運動生理學(xué)的學(xué)科起源
——基于骨骼肌相關(guān)研究的追溯*
2019-07-24 01:27:46蘇利強張克峰溫岱宗李加鵬陳海春
天津體育學(xué)院學(xué)報 2019年4期
蘇利強,張克峰,2,溫岱宗,李加鵬,陳海春
體育學(xué)科追根溯源是體育學(xué)研究關(guān)注的領(lǐng)域之一。目前,較普遍認(rèn)為運動訓(xùn)練學(xué)是體育的本源學(xué)科,其他眾多學(xué)科是隨著科學(xué)進步和學(xué)科分類的交叉重疊而融入體育領(lǐng)域,并成為體育學(xué)的重要支撐學(xué)科,如運動生理學(xué)、運動解剖學(xué)等基礎(chǔ)課程均是從生理學(xué)、解剖學(xué)分支出來的,母學(xué)科知識融入體育領(lǐng)域的過程也是體育學(xué)科豐富發(fā)展的歷程。隨著現(xiàn)代社會的發(fā)展,體育對綜合性知識需求越來越大,體醫(yī)融合的落實需要體育學(xué)與臨床醫(yī)學(xué)的知識做支撐,體育運動項目社會化需要體育學(xué)與社會學(xué)知識的交叉。這些問題逼著我們“走出體育看體育”,從更廣闊的視角來解決體育科學(xué)發(fā)展面臨的問題。“述往事,思來者”,歷史是一本教科書,是現(xiàn)實的指南。追溯母學(xué)科知識融入體育學(xué)的過程,講述運動生理學(xué)誕生前期的科研故事,探尋知識產(chǎn)生、積累與融合之初衷,對正確理解體育學(xué)的學(xué)科本位有指導(dǎo)意義,對母學(xué)科知識與體育學(xué)的現(xiàn)代交融有啟示作用,對解決新時期體育學(xué)遇到的新問題具有借鑒意義。
梳理運動生理學(xué)的起源有必要對生理學(xué)進行追溯。生理學(xué)成為一門科學(xué)是在1628年,威廉·哈維出版的一本An anatomical study of the motion of the heart and of the blood in animals,翻譯為《心血運動論》[1-2](見圖1),書中的血液循環(huán)學(xué)說被恩格斯評價為這是生理學(xué)成為科學(xué)的標(biāo)志,18世紀(jì)以后生理學(xué)在神經(jīng)、消化、呼吸以及內(nèi)分泌諸方面獲得了很大的進步,這些成果標(biāo)志著生理學(xué)生根發(fā)芽[3-5]。生理學(xué)知識的豐富和發(fā)展是運動生理學(xué)起源的基礎(chǔ)。

圖1心血運動論及不同版本的譯著[1-2] Figure1 The Motion of Heart and Blood and Different Versions of Translated Works(Cited From William Harvey
國內(nèi)對運動生理學(xué)發(fā)展史的資料較少,雖記載了第1本運動生理學(xué)教材是1889年的《身體運動的生理學(xué)》,但對該教材之前的研究提及甚少[6-9],楊錫讓[10]、王瑞元[11]曾綜述了運動生理學(xué)現(xiàn)狀與展望,未提及19世紀(jì)以前生理學(xué)中有關(guān)運動的研究。作為一門完整的學(xué)科,只有其內(nèi)部知識體系完整統(tǒng)一、新舊知識演化清晰,才能明確學(xué)科發(fā)展性特征[12-13]。追溯早期骨骼肌及運動的研究成果,探索這些知識與運動生理學(xué)知識體系的邏輯關(guān)系,不僅對追溯運動生理學(xué)學(xué)科起源有歷史意義,也對確立運動生理學(xué)的學(xué)科獨立性有重要意義。
本研究運用文獻資料法和文本分析法進行研究,檢索數(shù)據(jù)庫包括PubMed、MEDLINE、Nature及子刊,檢索詞為Muscle,檢索語言為英語,檢索時間為1889年12月31日之前,納入標(biāo)準(zhǔn):骨骼肌形態(tài)結(jié)構(gòu)、骨骼肌收縮、骨骼肌神經(jīng)支配、隨意運動、疲勞;排除標(biāo)準(zhǔn):疾病治療、與本研究主題無關(guān);共檢索764篇文獻,經(jīng)過篩查102篇符合要求,其中43篇文獻納入本課題重點研究,通過總結(jié)、歸納上述研究成果,結(jié)合骨骼肌研究成果與學(xué)科知識體系框架,從以下幾個方面闡述運動生理學(xué)的起源。
