一種米曲霉耐鹽蛋白酶的純化及酶學性質分析
2019-01-07 02:38:04毛丙永劉艷鳳趙國忠崔樹茂趙建新
食品科學 2018年24期
毛丙永,劉艷鳳,趙國忠,崔樹茂,趙建新,*
(1.江南大學食品學院,江蘇 無錫 214122;2.天津科技大學食品工程與生物技術學院,天津 300457)
米曲霉是目前醬油釀造中最重要的菌株,由于其不產毒素,且胞外蛋白酶系十分復雜[1],是分解大豆蛋白的最佳選擇[2],能夠賦予醬油獨特的風味[3]。米曲霉的胞外蛋白酶系通過頂端分泌的方式分泌至胞外[4-5],其基因組中有135 個胞外蛋白酶基因[6],在醬油發酵期間,蛋白酶之間交替發生作用。米曲霉產生的蛋白酶如此重要,引起學者們的廣泛關注。Su Guowan等[7]研究發現,米曲霉蛋白酶粗提物水解脫脂大豆的能力優于堿性蛋白酶、復合蛋白酶和木瓜蛋白酶等商業蛋白酶。微生物生產得到的工業蛋白酶,約75%被用于烘焙、釀造、發酵、明膠、肉類、皮革和洗滌劑等領域[8]。在醬油發酵后期,NaCl質量濃度高達12~18 g/100 mL,許多蛋白酶的活性被抑制,只有一些耐鹽的蛋白酶仍保持較高的酶活力,導致原料水解不徹底,發酵周期被延長。因此,從米曲霉中純化得到耐鹽蛋白酶,對于指導醬油發酵具有重要的意義。本實驗旨在從米曲霉3.042中,挖掘一種具有潛在工業價值的耐鹽蛋白酶,從而指導生產實踐。
1 材料與方法
1.1 材料與試劑
米曲霉3.042保藏于江南大學食品生物技術中心;Q-HP陰離子柱、Superdux 75凝膠柱 通用電氣醫療集團;BCA蛋白濃度測定試劑盒 上海碧云天有限公司;福林-酚、40%聚丙烯酰胺-甲叉雙丙烯酰胺(29∶1)、胰蛋白酶(均為分析純) 生工生物工程(上海)股份有限公司。
1.2 儀器與設備
AKTA avant全自動蛋白快速分離系統 瑞典GE公司;ultrafleXtreme基質輔助激光解析電離串聯飛行時間質譜儀 美國布魯克·道爾頓公司;MOS-450圓二色譜儀 法國Biologic公司;UV2450型分光光度計日本島津公司。
1.3 方法
1.3.1 種曲培養條件
稱取100 g麥麩,添加110 mL蒸餾水,按照30 g/500 mL三角瓶分裝,121 ℃滅菌20 min。待溫度冷卻至40 ℃以下,接入米曲霉,搖勻,30 ℃堆積培養,待瓶內泛白后搖瓶培養,生長至黃綠色時,將種曲裝入牛皮紙袋中,密封,50 ℃烘干,4 ℃貯存備用。
1.3.2 大曲培養條件
稱取60 g豆粕,添加110 g沸水浸泡30 min后,加入40 g麩皮,攪拌均勻,121 ℃滅菌20 min,按照質量分數2.5‰接種種曲,28 ℃、相對濕度90%培養,控制溫度低于40 ℃,待大曲變為黃色,立即收集并提取蛋白酶。
