小檗堿對EAE小鼠星形膠質細胞S1PR1、3受體表達影響的研究
2019-11-09 02:19:20曾思玉余巨明秦新月羅家明
中風與神經疾病雜志 2019年10期
陳 蓉,曾思玉,張 微,李 艷,余巨明,秦新月,羅家明
多發性硬化(multiple sclerosis,MS)是一種中樞神經系統(central nervous system,CNS)自身免疫性疾病,臨床呈慢性反復發作過程,中青年人群多見[1]。MS的病因復雜,發病機制尚待探明,可能在遺傳因素與環境因素交互作用下,自身免疫系統紊亂而誘發的中樞神經脫髓鞘[2]。脫髓鞘造成軸索損傷,甚至神經元死亡,使患者神經退行性病變,引起患者神經功能障礙,所致的臨床表現為殘疾,伴發其他并發癥時還會導致患者死亡[3]。MS主要病理改變是髓鞘脫失、膠質增生、軸索損傷等[4]。MS目前尚無治愈的方法,治療手段亦有限[5]。因此,研究MS的病因和發病機制,探尋有效的治療措施,成為MS亟待解決的問題[6]。
在MS及其動物模型實驗性自身免疫性腦脊髓炎(experimental autoimmune encephalomyelitis,EAE)研究中發現,星形膠質細胞(astrocyte,AST)活化在其發生、發展過程中扮演重要角色[7,8]。MS及EAE小鼠中樞神經系統炎性反應主要由AST維持,AST活化后釋放的一系列致炎性細胞因子和趨化因子,是中樞神經系統炎癥維持和放大的重要因素[9,10]。AST還是構成血腦屏障的主要細胞,活化后的AST釋放的致炎性細胞因子和趨化因子,可破壞血腦屏障,增加血腦屏障通透性,血腦屏障通透性增加也是MS發病的核心環節[10~12]。
1-磷酸鞘氨醇(sphingosine 1-phosphate,S1P)是細胞膜脂質代謝產物,系第二信號分子中的一種,在多種生理和病理過程中發揮重要作用[13]。Choi等[14]在對芬戈莫德(Fingolimod,FTY720)的中樞機制研究中發現,FTY720通過抑制AST上的S1PR1受體與S1P結合,從而緩解EAE小鼠臨床癥狀。該研究團隊進一步研究表明,AST上的S1PR1受體與S1P結合后,可導致EAE小鼠膠質增生、脫髓鞘、軸索損傷和致炎性細胞因子(如IL-1β、IL-6、IL-17等)的釋放,故AST上S1PR1與S1P結合是AST介導EAE發病的關鍵。MS患者腦組織中AST上S1PR3受體表達量較正常高[13],在EAE小鼠離體實驗研究中發現,S1P與其受體S1PR3結合后,通過S1P/S1PR3通路激活AST,是AST激活和引發膠質增生以及維持炎性反應的另一重要環節[15,16]。
小檗堿(Berberine,BBR)即黃連素,是從黃連、黃柏等中藥中提取的結構明確的中藥單體,有抗感染、驅蟲、止瀉等功效,且毒副作用小,使用安全,臨床應用廣泛。在阿爾茨海默病(Alzheimer disease,AD)和腦缺血性損傷等中樞神經系統疾病研究中發現,BBR能通過血腦屏障,直接作用于中樞神經系統,抑制致炎性細胞因子的釋放、調控神經遞質釋放、抗氧化和自由基的清除等[17~20]。研究表明,BBR可緩解EAE小鼠神經缺損癥狀,保護血腦屏障,但其具體的作用機制并不清楚[22,23]。研究顯示在小鼠糖尿病腎病模型中,BBR能抑制腎臟細胞鞘氨醇激酶1(sphingosine kinase 1,SPHK 1)表達,SPHK 1為生成鞘氨醇的限速酶,SPHK 1將鞘氨醇磷酸化為S1P。BBR抑制SPHK 1表達后,可減少S1P的產生,提示BBR能夠影響S1P的生成[24]。
本研究通過構建EAE模型小鼠,用不同劑量的BBR干預,觀察小鼠的臨床表現,對各組小鼠進行神經功能缺損進行評分。檢測小鼠AST上S1PR1、3受體表達情況,了解中樞神經炎細胞浸潤、脫髓鞘程度和膠質增生情況。用BBR干預原代細胞培養的AST,檢測BBR對離體AST的S1PR1、3受體表達的影響。
