湖北海棠枝干潰瘍病病原鑒定及生理學特性研究
2019-09-09 09:24:58張兆霞郭慧玲劉會香崔玉國申衛星王玉杰
山東林業科技 2019年4期
關鍵詞:生長
張兆霞,郭慧玲*,劉會香,崔玉國,申衛星,王玉杰,陳 巖
(1.泰山森林病蟲害防治檢疫站,山東 泰安271000;2.泰山風景名勝區管委會,山東 泰安 27100;3.山東農業大學,山東 泰安271018)
湖北海棠(Malus hupehensis(Pamp.)Rehd..)位于泰山南天門海拔1300m以上,在天街至碧霞祠一帶成片分布,因為適應泰山極頂的地理氣候環境,湖北海棠常被稱作“泰山海棠”,是泰山高海拔地區特有的水土保持和景觀樹種。每年春季岱頂大片海棠綻開蓓蕾,連天開放,碧霞祠一帶像是成了花的海洋,秋季則紅果累累,掛滿枝頭。泰山現有海棠林大部分為上世紀60年代人工造林而成,由于林齡過熟和山頂特殊氣候原因,普遍存在生長不良、樹勢衰弱,甚至出現樹木干枯死亡情況。對湖北海棠林分進行有害生物種類調查的基礎上,確定湖北海棠生長衰弱的原因主要是由于枝干潰瘍病的病菌侵染擴散所致。本文野外采集湖北海棠枝干潰瘍病的病菌病斑進行室內培養和分子鑒定,為下一步科學治理海棠潰瘍病提供依據。
1 材料與方法
1.1 試驗材料
從野外發生枝干潰瘍病的湖北海棠樹上剪取若干病枝條。
1.2 試驗方法
1.2.1 病菌的分離方法
在實驗室從海棠枝干病斑處切取大小約5mm病組織共15塊,采用常規組織分離法。先移置于PDA培養基平板放在25℃恒溫箱中培養2~3天[5]。統計各菌株出現的概率,純化后4℃冰箱保存備用。
1.2.2 病菌培養
湖北海棠潰瘍病菌分離純化后在25℃培養5天后,觀察其菌落特征,用顯微鏡觀察病菌子實體大小、形態及孢子的形態。在菌絲體中鉆取5個菌餅,將菌接種到PD(液體馬鈴薯)培養基中,放置于28℃,200rpm恒溫搖床上培養兩天,于-20℃保存。
1.2.3 病菌DNA的提取
DNA的提取采用2×CTAB法,將培養好的菌絲烘干加入液氮冷凍研磨至細小粉末,取0.25g粉末加入預熱的2-ME/CTAB抽提液(1ml)中,65℃保溫45~60min,顛倒混勻,加入等體積酚:氯仿:異戊醇(25:24:1)溶液混勻,12000rpm/min 離心 30min,取上清加入 0.5~0.6(350uL)倍體積冰冷的異丙醇,顛倒混勻,-20℃放置20min;12000rpm/min離心10min;去上清夜,DNA沉淀以70%乙醇洗滌并干燥后溶于50ug/mL RNase的TE緩沖液,置于-20℃保存備用。最后用10g/l瓊脂糖凝膠檢測提取DNA的質量。
1.2.4 rDNA及ITS序列擴增
利用真核生物的基因通用引物ITS1/ITS4進行ITS序列擴增,引物序列為 ITS1 (5’-TCCGTAGGTGAACCTGCGG-3’) 和 ITS4(5’ -TCCTCCGCTTATTGATATGC-3’)。PCR 總反應體系為 25 ul,其中10×PCR Buffer為2.5ul,Mg2+1ul,dNTP 1ul,ITS1和 ITS4各為 2uL,Taq酶 0.5uL,ddH2O14ul,DNA模板2uL,混勻后進行PCR擴增。PCR反應條件為94℃,預變性3min后進入以下循環:94℃變性 1 min,55℃退火 1 min,72℃延伸 2 min,30個循環后于72℃下10 min,并在4℃下保存備用。PCR反應結束后進行10g/l瓊脂糖凝膠電泳。
1.2.5 擴增產物純化及測序鑒定與系統發育樹的構建
利用Biospin膠DNA回收試劑盒對DNA進行純化擴增并電泳。染色后在凝膠成像系統紫外燈下觀察。從NCBI數據庫中下載公認Botryosphaeria dothidea序列為參考,參比菌株具體信息為KT182890.1,FJ493245.1,KF294012.1,MH715242.1,JX241645.1與供試菌株相應序列一起采用Mega5.1中ClustalW進行比對并校正。輪紋病菌以Fusarium oxysporum(JK747249.1 4-C10)為外源,基因序列自展法(Bootstrap)檢測,重復 1000次,構建鄰接樹(Neighbor-Joining Tree)。腐爛病菌以Botryosphaeria dothidea序列為外源,其他方法相同。
1.2.6 致病性測定
采用菌餅接種法[3]。選取PDA培養基上生長5天的菌餅采取刺傷接種健康蘋果果實上,以不接種病菌的蘋果果實為對照,重復15次,5天后觀察接種結果。
1.2.7 病菌生理學特性研究
1.2.7.1 病菌對不同溫度的適應性
將海棠輪紋病和腐爛病病菌分別接種于PDA培養基,打孔器內徑為0.5mm,分別于10℃、15℃、20℃、25℃、30℃、35℃和 40℃恒溫培養箱中培養,每隔一天后測量菌落直徑,每個菌株重復3次。
1.2.7.2 病菌對不同pH的適應性
將湖北海棠輪紋病菌接種于pH 為 3、4、5、6、7、8、9、10和 11的 PDA培養基上,打孔器內徑為0.5mm,然后置于恒溫培養箱中培養,每隔一天后測量菌落直徑,每個菌株重復3次。
2 結果與分析
2.2 病原菌的分離
選取的15塊病組織經分離純化共獲得15個菌株,放于25℃恒溫箱備用。
2.3 病原菌的鑒定
2.3.1 培養學特性
2.3.1.1 海棠輪紋病菌培養學特性觀察
海棠輪紋病菌在PDA培養基上生長迅速,培養初期菌落為白色,疏松,棉絮狀,氣生菌絲較發達,菌落邊緣整齊或不整齊。培養3d后,菌落逐漸變成灰色,10d左右,菌落呈深灰色至黑色。顯微鏡檢發現,菌絲初期無色,后為深褐色,菌絲粗細不勻,分枝發達,有隔(如圖1)。