1.3.3 粗酶液提取
稱取一定量的大曲,按照料液比為1∶20添加0.9%無菌生理鹽水,40 ℃水浴1 h,不斷攪拌,紗布過濾,4 ℃、8 000×g離心20 min,收集上清液。
1.3.4 硫酸銨分級沉淀
參考Wang Dong等[9]的方法。量取一定體積的粗酶液,低溫攪拌加入硫酸銨,使之飽和度分別達到10%、20%、30%、40%、50%、60%、70%、80%和90%,每一飽和度在4 ℃靜置4 h,9 000×g離心20 min,20 mmol/L pH 7.5 Tris-HCl重懸沉淀,使用8~14 kDa透析袋透析脫鹽,使用聚乙二醇20000濃縮樣品,凍干冷藏備用。
1.3.5 Q-HP陰離子交換
上樣緩沖液A為20 mmol/L pH 7.5 Tris-HCl,洗脫液B為1 mol/L NaCl,流速為2 mL/min,洗脫條件:100% A,0% B,15 BV;90% A,10% B,15 BV;20% B,15 BV。收集樣品測定酶活力,做十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(sodium dodecyl sulfatepolyacrylamide gel electrophoresis,SDS-PAGE)。
1.3.6 Superdux 75凝膠層析
用20 mmol/L pH 7.5 Tris-HCl平衡柱子,并洗脫,流速為0.4 mL/min,收集樣品測定酶活力,做SDS-PAGE。
1.3.7 蛋白酶活力測定
參考SB/T 10317—1999《蛋白酶活力測定法》的方法。取待測樣品1 mL以及質量濃度2 g/100 mL酪蛋白溶液,pH 7.2,40 ℃水浴2 min,取出立即加入1 mL酪蛋白,40 ℃水浴反應10 min,加入2 mL 0.4 mol/L三氯乙酸溶液,水浴20 min使蛋白質沉淀,隨后使用0.22 μm針頭式濾器過濾,然后取1 mL濾液,加入5 mL 0.4 mol/L Na2CO3溶液,加入1 mL稀釋3 倍的福林-酚試劑混勻,40 ℃水浴20 min,在波長660 nm處測定光密度。40 ℃時每分鐘水解酪蛋白生成1 μg酪氨酸,被定義為1 個蛋白酶活力單位(U)。1.3.8 蛋白質量濃度測定
采用BCA法測定蛋白質量濃度,以牛血清蛋白作為標準蛋白。
1.3.9 蛋白酶SDS-PAGE分析
參考Laemmli[10]的方法。濃縮膠質量濃度為5 g/100 mL,分離膠質量濃度為12 g/100 mL,利用考馬斯亮藍R250染色。
1.3.10 飛行時間質譜鑒定蛋白酶
