絲/蘇氨酸蛋白激酶DNA 甲基化在同型半胱氨酸致血管內(nèi)皮細(xì)胞凋亡中的作用
2019-04-02 07:43:32車(chē)雙雙徐華郭偉孫磊張鳴號(hào)楊曉玲姜怡鄧劉志宏
實(shí)用醫(yī)學(xué)雜志 2019年5期
關(guān)鍵詞:檢測(cè)
車(chē)雙雙 徐華 郭偉 孫磊 張鳴號(hào) 楊曉玲 姜怡鄧 劉志宏
寧夏醫(yī)科大學(xué)1公共衛(wèi)生與管理學(xué)院,3基礎(chǔ)醫(yī)學(xué)院(銀川750004);2寧夏血管損傷與修復(fù)研究重點(diǎn)實(shí)驗(yàn)室(銀川750004)
絲/蘇氨酸蛋白激酶(mammalian sterile 20?like kinase 1,MST1)是酵母Ste20 的哺乳動(dòng)物同源物,具有高度的保守性。研究表明,MST1 可以在多種促凋亡刺激如十字孢堿、紫外線、血清饑餓以及抗腫瘤的藥物等情況下激活,引起下游信號(hào)分子活化,進(jìn)而導(dǎo)致細(xì)胞凋亡[1],但有關(guān)其在人臍靜脈血管內(nèi)皮細(xì)胞(HUVECs)中的功能研究報(bào)道甚少。同型半胱氨酸(homocysteine,Hcy)是甲硫氨酸代謝的中間產(chǎn)物,在體內(nèi)參與一碳單位的轉(zhuǎn)移。循證醫(yī)學(xué)證據(jù)顯示高同型半胱氨酸血癥是動(dòng)脈粥樣硬化(atherosclerosis,As)的獨(dú)立危險(xiǎn)因子[2]。近年來(lái),基礎(chǔ)研究證實(shí)Hcy 可以損傷血管HUVECs 功能,加速動(dòng)脈粥樣硬化的發(fā)生和發(fā)展[3]。Hcy 通過(guò)甲硫氨酸循環(huán)影響基因DNA 甲基化水平參與疾病的發(fā)生,并已有文獻(xiàn)報(bào)道DNA 甲基化在Hcy 引起As 中起到重要作用[4]。但其是否影響MST1 的啟動(dòng)子區(qū)甲基化水平,進(jìn)而影響其表達(dá)變化目前仍不清楚。因此,本研究主要探討MST1 在Hcy 致HUVECs 損傷中的作用及其啟動(dòng)子區(qū)DNA 甲基化的調(diào)控機(jī)制,為防治As 提供全新的視角及實(shí)驗(yàn)依據(jù)。
1 材料與方法
1.1 主要實(shí)驗(yàn)儀器和試劑 人臍靜脈內(nèi)皮細(xì)胞株(HUVECs,中國(guó)科學(xué)院細(xì)胞生物學(xué)研究所,中國(guó));超凈工作臺(tái)(蘇州安泰,中國(guó));CO2培養(yǎng)箱(Heraeus,德國(guó));實(shí)時(shí)熒光定量PCR 儀(Bio?Rad,美國(guó));電泳儀(Bio?Rad,美國(guó));凝膠成像儀(Bio?Rad,美國(guó));RPMI1640 