1 骨骼肌形態(tài)結(jié)構(gòu)和功能的研究
對骨骼肌形態(tài)結(jié)構(gòu)的研究經(jīng)歷了從宏觀到微觀不斷深入的過程。19世紀(jì)初期對肌肉的認(rèn)識是通過解剖尸體觀察肌肉形態(tài)結(jié)構(gòu)[14-16],只到顯微鏡的發(fā)明和使用后,人才逐漸認(rèn)識到骨骼肌的微細(xì)結(jié)構(gòu),1866年有學(xué)者對骨骼肌光學(xué)顯微鏡下的微細(xì)結(jié)構(gòu)進行了描述[17],但骨骼肌微細(xì)結(jié)構(gòu)界限不太清晰,從顯微鏡下的視覺效果出發(fā),W.ABBEY[18]在1867年在普通骨骼肌切片的基礎(chǔ)上,采用了洋紅(Carmine)對骨骼肌細(xì)胞進行了染色,從而將細(xì)胞核和細(xì)胞質(zhì)與相鄰結(jié)構(gòu)通過不同顏色區(qū)分開來,使觀察到的骨骼肌微細(xì)結(jié)構(gòu)更加清晰。隨著對骨骼肌微細(xì)結(jié)構(gòu)的深入研究,1879年在檢查冷凍青蛙的完全伸展肌肉的一些纖維時,顯微鏡下發(fā)現(xiàn)肌肉溫度只要低于30℃,橫向條紋是消失,但溫度略高于30℃,橫向暗條開始出現(xiàn)。同時發(fā)現(xiàn)縱向減少與肌肉橫向測量的增加有關(guān),沒有任何體積損失,在靜息狀態(tài)下沒有縱向條紋,并且在肌肉收縮期間出現(xiàn)[19],這一成果是基于嚴(yán)密的實驗設(shè)計,探索了不同溫度對肌肉微細(xì)結(jié)構(gòu)的影響,從而發(fā)現(xiàn)了肌纖維的明帶暗帶,為骨骼肌收縮的肌絲滑行學(xué)說奠定了基礎(chǔ)。
19世紀(jì),骨骼肌運動功能的研究是基于醫(yī)學(xué)的需求,隨著對骨骼肌相關(guān)疾患的探索與治療,使人們對骨骼肌運動功能的認(rèn)識有了長足的進步。1871年在肌肉關(guān)節(jié)疼痛的相關(guān)研究中對腓腸肌的起點進行分析,描述了腓腸肌的屈膝功能,并指出了能輕微地幫助脛骨旋轉(zhuǎn)[20];1872年對胸大肌的起點與止點的位置進行了觀察,分析胸大肌和周圍其他肌肉的位置關(guān)系,說明了胸大肌對肩關(guān)節(jié)的功能[21];1873年對腹肌的分布與結(jié)構(gòu)研究中發(fā)現(xiàn)腹肌的后腹部是從深層而來的,前腹是從淺層來的,中間肌腱是原始未分層肌肉橫隔的殘余[22];1874年,在研究肌肉功能對骨骼發(fā)育的影響時,以沒有股方肌的病例為對象,發(fā)現(xiàn)其第六腰椎的脊柱上第一肋骨不發(fā)育[23],肌肉的輔助結(jié)構(gòu)病變影響肌肉的功能,因此對肌肉和肌腱的細(xì)微解剖結(jié)構(gòu)進行了研究[24]。從早期對骨骼肌運動功能的描述看,為了解決肌肉相關(guān)疼痛疾病,學(xué)者在骨骼肌結(jié)構(gòu)的基礎(chǔ)上對其機能進行了研究,這些研究不僅有助于治療骨骼肌疼痛的疾病,同時對骨骼肌運動功能的認(rèn)識也有較大的促進。可見,人們對知識探求的動力源自于對生活實踐的需求,現(xiàn)階段運動生理學(xué)的研究也應(yīng)該基于運動實踐,解決現(xiàn)實生活中人們切實的運動需求問題。
2 生物電與骨骼肌收縮
上述研究從宏觀到微細(xì)觀察層面分析了骨骼肌結(jié)構(gòu)與功能,但形態(tài)學(xué)解決不了骨骼肌收縮的動態(tài)機制,生物電檢測技術(shù)的發(fā)展使人們對骨骼肌收縮的認(rèn)識向前推進了一大步。1786年L.GALVANI(1737—1798)及其研究團隊用帶有靜電的金屬棒觸碰了青蛙腿的神經(jīng)并觀察到肌肉收縮,發(fā)現(xiàn)肌肉收縮與生物電有關(guān)。肌肉收縮程度與生物電的具體關(guān)系未知,1867年,學(xué)者觀察死亡后肌肉的生物電,結(jié)果發(fā)現(xiàn)發(fā)現(xiàn)死亡后的肌肉電流減少與肌肉興奮性的減弱成正比,肌肉的電興奮收縮產(chǎn)生的力量(electro-motor force)和興奮性同時終止,認(rèn)為肌肉收縮性功能離不開肌肉的電興奮性[25]。