參考Vejvoda等[11]的方法。將上述SDS-PAGE的蛋白條帶切下,切成1 mm3的小膠塊,置于1.5 mL EP管中,加入200~400 μL 100 mmol/L NH4HCO3/30% ACN混合溶液脫色,清洗至透明,去除上清液,用50 μL ACN脫水2 次,得到白色膠粒。加入5 μL 2.5~10 ng/μL胰蛋白酶溶液,4 ℃放置30~60 min,使膠塊充分溶脹,吸出剩余酶液。再加入20~30 μL 25 mmol/L NH4HCO3緩沖液,37 ℃水浴20 h。吸出酶解液,冷凍濃縮,點樣,進行質譜分析。檢索參數見表1。

表1 檢索參數設置Table 1 Parameters of database search
1.3.11 蛋白酶三級結構預測
飛行時間質譜鑒定得到的蛋白序列,在NCBI(https://www.ncbi.nlm.nih.gov/)中進行比對,得到蛋白酶的氨基酸序列,將此序列在Swiss model(http://www.swissmodel.expasy.org/)中進行模型擬合,按照GMQE值選擇匹配度較高的模型。
1.3.12 蛋白酶最適溫度和溫度穩定性測定
參考Lee等[12]的方法。pH 7.2時,在20、30、40、50、60、70 ℃和80 ℃測定蛋白酶活力,計算不同溫度條件下的相對酶活力,酶活力最高的溫度即為最適溫度。
pH 7.2時,把蛋白酶放置于35、40、45、50 ℃和55 ℃分別保溫20、40、60、80 min和100 min,在40 ℃、pH 6.5條件下測定其蛋白酶活力,保溫前的酶活力被定義為100%,計算其他條件下的相對酶活力,酶活力最高的溫度即為最適溫度。
1.3.13 蛋白酶最適pH值和pH值穩定性測定
參考Lee等[12]的方法。利用pH 4.5、5.0和pH 5.5檸檬酸緩沖液,pH 6.0、6.5、7.0、7.5、8.0、8.5和pH 9.0 Tris-HCl緩沖液配制質量濃度2 g/100 mL酪蛋白溶液,并調節酶液至相應的pH值,在40 ℃測定不同pH值條件下蛋白酶活力,計算不同pH值的相對酶活力,最高酶活力的pH值即為最適pH值。
將調至pH 6.0、6.5、7.0、7.5、8.0、8.5和pH 9.0的酶液,40 ℃分別保溫30、60、90 min以及120 min,在40 ℃、pH 6.5條件下測定其蛋白酶活力,保溫前的酶活被定義為100%,最高酶活的pH值即為穩定pH值,計算其他條件下的相對酶活力。
1.3.14 金屬離子對蛋白酶活力的影響