1 材料與方法
1.1 材料 實驗小鼠:體重15~20 g、8~10周齡的健康雌性C57BL/6小鼠60只,在川北醫學院實驗動物中心SPF級環境飼養,自由攝食,室內溫度25 ℃,保持自然晝夜節律光照。所有實驗均獲得川北醫學院倫理委員會批準。MOG35-55多肽(sigma公司,美國)、完全弗氏佐劑(sigma公司,美國)、(sigma公司,美國)、BBR(sigma公司,美國)、卡介苗(中國藥物研究所)、一抗(abcom公司,英國)、脂多糖(lipopolysaccharide,LPS)、二抗(abcom公司,英國)。
1.2 方 法
1.2.1 分組及造模方法 將實驗動物隨機分為5組,分別為CFA組(n=6):小鼠免疫建模時,用生理鹽水代替MOG35-55多肽片段構建模型,免疫后10 d,為不含免疫抗原乳劑的空白對照組;EAE組(n=6):MOG35-55多肽片段構建模型,為疾病組;BBR干預組[BBR按照50 mg/(kg·d)劑量灌胃,n=6]:構建EAE模型,從建模后第10天開始,每日給予不同劑量[50、100、300 mg/(kg·d)]BBR灌胃,為疾病干預組。CFA組和EAE組在建模后第10天起給與等量生理鹽水灌胃。采用經典造模方法,將MOG35-55多肽用pH值為7.2、濃度為0.01 mol/L的PBS液稀釋后,與含卡介苗10 g/L的完全弗氏佐劑(complete Freund’s adjuvant,CFA)等體積混合,使其終濃度為300 μg /ml。使用注射器通過連接后低溫環境反復推拉至油包水狀態,即為抗原乳劑。抗原乳劑注射方法采用經典的多點足墊皮下注射法,每只小鼠分4點注射,共注射0.2 ml抗原乳劑[MOG35-55多肽片段含量為300 μg/(ml·只)]。在抗原乳劑免疫后的0 h和48 h,分別給予百日咳菌稀釋液0.1 ml/只,腹腔注射(含百日咳桿菌活菌109個)作為增敏劑以提高模型發病率。每天觀察各組小鼠神經功能狀況、臨床表現,參照Kono[25]方法,對各組小鼠進行神經功能缺損程度評價。具體評分辦法如下:0分為正常,無發病;1分為動物尾部無力;2分為翻身不能;3分為雙后肢癱瘓;4分為四肢癱瘓;5分為瀕死狀態或死亡。經培訓的兩位實驗人員分別評分后取平均分作為最終得分。
1.2.2 LFB染色 LFB染色了解各組小鼠腦組織脫髓鞘情況。在各組小鼠干預后30 d,終止試驗,處死小鼠,用多聚甲醛心臟灌注法取材。LFB染色觀察脫髓鞘程度,評分標準如下:1分為輕微脫髓鞘;2分為顯著脫髓鞘;3分為脫髓鞘斑塊多處融合;4分為廣泛中樞脫髓鞘并伴中樞神經系統實質內炎細胞浸潤。每個組織計數方法取5個視野計算平均值。
1.2.3 免疫熒光雙標 運用免疫熒光法檢測各組小鼠脊髓和腦組織膠質增生情況,鏡下觀察并采集圖像,隨機挑選3個200倍視野拍照保存,Image-Pro Plus 6.0軟件分析計算出累積光密度值(integrated optical density,IOD)。免疫熒光雙標檢測各組小鼠脊髓和腦組織AST上S1PR1、S1PR3表達情況,鏡下觀察并采集圖像,分析方法同前述。
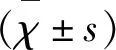
2 結 果
2.1 小鼠臨床表現 CFA組小鼠無發病。EAE組小鼠呈亞急性發病,精神食欲差,活動少,毛色暗淡,體重下降。EAE組小鼠抗原免疫后第8天,部分小鼠陸續發病;免疫后16~20 d,大部分小鼠癥狀達到頂峰,隨后癥狀逐漸好轉,但部分小鼠癥狀持續到免疫后30 d。BBR干預組:該組小鼠總體發病比EAE組更輕微,尤其100、300 mg/(kg·d)劑量組癥狀減輕顯著。BBR干預組小鼠體重下降程度和肢體癱瘓情況較EAE組輕,隨著BBR劑量增加,小鼠體重下降程度、癱瘓情況減輕越明顯(見圖1)。