培養(yǎng)基、胎牛血清(Gibco,美國(guó));青鏈霉素(碧云天生物技術(shù)研究所,中國(guó));Hcy(Sigma,美國(guó));Annexin V?FITC/PI 細(xì)胞凋亡檢測(cè)試劑盒(貝博公司,中國(guó));總RNA 提取試劑盒(北京天根生物技術(shù)有限公司,中國(guó));DNA 甲基化修飾試劑盒(ZYMORESEARCH,美國(guó));逆轉(zhuǎn)錄和qRT?PCR 試劑盒(Thermo 公司,美國(guó));蛋白提取試劑盒、蛋白定量試劑盒(南京凱基有限公司,中國(guó));MST1 兔抗人、鼠一抗(Abcam 公司,美國(guó)),辣根過(guò)氧化物酶(HRP)標(biāo)記的羊抗兔二抗(北京博奧森生物技術(shù)有限公司,中國(guó));引物由上海生工生物工程有限公司合成。
1.2 方法
1.2.1 細(xì)胞分組及處理 用含10%胎牛血清及1%雙抗的RPMI1640 培養(yǎng)液,于37 ℃、含5%CO2的培養(yǎng)箱中培養(yǎng)HUVECs。當(dāng)細(xì)胞密度達(dá)到85%左右時(shí),將細(xì)胞分為正常對(duì)照組(Control)組、Hcy 組(在培養(yǎng)液中加入Hcy,使其終濃度為100 μmol/L),每組為3 瓶,48 h 后收集細(xì)胞,用于后續(xù)實(shí)驗(yàn)。
1.2.2 MTT 法檢測(cè)HUVECs 的活性 取對(duì)數(shù)生長(zhǎng)期的細(xì)胞,制成1×105個(gè)/mL 細(xì)胞的懸液接種于96 孔板中,培養(yǎng)24 h 后,各組分別加入相應(yīng)藥物孵育48 h,加入MTT繼續(xù)培養(yǎng)4 h,吸棄液體,每孔加入150 μL DMSO 微量震蕩10 min,于酶標(biāo)儀上490 nm測(cè)定各孔OD值,通過(guò)與Control 組比較求出存活率。存活率(%)=實(shí)驗(yàn)組吸光度(A)值/Control 組吸光度(A)值×100%。
1.2.3 Hoechst染色法檢測(cè)細(xì)胞凋亡 生長(zhǎng)融合后,按實(shí)驗(yàn)分組并每組設(shè)3 個(gè)復(fù)孔,繼續(xù)培養(yǎng)48 h;吸盡培養(yǎng)液,用PBS 洗3 次,每次5 min,加入250 μL預(yù)冷的4%多聚甲醛室溫固定細(xì)胞20 min;用PBS洗滌3 次,每次5 min,吸盡液體;加入Hoechst 33258 染色液(2 μg/mL),室溫避光孵育5 min;去染色液,PBS 洗3 次,每次5 min,在激光共聚焦顯微鏡下觀察細(xì)胞核形態(tài)改變。