肌肉收縮是可變電流還是恒定電流引發(fā)的?1868年,當(dāng)W.RUTHERFORD用一個恒定電流通過刺激運動神經(jīng)時,發(fā)現(xiàn)肌肉只在電流開始或結(jié)束時收縮,在電流恒定時肌肉保持靜止?fàn)顟B(tài)(見圖2),肌肉收縮是可變電流引發(fā)的,既然肌肉收縮時和安靜時都存在生物電,那么可變電流引發(fā)的肌肉收縮時的生物電狀態(tài)與安靜時的生物電狀態(tài)什么區(qū)別?1871年,C.B.RADCLIFFE發(fā)現(xiàn),肌肉收縮時電狀態(tài)不是靜止的,而是流動的[26]。研究者將穿過神經(jīng)和肌肉的電流引起一種奇特的狀態(tài)稱為負(fù)電狀態(tài)(electrotonic state)或電緊張狀態(tài),認(rèn)為這種神經(jīng)和肌肉狀態(tài)的特征在于電勢和興奮程度的改變[27]。當(dāng)時有學(xué)者提出神經(jīng)和肌肉的生物電狀態(tài)是由極性分子(peripolar molecules)組成,神經(jīng)和肌肉的電流源自于極性分子,但是這一觀點很多生理現(xiàn)象不能解釋,如神經(jīng)肌肉活動過程中電流的減弱、兩個相反的電流如何區(qū)分。1871年,T.L.BRUNTON等[28]在這一學(xué)說的基礎(chǔ)上對肌肉收縮時電流變化的機制進行了進一步的推理,認(rèn)為肌肉中的電流由不同的原生質(zhì)之間的差異所產(chǎn)生。1876年,為驗證這一學(xué)說,G.J.ROMANES采用神經(jīng)斷裂的模型研究神經(jīng)滲透肌肉的電子條件,結(jié)果發(fā)現(xiàn)肌肉內(nèi)的神經(jīng)終板在電刺激時具有和神經(jīng)干不同的生物學(xué)特性[29]。1878年學(xué)者對所有可興奮的組織進行研究,發(fā)現(xiàn)可興奮組織在刺激作用下作出電-運動反應(yīng)時,原生質(zhì)在興奮部位的電位變?yōu)榱素?fù)值,而沒有興奮的部位保持不變,這對認(rèn)識骨骼肌收縮時的負(fù)電狀態(tài)有了較大的進步[30]。這一時期的研究從生物電與肌肉收縮現(xiàn)象出發(fā),在控制電流變化情況下了解了肌肉收縮是需要可變電流的,為了解釋這一現(xiàn)象,學(xué)者提出了學(xué)說,認(rèn)為電流是由不同原生質(zhì)之間的差異所產(chǎn)生,這一學(xué)說為今后的膜電位和動作電位的離子學(xué)說奠定了基礎(chǔ)。
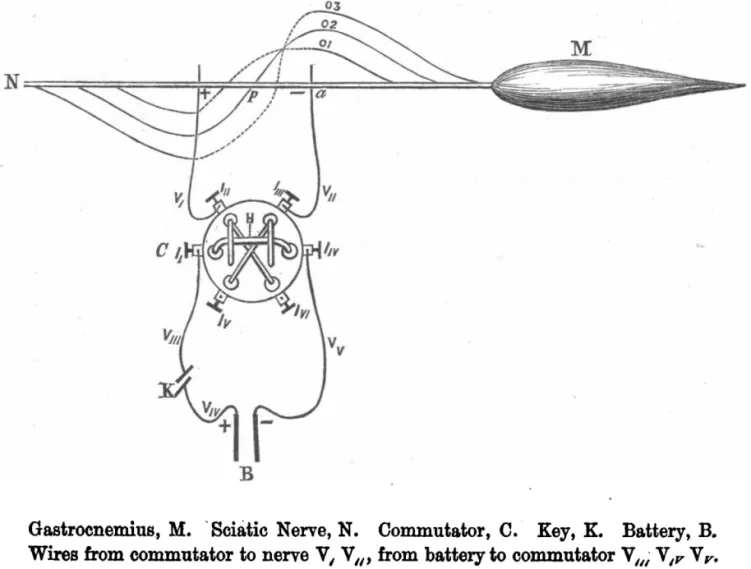
圖2電刺激神經(jīng)監(jiān)測肌肉活動示意圖[26] Figure2 Schematic Diagram of Muscle Activity Monitored by Electrical Stimulation of Nerves[26]
在認(rèn)識可變電流引起骨骼肌收縮的基礎(chǔ)上,學(xué)者開始對不同電流刺激骨骼肌進行了研究。1881年J.T.CASH采用磁電機旋轉(zhuǎn)裝置產(chǎn)生不同電流,通過反復(fù)測試并找到引起肌肉不同負(fù)重的最大收縮時的電流,通過肌肉最大收縮的電流刺激肌肉,記錄肌肉不同負(fù)重的肌肉收縮曲線,結(jié)果發(fā)現(xiàn)同一肌肉不同負(fù)重的肌肉收縮曲線不同,后來采用不同動物進行實驗,發(fā)現(xiàn)鴿子的胸肌、兔子的肌肉以及青蛙的肌肉存在不同的電刺激-收縮特征,這種電刺激-肌肉收縮的特征在不同動物、不同部位均表現(xiàn)不同特征(見圖3)[31]。改變研究中變量條件,獲得不同的實驗結(jié)果,是科學(xué)探索的重要手段。正如J.T.CASH[31]的研究,通過不同電流刺激不同骨骼肌,分析電刺激-肌肉收縮的特征,使人們逐漸認(rèn)識了紅白肌的特征,而紅白肌的形態(tài)機能特征則是當(dāng)時到以后一段時間內(nèi)運動生理學(xué)研究的重要內(nèi)容之一。
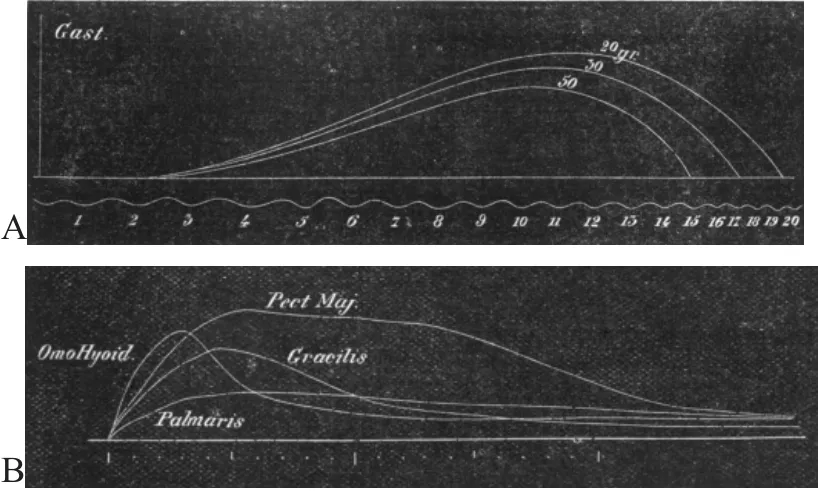

圖3不同負(fù)重不同肌肉大刺激收縮曲線Figure3 Contraction Curves of Different Muscle Stimuli with Different Weights