圖1 海棠輪紋病菌的培養學特征(5d)
2.3.1.2 泰山海棠腐爛病菌培養學特性觀察
海棠腐爛病菌在PDA培養基上初為白色,后變為淺黃色,菌絲匍匐,氣生菌絲不發達,基內生長,菌落邊緣整體,3~5天長滿整個平皿,此后菌絲繼續向培養基內生長,后期20天長出褐色的分生孢子器或者直接溢出分生孢子堆(如圖2)。

圖2 海棠腐爛病菌的培養學特征(20d)
2.3.2 形態學特性
2.3.2.1 海棠輪紋病菌鏡檢形態學特性
解剖鏡下鏡檢發現,海棠輪紋病菌分生孢子器呈黑褐色,初埋生于寄主表皮下,后突破表皮部分外露。分生孢子多為單生,單腔室,扁球形或球形,有孔口,大小約 110.12~154.28μm ×82.74~150.23μm,平均 129.67~114.27μm。內壁生分生孢子梗,孢子梗無色,不分支。分生孢子梭形,正直,無色,單孢,頂端鈍圓,基部平截,大小為16.83~24.55μm ×5.27~8.17μm,平均 18.35 ×6.34μm(如圖3)。
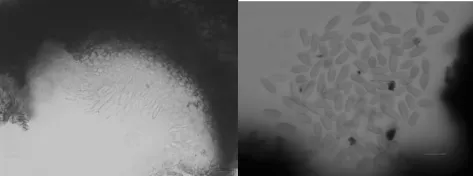
圖3 海棠輪紋病菌在解剖鏡下鏡檢形態特征
2.3.2.2 泰山海棠腐爛病菌鏡檢形態學特性
解剖鏡下鏡檢發現,腐爛病菌分生孢子器著生于外子座內,扁瓶狀,480~1600μm× 400~960μm,多個腔室,相通,同一孔口,室壁密生分生孢子梗;分生孢子單孢,臘腸形或香蕉形,3.6~8.0μm×0.8~1.7μm。孢子溢出形成橘黃色孢子角(如圖4)。