1.2.4 Western blot 檢測(cè)MST1、DNMT1、Bcl?2 及BAX 蛋白表達(dá) 按照試劑盒說(shuō)明書(shū)提取HUVECs的總蛋白,各組樣本分別取30 μg 總蛋白,經(jīng)SDS?PAGE 電泳、轉(zhuǎn)膜、封閉2 h 后,與MST1、DNMT1、Bcl?2 及BAX 抗體均勻混合后4 ℃孵育過(guò)夜。PBST洗滌3 次,與辣根過(guò)氧化物酶標(biāo)記的羊抗兔或鼠IgG 室溫下孵育2 h,加入ECL 顯色底物,凝膠成像分析儀上成像。設(shè)β?actin 為內(nèi)參,計(jì)算目的基因與β?actin 灰度值的比值,并進(jìn)行分析。
1.2.5 巢式降落式甲基化特異性PCR(nMS?PCR)檢測(cè)MST1 啟動(dòng)子區(qū)DNA 甲基化 按DNA提取試劑盒說(shuō)明書(shū)提取各組細(xì)胞全基因組DNA,檢測(cè)OD260/OD280 值,分析全基因組DNA 樣品純度和濃度。亞硫酸鹽修飾法對(duì)全基因組DNA 進(jìn)行甲基化修飾。nMS?PCR 法檢測(cè)MST1 啟動(dòng)子區(qū)DNA 甲基化改變。針對(duì)MST1 啟動(dòng)子區(qū),設(shè)計(jì)外引物及兩對(duì)內(nèi)引物:外引物:上游5′?AAAAATCTAC?TCATCAAATAAATAACTAAA?3′,下游5′?TTGTTG?GATAGT ATAAAGGTAGAA?3′;甲基化引物:上游5′?CGTTTGGGAGTTTAAGGACG?3′,下游5′?TAAA?CCTAACGAATTCCGACT?3′;非甲基化引物:上游5′?TGTTTGGGAGTTTAAGGATGG?3′,下游5′?CTC?TAAACCTAACAAATTCCAACT?3′。擴(kuò)增反應(yīng)條件:95 ℃5 min,95 ℃30 s,65 ℃30 s,72 ℃30 s,20 個(gè)循環(huán),每個(gè)循環(huán)降0.5~56 ℃,72 ℃7 min。以外引物的PCR 產(chǎn)物為模板,進(jìn)行內(nèi)引物的擴(kuò)增,反應(yīng)條件同外引物。取5 μL PCR 產(chǎn)物于2%的瓊脂糖凝膠上電泳,用凝膠成像分析儀成像并分析甲基化條帶及非甲基化條帶的光密度,按如下公式進(jìn)行結(jié)果的計(jì)算:非甲基化(%)=非甲基化(OD值)/[甲基化(OD值)+非甲基化(OD值)],每組實(shí)驗(yàn)重復(fù)3 次。