配制NaCl、CaCl2、MnSO4、CuCl2、FeSO4、FeCl3和KCl溶液,與蛋白酶混合使金屬離子終濃度達到2 mmol/L,在40 ℃、pH 7.0條件下測定酶活力,不添加金屬離子測得的酶活力定義為100%,計算不同金屬離子條件下的相對酶活力。
1.3.15 蛋白酶動力學特征測定
參考Lineweaver等[13]的方法。配制1.0、3.0、5.0、7.0、9.0、11.0 g/L和13.0 g/L的酪蛋白溶液,在40 ℃、pH 7.0條件下測定其反應速率,然后繪制1/S-1/V的Lineweaver-Burk雙倒數曲線,根據曲線的橫截距為-1/Km,縱截距為1/Vm,求出蛋白酶的米氏常數Km以及最大反應速率Vm。
1.3.16 金屬離子影響下蛋白酶二級結構測定
參考Sreerama等[14]的方法。樣品蛋白質量濃度范圍0.01~0.2 mg/mL,添加NaCl、CaCl2、MnSO4、CuCl2、FeSO4、FeCl3和KCl溶液,使金屬離子終濃度為2 mmol/L,掃描波長190~250 nm,掃描速率100 nm/min,光程1 mm,使用SELCON3進行分析。
1.3.17 蛋白酶的耐鹽特性
蛋白酶液中加入NaCl溶液,使其終濃度質量分別為5、10 g/100 mL和15 g/100 mL,在pH 7.0,40 ℃的條件下測定酶活力,不添加Na+測得的酶活力定義為100%,計算不同濃度Na+條件下的相對酶活力。
1.4 數據分析
實驗結果數據均以 ±s表示(n=3),圖形采用Origin 8.5軟件繪制。
2 結果與分析
2.1 米曲霉蛋白酶的分離純化

圖1 蛋白酶的鹽析曲線Fig. 1 Salting-out profile of protease
酶活力標準曲線:y=151.81x-1.582 4,R2=0.999 1。蛋白質量濃度標準曲線:y=0.507 2x+0.001 4,R2=0.999 7。由蛋白酶的鹽析曲線(圖1)可以看出,隨著硫酸銨飽和度的增加,析出的蛋白酶活力呈現出增加的趨勢,尤其是當硫酸銨飽和度達到70%、80%、90%時,酶活力分別達到(1 471.11±1.01)、(1 451.02±0.71)、(1 426.54±0.67) U/mL,但是比酶活并沒有顯著高于其他飽和度,故選用30%~90%飽和度進行分級沉淀,提高回收率。
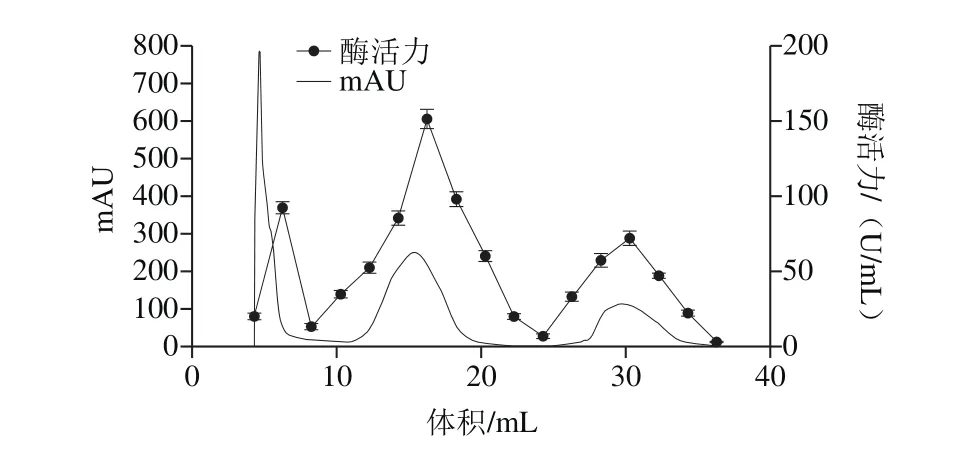
圖2 蛋白酶的Q-HP洗脫曲線Fig. 2 Elution profile of protease on Q-HP column
如圖2所示,檢測各個峰的酶活力顯示,第2個峰酶活力最高,達(151.51±4.41) U/mL,SDS-PAGE結果顯示有2條明顯條帶,故收集合并第2個峰的酶液,使用30 kDa超濾離心管濃縮,進行下一步純化。