3 骨骼肌收縮的潛伏期
隨著對電刺激肌肉單收縮研究的不斷推進,在描記肌肉單收縮圖形時發(fā)現(xiàn)從電刺激開始到肌肉收縮總有一定的時間間隔,這一時間間隔是儀器產(chǎn)生的誤差還是客觀存在的?另外神經(jīng)的電信號是如何傳到肌肉的?基于這些問題學(xué)者展開了一系列的研究。1859年,潛伏的肌肉激發(fā)期的持續(xù)時間(當(dāng)時沒有正式命名為潛伏期)和重要性受到質(zhì)疑,當(dāng)時的觀點認(rèn)為運動的有形表達的延遲可能是由于從骨骼肌收縮到描記杠桿傳遞所需的時間,以及描記圖形時使用的書寫筆與紙張的摩擦,在當(dāng)時的測試方法中所有的條件都有可能延遲書寫杠桿和書寫筆的可見運動,不存在肌肉潛在的激發(fā)期的理由是沒有任何一個物體的運動速度與電的速度相當(dāng)。可見當(dāng)時對潛伏期的認(rèn)識處在推斷階段,對假設(shè)的科學(xué)性和合理性有效的質(zhì)疑可以推動科學(xué)的不斷進步,接下來的實驗,為了避免儀器帶來的誤差,對多種檢測肌電的儀器(見圖4~圖6)進行了的不斷改進,測試方法優(yōu)化,確定了電刺激與反應(yīng)的最短時間間隔,圖7是當(dāng)時的儀器和測試出來的“潛伏期”圖。這一階段是對“潛伏期”的質(zhì)疑與解決階段。

圖4 電信號檢測儀示意圖[32] Figure4 Cambridge Electric Signal[32]

圖5 檢測完整肌肉或半塊肌肉的潛伏期裝置示意圖[32] Figure5 Showing the Arrangement of Lever for Comparing Latency of the Whole and Half Muscle[32]

圖6 記錄肌電反映的儀器示意圖[33] Figure6 Records the Instrumental Schematic of EMG[33]

圖7 2種儀器記錄五通道潛伏期電信號特征圖[32] Figure7 Facsimile of Five Tracings of the Two Kinds of Electric Signal Showing its Latency[32]
在確定了潛伏期存在的基礎(chǔ)上,學(xué)者開始對潛伏期的特征進行了研究。1883年G.F.YEO等對蛙腓腸肌通過改變刺激強度、增加肌肉的負(fù)重重量、改變溫度觀察單收縮的特征,并得出了增加肌肉收縮的速度和力量的合適溫度,并對極度疲勞的肌肉進行實驗,發(fā)現(xiàn)疲勞后的肌肉表現(xiàn)為潛伏期變長、收縮時間延長、曲線高度降低等特征[34]。隨著對肌肉單收縮研究的深入,學(xué)者也開始對第1次刺激后的第2次刺激產(chǎn)生興趣,為了檢測肌肉收縮特征,對有學(xué)者對肌動描記器進行了改進(見圖9)。1885年的一項研究采用兩次連續(xù)的誘導(dǎo)電刺激神經(jīng),用肌動描記器記錄肌肉變化,結(jié)果發(fā)現(xiàn)肌肉出現(xiàn)第2次收縮刺激效果的最短間隔是1/600 s,隨后采用肌動描記器(專門針對此目的進行調(diào)整的擺錘形測量儀)進行了進一步的研究,結(jié)果發(fā)現(xiàn)最大刺激的間隔只有0.001 s,并且認(rèn)為獲得最大刺激的最小間隔,第2刺激應(yīng)略強于第1刺激。在此基礎(chǔ)上對肌肉產(chǎn)生疲勞的刺激特征和興奮性特征進行了闡述[35]。

圖8 擺式肌動描記器示意圖[37] Figure8 Description of Figure[37]
上述對神經(jīng)刺激肌肉收縮的研究中基本確定了潛伏期的存在,并記錄了潛伏期的一下規(guī)律,接下來的問題是潛伏期是如何形成的?神經(jīng)的電刺激信號是如何傳遞到肌肉的?一些學(xué)者提出神經(jīng)沖動從神經(jīng)到肌肉收縮的過程可能經(jīng)歷了特定的“中間器官”,這一中間器官可能是潛伏期的形成原因,其中具有代表性的研究是1885年A.W.HOISHOLT等通過直接刺激肌肉來獲得最大肌肉收縮與刺激神經(jīng)干引起肌肉收縮的潛伏期存在很大的差異,推測可能是運動神經(jīng)末梢刺激過程的延遲,在此過程中,釋放的潛在功率將累積,直到其相當(dāng)于肌肉所需的刺激為止才引起肌肉收縮[36]。