1.3 統(tǒng)計(jì)學(xué)方法 采用SPSS 20.0 統(tǒng)計(jì)軟件進(jìn)行統(tǒng)計(jì)學(xué)分析,計(jì)量資料以均數(shù)±標(biāo)準(zhǔn)差表示,兩樣本均數(shù)間比較采用兩獨(dú)立樣本t檢驗(yàn),兩個(gè)變量相關(guān)性分析采用Pearson 相關(guān)系數(shù)分析,P<0.05 表示差異具有統(tǒng)計(jì)學(xué)意義。
2 結(jié)果
2.1 Hcy 對(duì)HUVECs 存活率的影響 100 μmol/L Hcy 干預(yù)細(xì)胞48 h 后,MTT 法檢測(cè)各組細(xì)胞存活率。結(jié)果顯示:與Control 組比較,Hcy 組細(xì)胞增殖受到明顯抑制,其細(xì)胞存活率明顯降低,差異具有統(tǒng)計(jì)學(xué)意義(P<0.01),結(jié)果提示Hcy 對(duì)HUVECs生長(zhǎng)有抑制作用,見(jiàn)圖1。

圖1 Hcy 對(duì)人臍靜脈內(nèi)皮細(xì)胞存活率的影響Fig.1 Effect of Hcy on survival rate of HUVECs
2.2 Hoechst 33258 染色觀察HUVECs 核形態(tài)改變 細(xì)胞經(jīng)DNA熒光染料Hoechst 33258染色5 min后,于共聚焦顯微鏡下觀察細(xì)胞核形態(tài)改變。結(jié)果發(fā)現(xiàn):Control 組細(xì)胞核大小均一、核圓且規(guī)整,核膜光滑無(wú)皺縮;Hcy 組細(xì)胞核與細(xì)胞質(zhì)邊界模糊,胞核固縮,大小不一,呈現(xiàn)不規(guī)則形狀改變,見(jiàn)圖2。
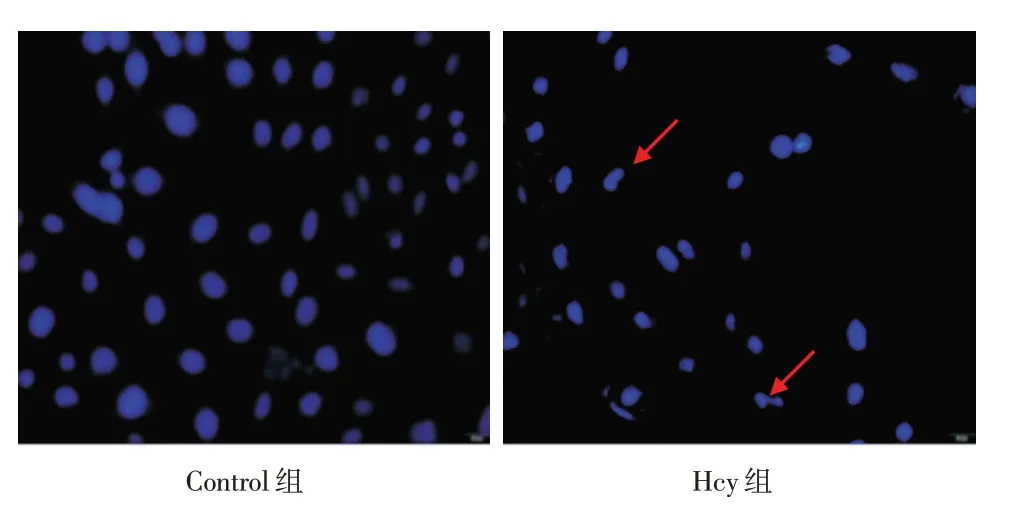
圖2 各組核形態(tài)通過(guò)Hoechst 33258 染色檢測(cè)(×400)Fig.2 Nuclear morphology stained by Hoechst 33258 staining in each group
2.3 Hcy 對(duì)HUVECs 中Bcl?2 和BAX 表達(dá)的影響 為了進(jìn)一步明確Hcy 在HUVECs 凋亡中的作用,Western blot 檢測(cè)了各組細(xì)胞中Bcl?2 和BAX 的蛋白表達(dá)水平(圖3A)。與Control 組相比,Hcy 刺激后,Bcl?2 的蛋白表達(dá)降低了11.04%,差異有統(tǒng)計(jì)學(xué)意義(P<0.05),BAX 的蛋白表達(dá)增加了1.23倍,差異具有統(tǒng)計(jì)學(xué)意義(P<0.05),Bcl?2 與BAX的比率降低,差異具有統(tǒng)計(jì)學(xué)意義(P<0.05),提示Hcy 可能介導(dǎo)HUVECs 凋亡(圖3B)。