如圖3所示,凝膠層析分離效果較好,收集第1個峰的樣品檢測其具有蛋白酶活力,SDS-PAGE呈現單一條帶。Li Caihong等[15]利用硫酸銨分級沉淀,離子交換柱和凝膠層析聯合使用,從米曲霉FC76中純化得到一種酸性蛋白酶。Rafael等[16]通過乙醇沉淀,凝膠層析和離子交換柱聯合使用,從米曲霉中純化出一種絲氨酸蛋白酶。也由此可以看出,離子交換柱和凝膠層析聯合使用,是蛋白酶純化過程中常用的技術手段。
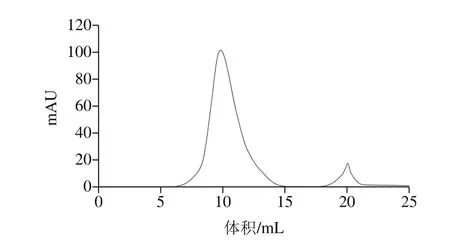
圖3 蛋白酶的Superdux 75洗脫曲線Fig. 3 Elution profile of protease on Superdux 75 column
如表2所示,隨著純化步驟的增加,蛋白的比酶活和純化倍數呈增長的趨勢,Superdux 75凝膠層析后比酶活達到1 422.76 U/mg,純化倍數為94.47,回收率為2.73%。
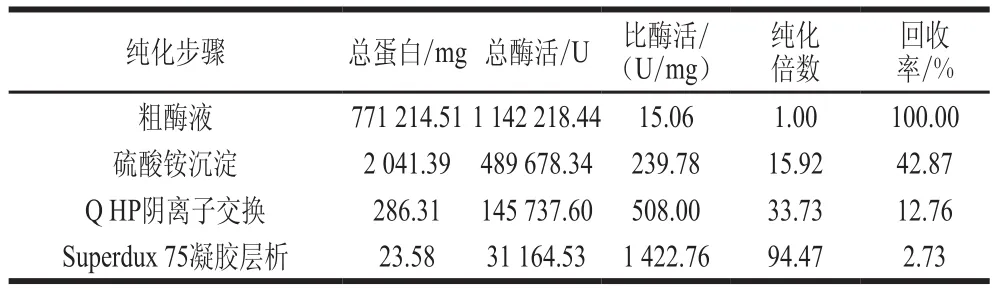
表2 蛋白酶純化結果Table 2 Purification of the protease
2.2 蛋白酶SDS-PAGE分析

圖4 蛋白酶的SDS-PAGE分析Fig. 4 SDS-PAGE analysis of the protease
由圖4可以看出,純化得到的蛋白酶達到電泳純,分子質量約為93.33 kDa。
2.3 飛行時間質譜鑒定蛋白酶
飛行時間質譜鑒定該蛋白酶可能為鈣蛋白酶RIM13(Gene ID:AO090005000756),是一種半胱氨酸蛋白酶,分子質量為88.89 kDa,該蛋白酶于2012年在米曲霉3.042中被鑒定[17]。米曲霉3.042中也曾純化得到中性蛋白酶,分子質量分別為50 kDa[18]、27 kDa和58 kDa[19]、37 kDa和45 kDa[20]。鈣蛋白酶RIM13首次從米曲霉3.042中分離得到,安全特性高,可以直接應用于醬油工業發酵。鈣蛋白酶是一種信號蛋白酶,在堿性環境中,通過轉錄因子PacC,分泌堿性蛋白和異青霉素N-合成酶等,提高菌體在高pH值環境中的適應性[21]。半胱氨酸的活性中心是Cys-His-Asn三聯體[22],應用廣泛,也被用于降解乳品工業廢棄物,如干酪乳清等[23]。
2.4 蛋白酶三級結構預測結果
蛋白酶在Swiss model中匹配到的結果是鈣蛋白酶[24],GMQE值為0.33,該蛋白酶包含3 個肽段,分別為鈣蛋白酶-2催化亞基、鈣蛋白酶小亞基,鈣蛋白酶抑制蛋白,以及10 個非共價的Ca2+配體(圖5)。三級結構的表征更有助于從結構上驗證該蛋白酶的性質。