這一時期的一系列研究是以問題為導(dǎo)向,以檢測技術(shù)革新為基礎(chǔ),從檢測技術(shù)層面逐步解決了對潛伏期的質(zhì)疑,當(dāng)然了解了潛伏期的存在并不是研究的終點,潛伏期產(chǎn)生的機制又成為了下一個新的研究問題,在提出“中間器官”的假設(shè)基礎(chǔ)上,學(xué)者進行了各種實驗驗證了,這些結(jié)果已具有神經(jīng)肌肉接點的突觸傳遞理論的雛形,也很好的解釋了潛伏期產(chǎn)生的機制(見表1)。可見,無論是生理學(xué)還是運動生理學(xué)的研究,以問題為導(dǎo)向的研究思路是獲取知識的重要途徑。

表1 骨骼肌興奮收縮耦聯(lián)知識發(fā)展與追溯Table1 Development and Retrospect of Excitation-Contraction Coupling
4 骨骼肌的神經(jīng)支配與隨意運動
19世紀(jì)初,電刺激神經(jīng)引起肌肉收縮被廣泛認(rèn)可,但神經(jīng)沖動傳達給骨骼肌的方式仍然有爭議。一些實驗結(jié)果顯示,在大腦和脊髓被破壞之后心臟仍然會持續(xù)一段時間收縮,另外,刺激心臟和腸道神經(jīng),其肌肉收縮效應(yīng)和骨骼肌收縮不同,認(rèn)為肌肉本身可以產(chǎn)生興奮收縮,當(dāng)時這一觀點占決定性優(yōu)勢。與之相反的另一種觀點是,神經(jīng)對肌肉收縮有調(diào)控作用,電信號通過神經(jīng)將興奮傳播到肌肉,引起肌肉纖維收縮。在這種爭論的背景下,為了驗證神經(jīng)對肌肉的調(diào)控作用,一些神經(jīng)生理學(xué)家設(shè)計了一系列的實驗。1832年,學(xué)者采用6周齡的兔子暴露切斷坐骨神經(jīng),當(dāng)用電偶導(dǎo)線鏈接切斷神經(jīng)兩端,刺激切斷神經(jīng)的中樞時,可引起肌肉收縮,但這種刺激神經(jīng)引起肌肉收縮一段時間內(nèi)會失效,相繼的一些實驗證明骨骼肌興奮性和神經(jīng)力量(nervous power)之間存在著一種基本的關(guān)系[38]。肌肉的可刺激性通常被認(rèn)為是肌肉組織的特性,它在刺激的影響下可以收縮,但離開了神經(jīng)的支配肌肉的興奮性將會耗竭,也證明了肌肉收縮是受神經(jīng)支配的[25],無疑這些研究通過實驗證實了骨骼肌收縮需要神經(jīng)的調(diào)控。同時這一爭論也推動了人們對心肌和平滑肌的深入研究。可見,在研究神經(jīng)支配骨骼肌收縮的過程中,科學(xué)的爭論無疑是促進學(xué)科知識發(fā)展的動力,嚴(yán)謹(jǐn)?shù)目茖W(xué)實驗是驗證和解決爭論的途徑。
對中樞控制與骨骼肌的隨意運動的研究經(jīng)歷了從局部到整體的研究歷程。電刺激引起骨骼肌的收縮是局部實驗的結(jié)果,驗證了神經(jīng)對骨骼肌的支配作用,但是人體是完整的有機體,科學(xué)研究最終還應(yīng)回歸整體。1886年,V.HORSLEY觀察到肌肉反應(yīng)的節(jié)律與神經(jīng)中樞的電興奮所產(chǎn)生的節(jié)律基本相同,骨骼肌的每一次收縮都是由神經(jīng)中樞任何部位的興奮引起的[39]。為了確定神經(jīng)束分支是否對應(yīng)相應(yīng)的骨骼肌,學(xué)者通過對骨盆和肩帶的肌肉與其支配神經(jīng)關(guān)系的相似性進行了觀察和分析,發(fā)現(xiàn)當(dāng)肌肉的位置發(fā)生變化時,可通過其供應(yīng)神經(jīng)來識別肌肉,認(rèn)為肌肉應(yīng)被視為神經(jīng)的終末器官[40]。這些實驗研究成果顯示了骨骼肌收縮需要神經(jīng)支配,中樞興奮可以引起骨骼肌收縮,從整體上使骨骼肌收縮與中樞控制建立了聯(lián)系,為開展骨骼肌的隨意運動研究奠定了基礎(chǔ)。