2.2 各組小鼠腦組織炎性浸潤和脫髓鞘改變情況 CFA組小鼠腦組織切片檢測發現炎細胞浸潤較少。EAE組小鼠腦組織切片中,炎細胞浸潤較多,主要聚集在小血管周圍,腦實質內也有炎細胞浸潤。BBR干預組腦實質中炎細胞浸潤較EAE組少,100/300 mg/(kg·d)組減少更明顯。炎性程度評分結果提示100 mg/(kg·d)和300 mg/(kg·d)劑量組較EAE組低(P<0.01)(見圖2)。CFA組小鼠腦組織無明顯脫髓鞘改變。EAE組小鼠腦組織可見明顯髓鞘破壞,主要表現為LFB染色局部呈淡染,髓鞘纖維纖細,間距增寬,空泡區較多,部分切片出現髓鞘斷裂或缺如。腦組織脫髓鞘評分結果顯示,100 mg/(kg·d)和300 mg/(kg·d)劑量組較EAE組低(P<0.01)(見圖2)。
2.3 熒光免疫檢測各組小鼠腦和脊髓組織膠質增生情況 通過GFAP標記腦和脊髓組織膠質增生程度結果顯示,EAE組小鼠腦組織膠質增生明顯,小檗堿干預各組膠質增生受到抑制。通過IOD值分析顯示,小檗堿干預各組小鼠腦和脊髓GFAP表達較EAE組小鼠低,差異有統計學意義(P<0.01),劑量越大,減輕程度越明顯(見圖3)。
2.4 各組小鼠腦和脊髓組織星形膠質細胞上S1PR1、3受體 各組小鼠進行GFAP和S1PR1、S1PR3受體熒光標記,結果顯示:EAE組小鼠腦組織AST上S1PR1、3受體表達量較CFA組高,差異有統計學意義(P<0.01)。BBR各劑量干預組小鼠腦組織中,AST上SPR1、3受體表達量較EAE組減少顯著,差異有統計學意義(P<0.01)。BBR各干預組間存在劑量梯度變化差異,劑量越大,AST上S1PR1、3受體表達量越少,差異有統計學意義(P<0.01),劑量越大,表達量越少(見圖4)。

A:各組小鼠神經功能評分變化趨勢;B:各組小鼠最大神經功能缺損評分比較;C:各組小鼠神經功能缺損累積評分比較
圖1 各組小鼠神經功能評分變化趨勢、最大評分和累積評分
3 討 論
傳統中藥黃連在中國擁有長達千年的臨床使用歷史,黃連素是從中藥黃連中提取的天然中藥單體,具有廣泛的生物活性[21]。研究發現BBR通過作用多條信號通路發揮重要的生物活性作用,使其用于多種疾病的治療[18,24]。近年來,BBR因其易透過血腦屏障,調控神經遞質、抑制炎細胞因子釋放、清除自由基和抗氧化等功能,在中樞神經系統疾病中的治療作用越來越受到重視[17,19,20]。
MS發病機制尚不清楚,膠質增生是其主要的病理改變。膠質增生即中樞神經系統中大量AST活化、增生,膠質增生后AST標記物GFAP顯著升高,AST活化后釋放10余種致炎性細胞因子,維持和放大中樞神經系統炎癥[26,27]。MS膠質增生過程如下:少突膠質細胞上髓鞘病理性破壞后,使小膠質細胞極化,極化后的小膠質細胞釋放的致炎性細胞因子作用于AST,進一步激活AST,AST是致炎性細胞因子的主要效應細胞[28]。膠質增生嚴重程度與MS和EAE的臨床癥狀呈正相關,也與神經功能評分呈正比[8,13,29]。S1P與表達細胞表面的5種G蛋白藕聯受體1-磷酸鞘氨醇受體1-5(sphingosine 1-phosphate receptor1-5,S1PR1-5)結合后,引發一系列生理效應,如調節細胞生存、增殖、遷移以及傳遞各種細胞信號等[13,27]。同時,S1P亦是導致中樞神經系統膠質增生的重要因素,研究發現脊髓損傷后,膠質增生與S1P含量呈正相關[14]。顱內注射外源性S1P能加重腦組織膠質增生,提示升高的S1P可導致中樞膠質增生[30]。S1P還參與缺血性腦損傷、MS等多種疾病的發病[14~16]。研究顯示MS患者顱內病灶中,S1P含量較正常高,提示MS發病可能與S1P升高有關[13,17,30]。