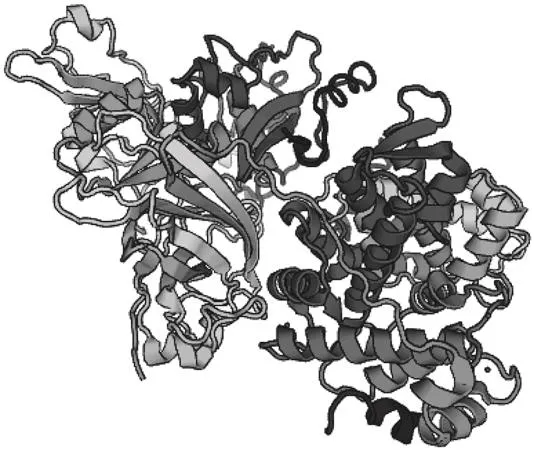
圖5 蛋白酶的三級結構預測Fig. 5 Predicted tertiary structure of the protease
2.5 蛋白酶的酶學性質
2.5.1 蛋白酶的最適溫度及溫度穩定性

圖6 蛋白酶的溫度-相對酶活力曲線Fig. 6 Optimum temperature of the protease
如圖6所示,在20~50 ℃,蛋白酶活力呈現增加的趨勢,在50 ℃時酶活力最高,為(90.28±0.76) U/mL。當溫度升至50 ℃以上時,酶活力迅速降低,80 ℃時酶活力降至(0.61±0.30) U/mL,這也表明此蛋白酶不耐高溫,60 ℃以上酶幾乎失活。如圖7所示,該蛋白酶在35 ℃和40 ℃時蛋白酶保持較穩定,35 ℃和40 ℃保持100 min,酶活力分別保留
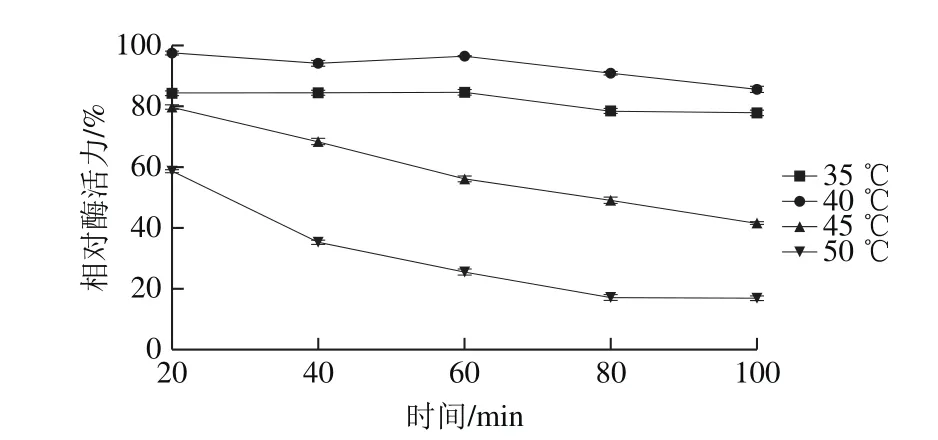
圖7 蛋白酶的溫度-相對酶活力穩定性曲線Fig. 7 Effect of temperature on the activity of the protease
77.96 %和85.70%。在45 ℃和50 ℃時隨著保溫時間延長,蛋白酶活力迅速下降,保溫100 min后,酶活力分別保留41.70%和16.95%,表明該蛋白酶在此溫度下不能穩定保存。
2.5.2 蛋白酶的最適pH值及pH值穩定性
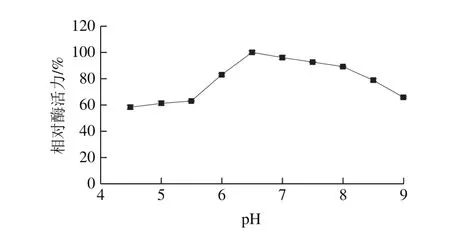
圖8 蛋白酶的pH值相對酶活力曲線Fig. 8 Optimum pH profile of the protease
如圖8所示,pH 4.5~6.5,酶活力呈現增加的趨勢,pH 6.5時酶活力最高,達到(89.06±1.07)U/mL。當pH值繼續升高時,酶活力呈下降趨勢,在pH 9.0時酶活力下降至(58.70±0.70)U/mL,故此蛋白酶最適pH值為6.5。每一種酶均具有最適pH值,是由于Hofmeister效應,在不同pH值的體系中,酶表面的氨基基團和羧基基團的電荷分布受到影響,進而影響酶與底物的結合以及催化效率[25]。
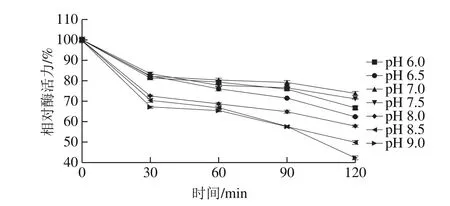
圖9 蛋白酶的pH值-相對酶活力穩定性曲線Fig. 9 Effects of pH on the activity of the protease