19世紀(jì)中后期,在體動物實驗對生理學(xué)知識的擴充起到了重要作用,畢竟動物的研究始終不能代替人體的研究,對人體功能和規(guī)律的認(rèn)識仍需以人體作為研究對象。為全方位了解肌肉的收縮,發(fā)現(xiàn)用聽診器可以聽到骨骼肌收縮時的聲音,聽到的聲音通常歸因于周期性活動組織的某些振動,當(dāng)時認(rèn)為正常的肌肉音可能被認(rèn)為是證明正常神經(jīng)沖動是間歇性的重要環(huán)節(jié),連續(xù)的自主收縮是由單個收縮的總和所組成的,就像人工強直是由間斷的刺激造成的,人們認(rèn)為收縮骨骼肌產(chǎn)生的聲音取決于強直性收縮的張力。1885年的研究通過聲學(xué)手段以準(zhǔn)確的音調(diào)進行了研究,發(fā)現(xiàn)在自主收縮中聽到的音調(diào)并不是自然神經(jīng)沖動不連續(xù)的證據(jù),而是僅僅取決于力的變化或刺激分布的顫動[41],1886年,E.A.SCH?FER研究意識對隨意肌節(jié)奏反應(yīng)的影響,發(fā)現(xiàn)人的中樞意識產(chǎn)生的神經(jīng)沖動引起肌肉反映與其他哺乳動物相當(dāng),對人體意志刺激的肌肉反應(yīng)速率與脊髓活動引起的肌肉反應(yīng)速度相同,人骨骼肌長時間的隨意收縮是由每秒8~13次連續(xù)神經(jīng)沖動產(chǎn)生的肌肉不完全強直收縮(見圖9)[42]。在探索這些科學(xué)問題時,采用不同的研究手段可從不同的角度獲取知識,有助于多方位闡釋和解決科學(xué)問題。

圖9 隨意肌收縮記錄儀[42] Figure9 Apparatus Employed to Record the Contractions of the Voluntary Muscles in Man[42]
隨意運動是生理學(xué)研究領(lǐng)域,也是神經(jīng)生物學(xué)研究的內(nèi)容,更是運動生理學(xué)研究的基礎(chǔ)。神經(jīng)科學(xué)的知識是沒有界限的,在運動層面這些知識可以用來解釋中樞控制與肌肉收縮的生理機制,從學(xué)科角度來看,運動生理學(xué)研究也需要掌握中樞控制與肌肉收縮相關(guān)的生理學(xué)、神經(jīng)科學(xué)的知識,從而更好的解決運動科學(xué)中出現(xiàn)的問題。
5 骨骼肌疲勞與工作效率
在農(nóng)業(yè)、手工業(yè)時代體力勞動是創(chuàng)造價值的主要手段,生產(chǎn)勞動離不開勞累,勞累是疲勞的一種,雖然當(dāng)時沒有對疲勞明確定義,但19世紀(jì)中期已經(jīng)有相關(guān)疲勞與抗疲勞的研究,然而對疲勞的研究也是逐步深入的過程。1863年S.HAUGHTON對長時間靜力性收縮引起肌肉不適注意到了這一現(xiàn)象,并對運動時肩關(guān)節(jié)角度變化與參與肌肉不適的關(guān)系進行研究,認(rèn)為靜力性收縮的肌肉維持肢體狀態(tài)的能量消耗可引起肌肉疲勞,并計算了手臂保持水平時靜力性收縮肌肉的能量消耗[43];1871年,S.HAUGHTON始關(guān)注肌肉動力性收縮做功情況,他對投擲、抗阻和維持姿勢的肌肉效率(Useful effect)與肌肉負(fù)荷(weight)進行了一系列的測試,并繪制了函數(shù)關(guān)系圖(見圖10)[44],得出了不同運動形式的肌肉效率與肌肉負(fù)荷的關(guān)系。1875年,F(xiàn).E.NIPHJER對肌肉疲勞與做功(Mechanical Work)的關(guān)系進行多次實驗,提出肌肉收縮到疲勞的時間與肌肉工作時的負(fù)荷、收縮速度以及肌肉恢復(fù)情況有關(guān)[45]。肌肉疲勞程度是影響肌肉工作的重要因素,1875年G.HINRICHS研究了肌肉疲勞的評價方法,指出靜力性工作時肌肉持固定負(fù)荷、保持固定姿勢,在肌肉不出現(xiàn)疼痛的情況下維持這一姿勢和負(fù)荷的時間長短可以評價靜力性疲勞,動力性疲勞可以固定負(fù)荷,做周期性運動,到肌肉疲勞重復(fù)的次數(shù)評價疲勞[46]。這些研究通過觀察記錄相關(guān)數(shù)據(jù),分析運動與疲勞的關(guān)系,獲取了一些評價疲勞的簡單方法,目前這些方法依然是評價運動疲勞的經(jīng)典方法。
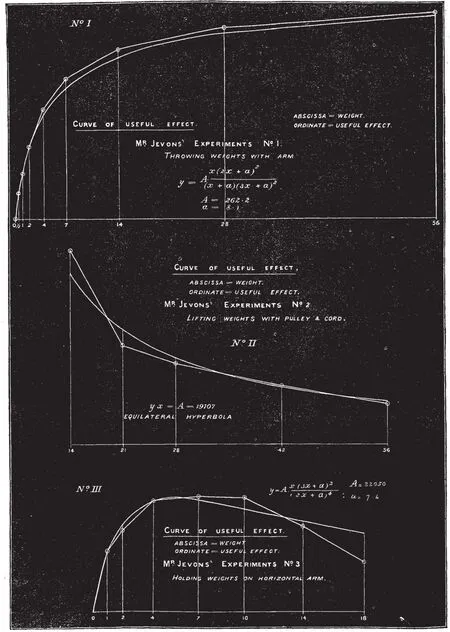
圖10 不同運動形式肌肉效率與肌肉負(fù)荷曲線[44] Figure10 Curve of relation of Useful Effect and Muscle Load in Different Action[44]
