker DL2000;1-3:Bacteria liquid of monoclonal;In chart B,M:94 kDa;2:Target protein.
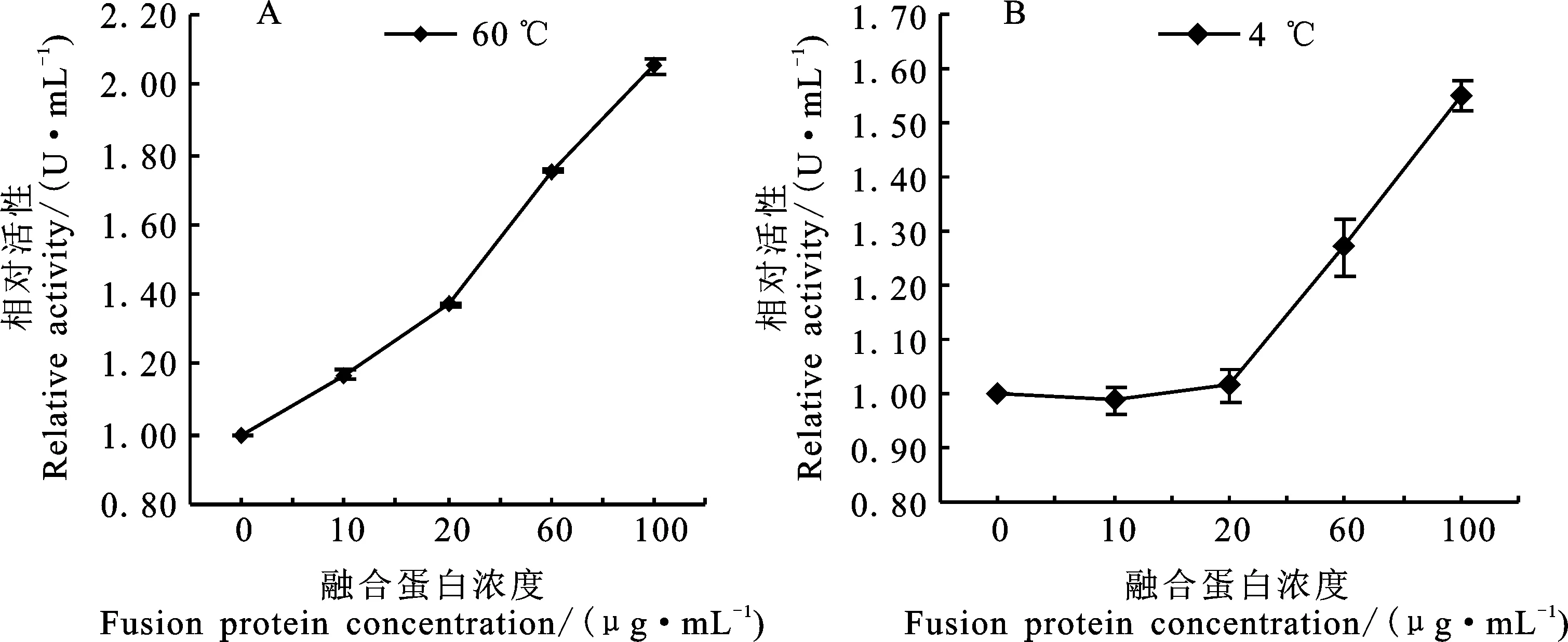
圖6 融合蛋白HIS-TaDHN-2對α-淀粉酶活性的影響Fig.6 Effect of HIS-TaDHN-2 on the activity of α-amylase
3 討 論
逆境脅迫嚴重影響著植物生長發育的進程[24]。脫水素是一類在逆境脅迫條件下(如干旱和高鹽等)大量表達的功能蛋白[7]。DHN蛋白通常含有高度保守的C端K片段[EKKGIME/DKIKEKLPG]、N端的Y片段[保守的(V/T)D(E/Q)YGNP]和中間的4~10個絲氨酸殘基串的S片段[LHRSGS4~10(E/D)],根據三個保守區域的組成不同,將脫水素分成SKn、Kn、YnSKn、KnS和YnKn五類[7,25]。本研究克隆的 TaDHN-2>的編碼蛋白包含兩個K片段保守結構域(C端)、1個Y片段和1個S片段保守域(N端),屬于典型的YnSKn類脫水素。研究發現,脫水素K片段可以形成與A2相似的雙親性α-螺旋并與蛋白質的疏水作用位點結合,避免蛋白質的不可逆變性和聚集[26-27]。K片段保守域有可能與 TaDHN-2>在抗逆過程中的響應密切相關[28]。另外, TaDHN-2>還有N端豆蔻酰化位點,CK2磷酸化位點,和酰胺化位點,這些位點也可能在脫水素行使功能的過程中發揮重要作用,但具體的機理還有待進一步研究。
植物的非生物脅迫響應包括ABA的依賴型和非依賴型兩個類型,前者在干旱等細胞脫水脅迫過程中發揮了極其重要的功能[29]。在本實驗所用的兩個小麥品種中, TaDHN-2>在ABA的誘導下均上調表達,證明該基因屬于ABA依賴型的非生物脅迫響應途徑。脫水素在不同的組織、器官以及不同的逆境環境下有不盡相同的表達模式。如大麥中的YnSKn型脫水素基因 DHN12>只在胚胎發育過程中表達,而營養組織中不表達[30];Yang等[31]對葡萄的多個脫水素基因進行研究發現,干旱條件下只有DHN1表達;本研究結果表明, TaDHN-2>在小麥根系和葉片中均可被誘導表達,但是抗旱品種洛旱2號中其被誘導表達的時間明顯長于中國春,推測這可能與洛旱2號抗逆性強的品種特性有關。
研究發現,小麥WRAB18、玉米ZmLEA3和高粱SbDhn2脫水素蛋白K片段形成的α-螺旋結構可能與變性的LDH疏水基團作用,防止LDH進一步變性,以維持乳酸脫氫酶的正常功能[32-35]。Nakayama等[9]在對Cor15am的研究中發現,其在溫度低于43 ℃的條件下對酶底物具有一定的保護作用。本研究發現,TaDHN-2融合蛋白的濃度為100 μg·mL-1時,在溫度為60和4 ℃的條件下均能有效提高α-淀粉酶活性,這說明TaDHN-2蛋白對維持酶活性的功能具有積極作用,這是否與逆境脅迫下功能蛋白的活性保護有關仍有待于進一步證實。