將此蛋白酶在不同pH值,40 ℃水浴不同時間測定其酶活力,如圖9所示,在不同pH值隨著水浴時間延長,酶活力均呈現降低的趨勢,蛋白酶在不同pH值保溫120 min時,相對酶活力最高的是pH 7.0,然后依次是pH 7.5、6.0、6.5、8.0、8.5、9.0,故此蛋白酶屬于中性蛋白酶。
2.5.3 金屬離子對蛋白酶活力的影響
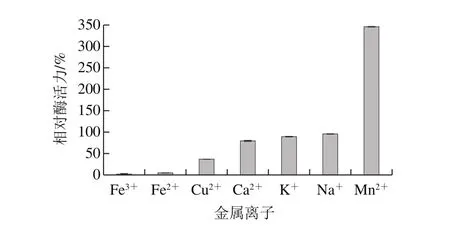
圖10 金屬離子對蛋白酶活力的影響Fig. 10 Effects of metal ions on the activity of the protease
如圖10所示,Fe3+、Fe2+、Cu2+、Ca2+、K+和Na+對蛋白酶呈現出抑制作用,且呈現減弱的趨勢。Mn2+對蛋白酶活力呈現促進作用,酶活力是不添加金屬離子時的(3.46±1.07)倍。添加Ca2+時,蛋白酶的酶活力是不添加金屬離子時的(0.80±0.1)倍,這可能是由于鈣蛋白酶抑制蛋白與鈣蛋白酶結合,降低鈣蛋白酶活力。真菌中的中性蛋白酶,一般可以被金屬離子螯合劑抑制[26]。
2.5.4 金屬離子對蛋白酶二級結構的影響
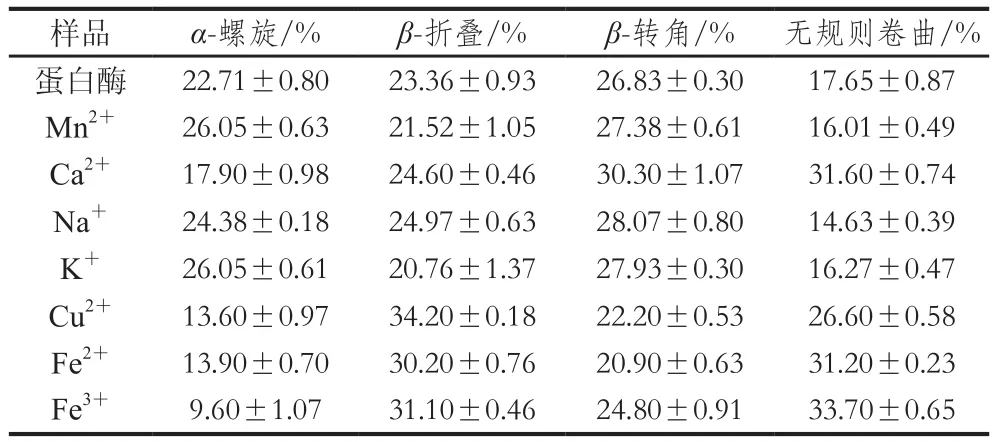
表3 金屬離子對蛋白酶二級結構的影響Table 3 Effects of metal ions on the secondary structure of the protease
樣品蛋白質量濃度為0.09 mg/mL,不同金屬離子對蛋白酶結構的影響結果如表3所示,Mn2+促進蛋白酶活,使蛋白酶的α-螺旋增加,β-折疊減少,β-轉角增加和無規則卷曲減少。可能這種結構變化促進蛋白酶與底物結合,使得蛋白酶呈現增加的趨勢。Ca2+使蛋白酶的α-螺旋減少,β-折疊增加,β-轉角增加和無規則卷曲增加。Na+是影響酶活的重要因素,Na+使蛋白酶α-螺旋增加,β-折疊增加,β-轉角增加和無規則卷曲減少。K+、Cu2+、Fe2+也均使蛋白酶的結構發生不同程度的變化。Fe3+幾乎完全抑制了蛋白酶的酶活,Fe3+使蛋白酶的二級結構發生很大的改變,其中α-螺旋由22.71%下降到9.60%,β-折疊增加了7.74%,無規則卷曲增加了16.05%,β-轉角減少了2.03%,如此巨大的結構變化使得該蛋白酶的結構紊亂,酶活幾乎喪失。米曲霉中一種中性蛋白酶在NaCl的環境中,半衰期增加,這是由于在高鹽環境中中性蛋白酶的吉布斯自由能和活化能均升高的原因,同時,在加入NaCl后中性蛋白酶二級結構中的α-螺旋也發生明顯變化。雖然高鹽環境改變了蛋白酶原本的結構,但是半衰期的延長也具有一定的應用價值。2.5.5 蛋白酶的動力學特征