在觀察疲勞表象的同時,生理學(xué)家沒有停止探討疲勞發(fā)生的內(nèi)在規(guī)律,生物電與骨骼肌疲勞的研究也在不斷深入,各種組織的電信號檢測儀也在不斷發(fā)展(見圖11)。1871年學(xué)者采用了當(dāng)時精密的儀器做了一系列的實驗,推導(dǎo)出肌肉興奮性的變化在一定程度上影響疲勞的變化[27]。1883年發(fā)現(xiàn)了極度疲勞的肌肉用電刺激引起收縮的潛伏期變長、收縮時間延長、曲線高度降低[34],在對骨骼肌興奮性維持的研究時,發(fā)現(xiàn)離開了血液的營養(yǎng),肌肉的興奮性將會耗竭,認(rèn)為血液為肌肉提供營養(yǎng),并可維持肌肉興奮性[25]。1885年驗證了良好的休息可使連續(xù)電刺激致疲勞的肌肉收縮能力恢復(fù)[35]。1887年,鹽水中肌肉頻繁收縮引起肌肉收縮功能減弱(既肌肉出現(xiàn)疲勞),向鹽水中添加鉀鹽或石灰鹽時肌肉收縮功能加強[47],添加鈣鹽溶液還可以改善肌肉的狀態(tài),并使收縮和放松加速,增加收縮次數(shù)[48]。這一時期對運動疲勞的研究歷程來看,學(xué)者采用觀察法認(rèn)識了疲勞現(xiàn)象,采用實驗法從生物電和肌肉興奮性層面探索了運動疲勞產(chǎn)生的機制,使運動疲勞機制的研究取得了較大進展。
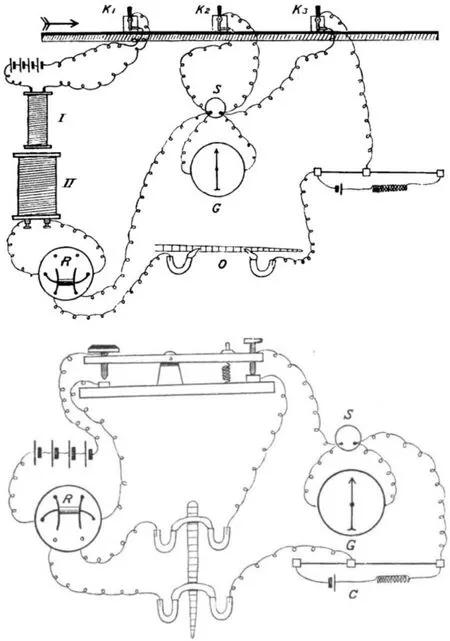
圖11 組織電信號檢測儀示意圖[49] Figure11 Organization of Schematic Diagram of Electrical Signal Detector[49]
疲勞產(chǎn)生的機制與消除是運動生理的主要研究課題,在運動生理學(xué)萌芽前期已廣泛開展了疲勞的相關(guān)研究,當(dāng)時對肌肉做功效率的研究、對疲勞的評價以及各種體外消除疲勞的方法為后續(xù)研究疲勞奠定了基礎(chǔ)。運動疲勞是運動生理學(xué)學(xué)科特征性表現(xiàn)的重要概念,本研究中提供的早期肌肉疲勞素材使疲勞相關(guān)知識體系演變的邏輯起點更加清晰,是運動生理學(xué)演變?yōu)橐婚T獨立學(xué)科的開端。
6 骨骼肌中糖的發(fā)現(xiàn)
肌肉中糖的存在被認(rèn)為是生理化學(xué)領(lǐng)域中最美麗的發(fā)現(xiàn)之一。1849年,德國化學(xué)家H.FEHLING為了鑒定溶液中還原性的糖存在發(fā)明了斐林試劑(fehling's solution),斐林試劑是含有硫酸銅和酒石酸鉀鈉的氫氧化鈉溶液,加熱斐林試劑,若無糖存在溶液中會生成黑色的氧化銅沉淀,若有還原糖存在,則產(chǎn)生黃色或磚紅色的氧化亞銅沉淀,因此,斐林試劑可通過加熱溶液觀察沉淀物的顏色來鑒定糖的存在[50]。在1861年8月,梅斯納宣布肌肉中發(fā)現(xiàn)了糖,這一發(fā)現(xiàn)被記錄在戈廷根皇家科學(xué)院題為“關(guān)于我們對肌肉中物質(zhì)的蛻變的認(rèn)識”的一份聲明中。肌肉中糖的存在開啟了對骨骼肌功能研究的新領(lǐng)域。處于靜息狀態(tài)和運動狀態(tài)的肌肉中糖如何變化?1867年的實驗顯示強直收縮后肌肉中糖含量高于靜止的肌肉,還發(fā)現(xiàn)運動后肌肉中確實存在一種可發(fā)酵的糖,這種糖的量是通過肌肉的運動而增加,并推斷肌肉中的糖確實來源于這個器官本身(而不是血液)[51]。這一研究存在諸多不足,如斐林試劑實驗中肌酐會對檢驗糖的結(jié)果造成干擾、血糖和肌糖原之間轉(zhuǎn)換關(guān)系被忽略等,但也確定了肌肉中重要能源物質(zhì)糖的存在,并指出了運動與糖發(fā)酵有關(guān)系,這無疑對骨骼肌運動功能認(rèn)識的巨大進步。在19世紀(jì)中后期對糖和肌肉的關(guān)系已有相關(guān)實驗,能夠驗證肌肉運動和糖有關(guān)系,并能認(rèn)識到了肌肉中發(fā)酵糖的存在,這為研究運動的能量供應(yīng)和疲勞機制研究提供了突破口(見表2),為運動生理學(xué)形成一門學(xué)科奠定了基礎(chǔ)。

表2 疲勞堵塞學(xué)說知識發(fā)展與追溯Table2 Knowledge Development and Retrospect of Theory of Sporting Fatigue Jam