圖4 海棠腐爛病菌形態特征
2.3.3 泰山潰瘍病病原菌的分子鑒定
2.3.3.1 泰山海棠輪紋病菌ITS序列測定及系統樹構建
海棠輪紋病菌經PCR檢測,其大小介于500-750bp之間,序列測定發現海棠輪紋病菌的ITS序列長583bp。經Blast軟件分析對比,分子鑒定認為泰山海棠輪紋病是子囊菌門(Ascomycotina),葡萄座腔菌(Botryosphaeria dothidea),其系統樹構建如圖5所示。

圖5 基于ITS基因序列的海棠輪紋病菌系統發育樹
由圖5看出,所測定的序列確與Botryosphaeria dothidea真菌的系統發育最近,再結合病菌的培養學和形態學特征,可以明確泰山海棠輪紋病菌為Botryosphaeria dothidea。
2.3.3.2 泰山海棠腐爛病菌ITS序列測定及系統樹構建
海棠腐爛病菌系統樹構建如圖6所示。

圖6 基于ITS基因序列的海棠腐爛病菌系統發育樹
由圖6看出,海棠腐爛病菌經過序列測定和系統樹構建并結合病菌的培養學和形態學特征,鑒定泰山海棠腐爛病是由Valsa ceratosperma真菌引起。
2.4 海棠潰瘍病病原菌的致病性測定

圖7 海棠輪紋病和腐爛病菌接種蘋果后的不同癥狀
室內人工接種蘋果果實(如圖7)發現,分離的2種病原菌對蘋果果實均有致病性,其發病癥狀和蘋果輪紋病、腐爛病病癥表現一致。再次證明Botryosphaeria dothidea是泰山海棠輪紋病的病原菌;Valsa ceratosperma是泰山海棠腐爛病的病原菌。
2.5 病菌對溫度的適應性
在不同溫度下的PDA培養基上,湖北海棠輪紋病菌和腐爛病菌的生長特性如圖8和表1所示。

圖8 湖北海棠輪紋病菌在不同溫度下的生長特性(3d)(從左到右從上到下依次是在 10℃、15℃、20℃、25℃、30℃和35℃輪紋病菌的生長狀況)

表1 湖北海棠輪紋病菌和腐爛病菌不同溫度生長情況/mm
由圖8和表1可以看出,泰山海棠輪紋病菌和腐爛病菌在10℃~35℃條件下均可生長,隨著溫度的升高,兩種病菌的生長速度加快,病菌在25℃生長最好,其次是30℃、20℃和15℃,其他溫度下病菌的生長速度明顯減慢,腐爛病菌的耐高溫能力稍強于輪紋病菌。對溫度的適應性更強一些。
2.6 病菌對不同pH的適應性
泰山海棠輪紋病菌和腐爛病菌在25℃下,不同pH的PDA培養基生長結果見表2。由表4可知,不同pH條件下,海棠輪紋病菌和腐爛病菌生長特征存在差異,海棠輪紋病菌的生長范圍稍窄一些,在pH4~10之間;而海棠腐爛病菌在pH3~11之間均可生長。隨著pH的升高,兩種病菌的生長速度加快,其中腐爛病菌在pH6時生長最快,輪紋病菌在pH5生長最快,總體表現為輪紋病菌生長稍慢于腐爛病菌生長,說明海棠腐爛病菌對酸堿度的適應性更強一些。

表2 湖北海棠輪紋病菌和腐爛病菌不同pH生長情況/mm
3 結論與討論
通過海棠枝干潰瘍病原菌的培養學、形態學、分子序列特征、系統樹構建及致病性測定表明,海棠枝干潰瘍病主要由輪紋病和腐爛病兩種病害所致,鑒定病原菌2種,其中海棠輪紋病原菌為葡萄座腔菌(Botryosphaeria dothidea);海棠腐爛病原菌為 Valsa ceratosperma。生理學研究表明,兩種病菌對溫度和pH的要求均隨著溫度或者酸堿度提高而逐漸生長加速,但不同病菌的適應性不同,其中腐爛病菌對溫度和酸堿度的適應性更強一些。2種病菌造成湖北海棠枝干潰瘍病的發生在國內鮮有報道,尤其是大面積分布在泰山高海拔區1300m處的湖北海棠林分。本研究明確了湖北海棠枝干潰瘍病原菌發生的種類及致病病原菌,為下一步綜合治理湖北海棠枝干潰瘍病提供了理論支持。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