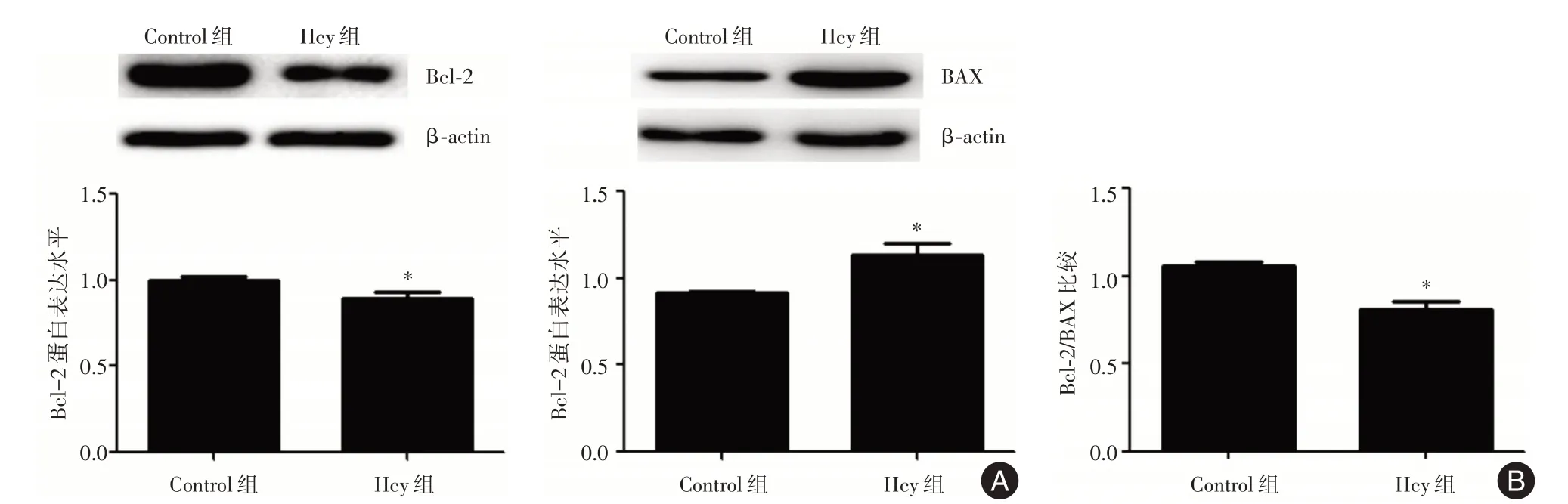
圖3 HUVECs 中Bcl?2 和BAX 的蛋白表達(dá)Fig.3 The protein expression of Bcl?2 and BAX in HUVECs
2.4 Hcy 干預(yù)HUVECs 后MST1 的蛋白表達(dá) 為了探討MST1 在Hcy 介導(dǎo)HUVECs 凋亡中的作用,Hcy 干預(yù)細(xì)胞后檢測(cè)MST1 的表達(dá)。結(jié)果顯示,與Control 組比較,Hcy 組MST1 蛋白表達(dá)水平為Con?trol 組的1.13 倍(P<0.01),表明MST1 的表達(dá)上調(diào)可能在高Hcy 介導(dǎo)HUVECs 凋亡中發(fā)揮著重要的作用,見(jiàn)圖4。
2.5 HUVECs 中MST1 表達(dá)與Bcl?2/BAX 的相關(guān)性分析 為進(jìn)一步驗(yàn)證MST1 與HUVECs 凋亡的相關(guān)性,將MST1 蛋白表達(dá)水平與Bcl?2/BAX 進(jìn)行了相關(guān)性分析,結(jié)果顯示:細(xì)胞MST1 表達(dá)水平與Bcl?2/BAX 呈負(fù)相關(guān)(r2=0.844 6,P<0.001),提示MST1 與HUVECs 凋亡密切相關(guān),見(jiàn)圖5。

圖4 HUVECs 中Hcy 對(duì)MST1 蛋白表達(dá)的影響Fig.4 The effect of Hcy on the protein expression of Bcl?2 and BAX in HUVECs

圖5 MST1 水平與Bcl?2/BAX 比值的相關(guān)性分析Fig.5 Analysis of correlation between MST1 protein levels and the ratio of Bcl?2/BAX
2.6 MST1 生物信息學(xué)分析 利用生物信息學(xué)軟件MethPrimer 分析MST1 啟動(dòng) 子 區(qū)CpG 島,選取MST1 基因上游2 500 bp 作為研究對(duì)象,發(fā)現(xiàn)MST1啟動(dòng)子區(qū)含有CPG島,分別位于?1516/?1459、?1368/?871、?850/?682,提示MST1 基因很可能受到甲基化的調(diào)控,見(jiàn)圖6。
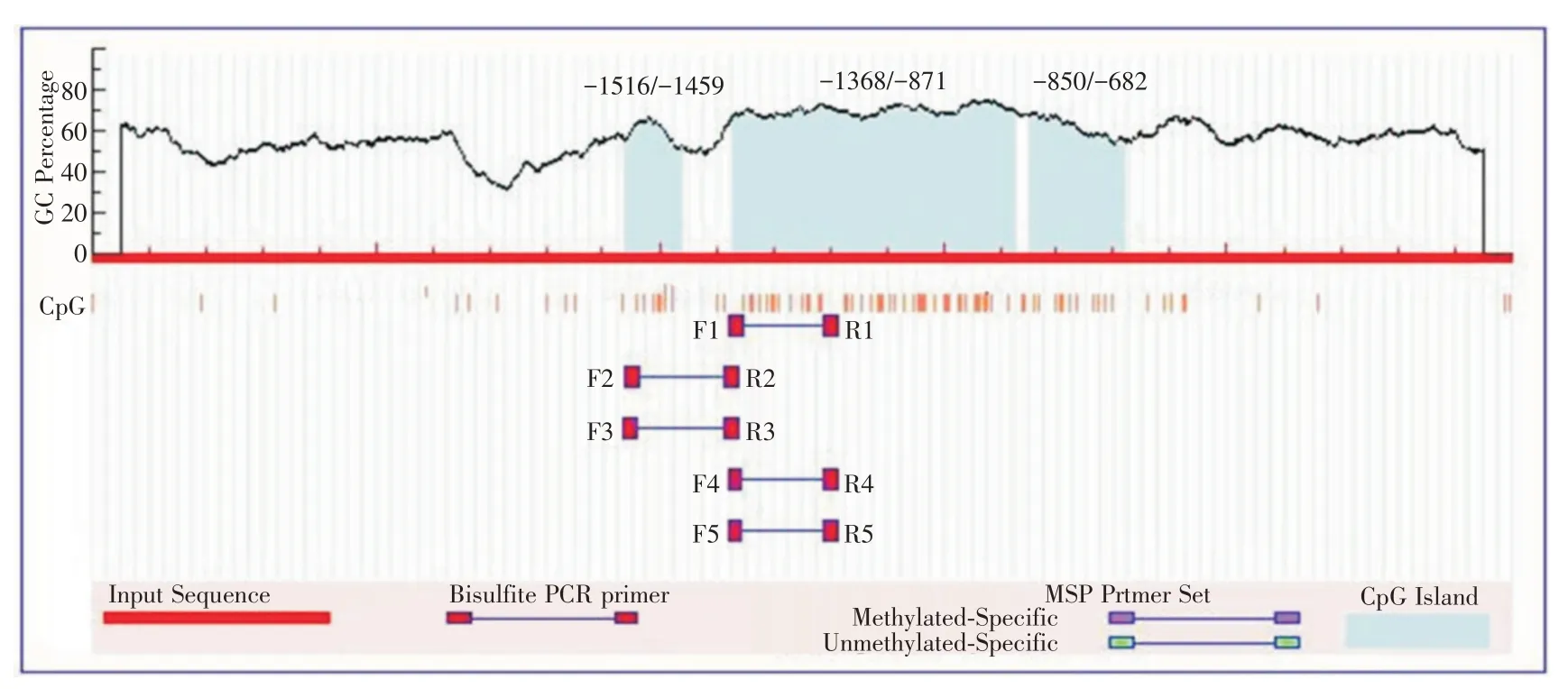
圖6 MST1 啟動(dòng)子區(qū)的生物信息學(xué)分析Fig.6 Bioformatics analysis of MST1 promoter region