這期間還有一些運動生理相關(guān)的研究成果相繼出現(xiàn),1871年有實驗發(fā)現(xiàn)血紅蛋白與肌肉活動之間存在一定的關(guān)系[27],1878年作者刺激橈神經(jīng)會引起伸肌反射收縮,導(dǎo)致流量增加,而刺激迷走神經(jīng)影響呼吸肌不會引起伸肌反射收縮,對血流量增加影響不大[52],1887年的實驗顯示肌肉發(fā)酵物可以加速肌肉凝固[53],1888年發(fā)現(xiàn)咖啡因?qū)χ袠猩窠?jīng)系統(tǒng)的作用可引起肌肉強直[54]。隨著對骨骼肌研究的不斷深入,使骨骼肌運動功能的知識不斷積累,逐步形成了系統(tǒng)化的知識體系,為運動生理學(xué)科的形成奠定了基礎(chǔ)。
7 小結(jié)
17~19世紀(jì)生理學(xué)家對骨骼肌的研究歷程來看,為了解決生活實踐活動中肌肉相關(guān)疼痛疾病,探索了骨骼肌形態(tài)、結(jié)構(gòu)和功能,這些內(nèi)容屬于普通生理學(xué)范疇,為了對骨骼肌機能進行深入研究,骨骼肌收縮的生物電、潛伏期和神經(jīng)支配相關(guān)的專門生理學(xué)研究隨之興起,這種專門生理學(xué)的研究對骨骼肌收縮本質(zhì)的認(rèn)識起重要推動作用。同時,在研究骨骼肌潛伏期時學(xué)者采用了比較的方法進行探討,發(fā)現(xiàn)了骨骼肌在相應(yīng)電刺激條件下的不同適應(yīng),為全面了解骨骼肌的功能提供了實驗支持。比較生理學(xué)的進步促進了應(yīng)用生理學(xué)的發(fā)展,骨骼肌不同條件下疲勞及消除的比較研究與人類的生產(chǎn)、生活密切相關(guān),所以,對骨骼肌疲勞和工作效率的研究將生理學(xué)發(fā)展推到了運動生理學(xué)層面,因此,運動生理學(xué)是在普通生理學(xué)和比較生理學(xué)等學(xué)科知識的基礎(chǔ)上,研究生產(chǎn)生活中運動相關(guān)的生理現(xiàn)象和規(guī)律的一門應(yīng)用生理學(xué)。
1889年,第1本運動生理學(xué)教材問世之前,骨骼肌運動功能的研究并非一帆風(fēng)順,但也已取得了一些可喜的成果。總結(jié)這一階段研究的經(jīng)驗與教訓(xùn),對未來運動生理的發(fā)展有如下啟示:運動生理學(xué)的研究應(yīng)以問題為導(dǎo)向,以解決現(xiàn)實生活中人們切實遇到的運動需求問題是研究出發(fā)點和落腳點;對運動生理學(xué)中的未解問題進行合理的推論,鼓勵開展自由的學(xué)術(shù)爭論,通過嚴(yán)謹(jǐn)?shù)目茖W(xué)實驗來驗證和解決未解問題,可促進運動生理學(xué)知識的豐富和發(fā)展;運動生理的研究從運動現(xiàn)象開始,逐步深入到器官、細(xì)胞、分子水平,但也要從整體方面考慮機體的調(diào)控;1867年,M.ROUGET明確提到使用肌動描記器研究肌肉功能[55],在運動領(lǐng)域;1890年,意大利學(xué)者A.MOSSO[56]使用肌動描記器首次記錄了肌肉疲勞曲線。因此,解決體育領(lǐng)域的科學(xué)問題時,可更早的采用生命科學(xué)領(lǐng)域的先進技術(shù)和手段,以便運動生理學(xué)與生命科學(xué)在實驗手段上站在同一科學(xué)技術(shù)水平的高度。
猜你喜歡
現(xiàn)代電生理學(xué)雜志(2022年2期)2022-06-26 12:24:16
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
廣西醫(yī)科大學(xué)學(xué)報(2021年10期)2021-11-13 08:31:00
天津外國語大學(xué)學(xué)報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
現(xiàn)代畜牧科技(2021年7期)2021-07-28 06:39:58
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設(shè)計與研究(2019年4期)2019-05-21 07:21:24
現(xiàn)代電生理學(xué)雜志(2018年1期)2018-04-08 07:27:19
汽車工程學(xué)報(2017年2期)2017-07-05 08:13:02