根據1/S-1/V的Lineweaver-Burk雙倒數曲線(圖11),計算得到蛋白酶水解酪蛋白時米氏常數Km為2.43 g/L,最大反應速率Vm為103.09 mg/(L·min)。Km值較低,表明此蛋白酶與酪蛋白親和力較高,蛋白酶的活性中心容易與酪蛋白結合生成酶-底物復合中間體。
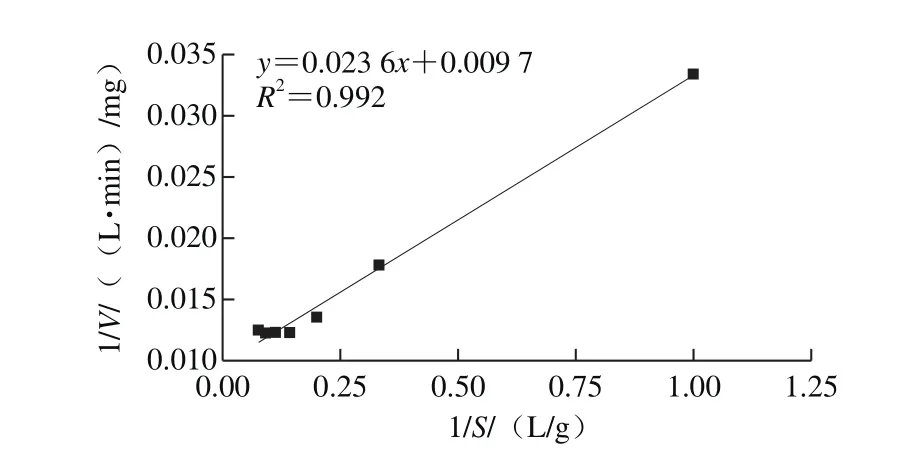
圖11 蛋白酶的雙倒數曲線Fig. 11 Lineweaver-Burk plot of the protease
2.5.6 蛋白酶的耐鹽性
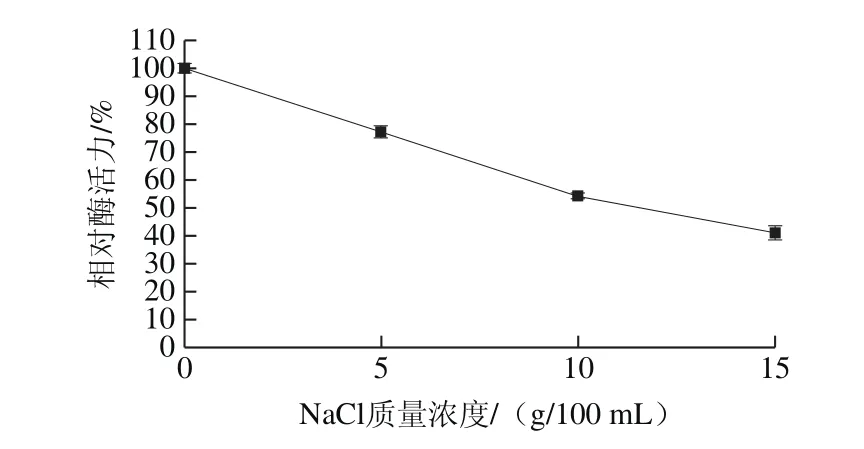
圖12 鹽度對蛋白酶活力的影響Fig. 12 Effect of salinity on the activity of the protease
由圖12可知,隨著NaCl質量濃度的上升,蛋白酶活力呈現下降的趨勢,酶液中NaCl質量濃度分別為5、10 g/100 mL和15 g/100 mL時,相對酶活力分別為77.22%、54.39%和41.15%。鹽度是影響酶活力的重要因素,醬油大曲中提取的粗酶液,在18% NaCl環境中酶活力僅能保持大約3%[27]。有學者從米曲霉LK-101中純化得到一種耐鹽蛋白酶,該蛋白酶在5、10 g/100 mL和15 g/100 mL NaCl環境中酶活力分別能保持80%、50%以及30%[12]。曲霉菌FC-10中純化得到的耐鹽蛋白酶,18% NaCl環境中酶活力能保持50%[28]。由此可以看出,該蛋白酶和已純化得到的耐鹽蛋白酶耐鹽性相近,即使在高鹽度的環境中也保持著較高的酶活。隨著米曲霉基因組測序的完成,米曲霉分子遺傳改造成效卓著,人溶菌酶和牛凝乳酶在米曲霉中異源表達,以及單個堿基的精準敲除或者插入[29]。因此,有望將此胞外耐鹽蛋白酶進行過表達,或者異源表達,應用于醬油工業生產,改善醬油風味[30]。
3 結 論
鈣蛋白酶RIM13首次從米曲霉3.042中分離得到,高鹽環境中保持較高的酶活力。該蛋白酶的最適溫度50 ℃;穩定溫度40 ℃;最適pH 6.5;穩定pH 7.0。Mn2+促進蛋白酶活,Fe3+、Fe2+、Cu2+、Ca2+、K+、Na+抑制蛋白酶活力,以上金屬離子對蛋白酶的二級結構也產生不同程度的影響;米氏常數Km為2.43 g/L,最大反應速率Vm為103.09 mg/(L·min)。在5、10 g/100 mL和15 g/100 mL的NaCl質量濃度下,保留的酶活力分別為77.22%、54.39%以及41.15%。該蛋白酶的酶學性質也表明,其適合應用于醬油醬醪發酵過程,在醬油工業發酵中具有很大的潛在應用價值。