2.7 Hcy 對(duì)HUVECs 中MST1 啟 動(dòng) 子 區(qū)DNA 甲基化的影響 為探討Hcy 致HUVECs 凋亡中調(diào)控MST1 表達(dá)的機(jī)制,Hcy 干預(yù)HUVECs 后nMS?PCR法檢測(cè)MST1 啟動(dòng)子區(qū)DNA 甲基化水平。結(jié)果顯示:與Control 組相比,Hcy 組MST1 DNA 甲基化水平明顯降低18.39%,差異具有統(tǒng)計(jì)學(xué)意義(P<0.01),提示DNA 甲基化參與了MST1 的表達(dá)調(diào)控,見(jiàn)圖7。
2.8 Hcy 干預(yù)后DNMT1 蛋白表達(dá)的改變 100 μmol/L Hcy 處理HUVECs 48 h 后,Western blot 檢測(cè)了Control 組和Hcy 組中DNMT1 的蛋白表達(dá)。結(jié)果顯示:Hcy 干預(yù)后,DNMT1 蛋白表達(dá)顯著降低,差異具有統(tǒng)計(jì)學(xué)意義(P<0.01),提示DNMT1 可能在MST1 的表達(dá)調(diào)控中發(fā)揮重要作用,見(jiàn)圖8。
3 討論
動(dòng)脈粥樣硬化(As)是心腦血管疾病共同的病理基礎(chǔ),嚴(yán)重危害人類健康[5]。Hcy 是As 的獨(dú)立危險(xiǎn)因子。研究報(bào)道,Hcy 致As 的機(jī)制涉及內(nèi)皮細(xì)胞損傷、VSMCs 增殖以及泡沫細(xì)胞的形成[6]。內(nèi)皮細(xì)胞損傷是As 形成的早期始動(dòng)環(huán)節(jié),同時(shí)也是發(fā)生As 的必備條件。引起內(nèi)皮功能損傷的因素眾多,其中內(nèi)皮細(xì)胞凋亡是重要因素之一[7]。因此,深入研究其功能的改變及機(jī)制,具有潛在的臨床應(yīng)用前景。
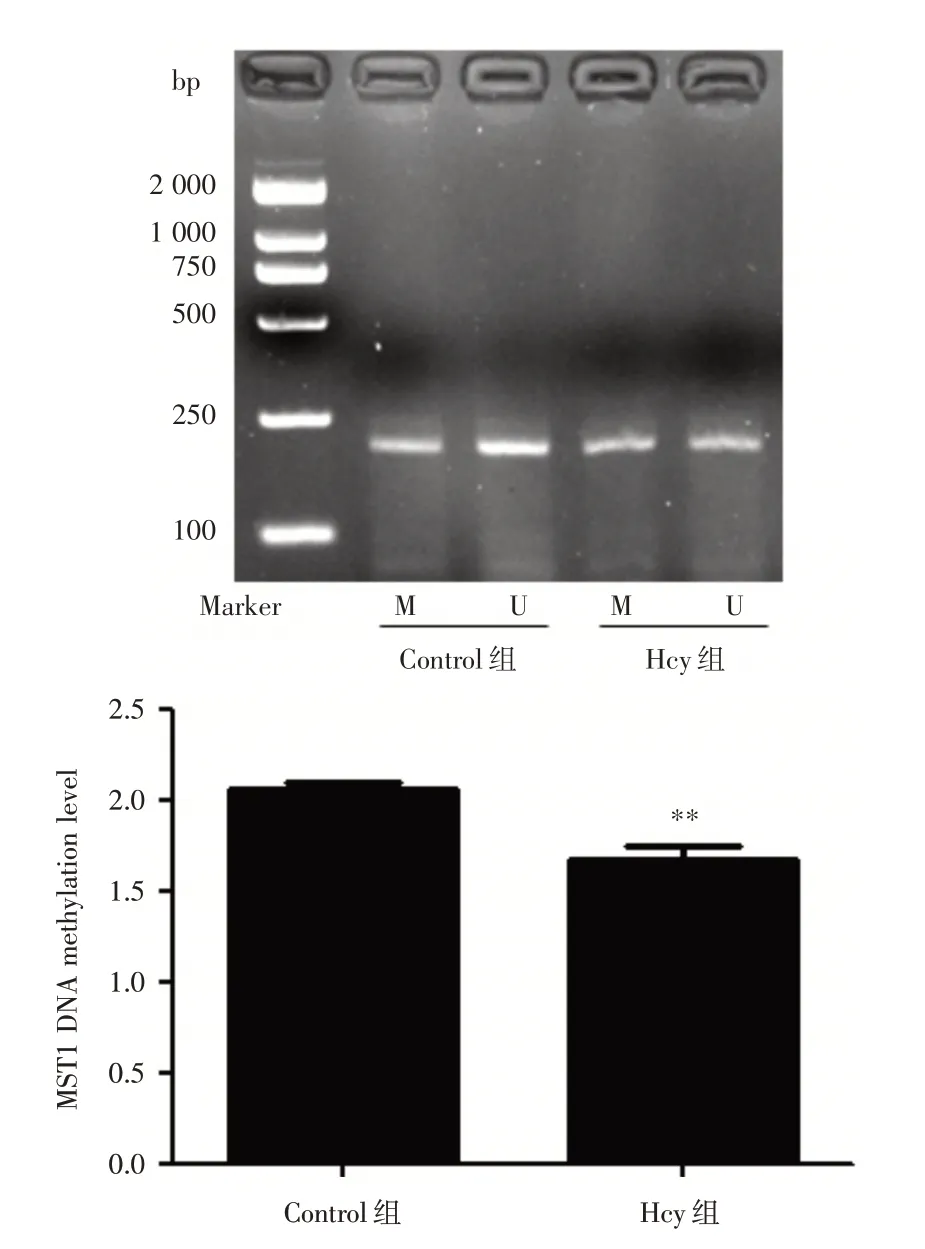
圖7 在HUVECs 中Hcy 誘導(dǎo)MST1 甲基化水平改變Fig.7 Hcy induced change of MST1 DNA methylation level in HUVECs
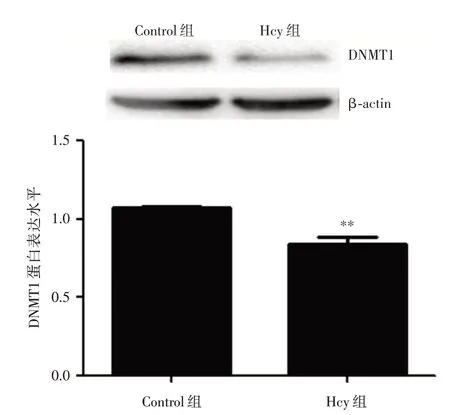
圖8 HUVECs 中Hcy 對(duì)DNMT1 表達(dá)的影響Fig.8 The effect of Hcy on the expression of DNMT1 in HUVECs
本文在前期研究的基礎(chǔ)上,篩選出100 μmol/L Hcy 的濃度對(duì)內(nèi)皮細(xì)胞增殖抑制率明顯升高[8],采用該濃度Hcy干預(yù)HUVECs 建立As體外實(shí)驗(yàn)?zāi)P停杂^察Hcy 對(duì)HUVECs 凋亡的影響,并從內(nèi)皮細(xì)胞凋亡的角度探討Hcy 在As 中作用。實(shí)驗(yàn)中可觀察到Hcy 能夠使HUVECs 存活率降低,對(duì)HUVECs 損傷具有促進(jìn)作用。采用Hoechst 33258 細(xì)胞核熒光染色和Western blot 技術(shù)進(jìn)一步觀察細(xì)胞的凋亡情況,結(jié)果發(fā)現(xiàn):Control 組細(xì)胞核呈微弱均勻藍(lán)色熒光;Hcy 組細(xì)胞核顯示較深藍(lán)色熒光狀,可見(jiàn)核固縮、凋亡小體等典型的凋亡形態(tài)學(xué)特征。細(xì)胞凋亡是一系列基因活動(dòng)引起的級(jí)聯(lián)反應(yīng)的結(jié)果,由誘導(dǎo)基因和抑制基因共同調(diào)控。Bcl?2 具有抗細(xì)胞凋亡的功能,它能通過(guò)抵抗多種形式的細(xì)胞凋亡延長(zhǎng)細(xì)胞壽命。而B(niǎo)AX是Bcl?2的同源基因,其表達(dá)上調(diào)可拮抗Bcl?2 的保護(hù)效應(yīng)使細(xì)胞趨于凋亡。細(xì)胞凋亡是否發(fā)生是由Bcl?2 和BAX 決定的[9]。Western blot 結(jié)果顯示,與Control 組比較,加入Hcy后,Bcl?2 的蛋白表達(dá)顯著降低,而Hcy 組中BAX的蛋白表達(dá)明顯增高,以上結(jié)果提示:Hcy 能夠誘導(dǎo)HUVECs 的凋亡。
MST1 是體內(nèi)普遍表達(dá)的一種絲/蘇氨酸蛋白激酶,主要參與細(xì)胞生長(zhǎng)發(fā)育、細(xì)胞遷移和分化、增殖、凋亡以及器官大小等的調(diào)控[10]。有研究[11]表明,多種促凋亡刺激和細(xì)胞應(yīng)激如紫外線照射、TNF?α 及抗腫瘤藥物可激活MST1,引起下游信號(hào)分子活化,進(jìn)一步導(dǎo)致細(xì)胞凋亡。張輝等[12]研究發(fā)現(xiàn),MST1 基因在腎癌組織中表達(dá)明顯下調(diào),過(guò)表達(dá)MST1 后,可使腎癌凋亡明顯升高;有學(xué)者的研究也發(fā)現(xiàn),在穩(wěn)定轉(zhuǎn)染MST1 的HeLa 細(xì)胞中,可以加速caspase?3 的激活使得細(xì)胞對(duì)死亡受體介導(dǎo)的凋亡更敏感[2,13]。本研究結(jié)果發(fā)現(xiàn),與Control 組比較,Hcy 組細(xì)胞中MST1 蛋白表達(dá)明顯升高,與HUVECs 中Bcl?2 蛋白水平呈負(fù)相關(guān),而與BAX 蛋白水平呈正相關(guān),提示MST1 可能是促進(jìn)HUVECs凋亡的重要調(diào)節(jié)因子。
Hcy 是一種含硫的非必需氨基酸,其作為甲硫氨酸代謝的中間產(chǎn)物,參與體內(nèi)甲硫氨酸循環(huán)的關(guān)鍵環(huán)節(jié),通過(guò)將甲基轉(zhuǎn)移至DNA、蛋白質(zhì)等物質(zhì)發(fā)揮生物學(xué)效應(yīng)[14]。DNA 甲基化是一種重要的表觀遺傳學(xué)修飾,以未改變核苷酸順序及其組成的方式,影響DNA 構(gòu)象穩(wěn)定性,從而調(diào)控基因表達(dá)。一般情況下,DNA 甲基化誘導(dǎo)基因的表達(dá)沉默,而去甲基化則激活基因的表達(dá)[15]。前期研究發(fā)現(xiàn),在As 中Hcy 參與多種基因甲基化修飾狀態(tài)的改變。生物信息學(xué)分析發(fā)現(xiàn)MST1 啟動(dòng)子區(qū)富含CpG 位點(diǎn),因此筆者推測(cè),DNA 甲基化這一表觀遺傳學(xué)機(jī)制參與了Hcy 調(diào)控MST1 的表達(dá)進(jìn)而促進(jìn)HUVECs 凋亡,為了驗(yàn)證這一假設(shè),本課題組進(jìn)一步檢測(cè)Hcy 干預(yù)后MST1 啟動(dòng)子區(qū)甲基化程度的改變。結(jié)果顯示,Hcy 組細(xì)胞中MST1 啟動(dòng)子區(qū)DNA 甲基化程度明顯降低,DNA 低甲基化作為基因轉(zhuǎn)錄增強(qiáng)的重要標(biāo)志之一,與MST1 表達(dá)水平上調(diào)相一致。同時(shí),本研究發(fā)現(xiàn)Hcy 干預(yù)后HUVECs中DNMT1 表達(dá)降低,其主要原因是DNA 甲基化主要是通過(guò)DNA 甲基轉(zhuǎn)移酶家族(DNMTs)催化發(fā)生并維持的,而DNMT1 是哺乳動(dòng)物體內(nèi)含量最多的一種DNMTs[3,16],其表達(dá)降低,會(huì)導(dǎo)致DNA 低甲基化、基因的表達(dá)激活,從而誘導(dǎo)細(xì)胞的凋亡改變。
綜上所述,本研究結(jié)果表明,Hcy 通過(guò)下調(diào)DNMT1 表達(dá)導(dǎo)致MST1 基因啟動(dòng)子區(qū)DNA 甲基化水平降低,進(jìn)而引起MST1 的表達(dá)升高,促進(jìn)HU?VECs 凋亡。因此,繼續(xù)對(duì)MST1 在Hcy 誘導(dǎo)的HU?VECs 凋亡調(diào)控機(jī)制中的深入研究,將有望為防治As 找到新的靶點(diǎn)和方向。
猜你喜歡
中國(guó)設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