Kim等[31]在MS的另一種動物模型雙環已二酮草酰二腙(cuprizone,CPZ)誘導的脫髓鞘模型中(該模型僅用毒性的CPZ直接破壞中樞神經系統髓鞘,無外周免疫誘導機制參與),發現AST上S1PR1與S1P結合是導致膠質增生的重要環節。由于該研究使用的模型是非免疫誘導直接毒性作用造成的中樞脫髓鞘和膠質增生,由此說明S1P與S1PR1結合導致膠質增生,且不依賴外周免疫系統參與。S1P與其受體結合后的作用在MS和EAE的發病中發揮重要作用,該通路成為近年MS研究的熱點[13~15]。
S1P信號通路在EAE小鼠或MS患者神經退行性病變中起重要作用[28]。S1P是導致中樞膠質增生的重要原因之一,腦組織是S1P含量最高的器官,病理情況下濃度更高[15]。顱內的S1P除來源于血液之外,還可內源性產生。Choi等[14]發現AST上的S1PR1受體與S1P 結合是導致EAE小鼠脫髓鞘、膠質增生、軸索損傷和致炎性細胞因子釋放的關鍵。由此推測,AST上S1PR1受體與S1P結合可能是AST激活的關鍵環節,S1P亦是導致中樞膠質增生的重要原因之一。本研究首先證實EAE小鼠AST上S1PR1、3受體表達量增高,而BBR能抑制EAE小鼠模型AST上S1PR1、3受體表達。提示BBR通過抑制S1PR1、3受體表達,減少S1P與S1PR1和S1PR3兩種受體結合,這可能是BBR抑制AST激活的原因之一。本研究還發現BBR緩解EAE小鼠臨床癥狀、降低神經功能缺損評分,與劑量大小呈正相關,100 mg/(kg·d)和300 mg/(kg·d)劑量干預組較50 mg/(kg·d)效果更明顯。
與以往研究相比,本研究有兩個不同之處:第一,既往研究僅給予單一劑量的BBR干預,不清楚劑量大小對MS 發病是否有影響。本研究設置了3個劑量梯度,觀察到臨床療效、膠質增生和炎細胞浸潤和脫髓鞘程度等,劑量越大,臨床癥狀和脫髓鞘程度越輕,膠質增生和炎細胞浸潤、致炎性細胞因子釋放越少。本研究進一步證實BBR對EAE小鼠有一定治療效果,為以后的臨床研究和治療提供理論依據。第二,既往研究從BBR對EAE外周免疫反應以及其對血腦屏障的影響角度入手,未研究其在EAE發病的中樞損傷機制[20,22]。本研究針對EAE發病的關鍵環節,中樞神經系統AST上S1P受體入手,對其作用機制進行更深層次的探索。
本研究發現經BBR干預后,EAE小鼠AST特異標記物GFAP及致炎性細胞因子IL-1β、IL-6表達顯著下降,提示BBR能抑制AST活化。BBR干預后的EAE小鼠膠質增生和炎細胞浸潤減少,臨床癥狀和脫髓鞘程度減輕,且與劑量呈正相關。本研究證實了BBR能緩解EAE小鼠臨床癥狀。同時還發現BBR能抑制中樞膠質增生和脫髓鞘,可能與其抑制AST上S1PR1、3受體表達、致炎性細胞因子釋放有關,為BBR在MS的基礎研究與臨床應用提供了一定的理論依據。

A:各組小鼠腦組織HE染色示炎性細胞浸潤情況;B:各組小鼠腦組織炎性細胞浸潤評分結果比較;C:各組小鼠腦組織LFB染色示脫髓鞘情況;D:各組小鼠腦組織脫髓鞘評分結果比較
圖2 各組小鼠腦組織HE和LFB染色

A:各組小鼠腦組織膠質增生情況;B:各組小鼠膠質增生評分比較情況

A:各組小鼠小鼠腦組織星形膠質細胞S1PR1、3表達情況;B:各組小鼠腦組織星形膠質細胞S1PR1受體表達比較;C:各組小鼠腦組織S1PR3受體表達比較
圖4 各組小鼠腦組織S1PR1、3表達情況
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
課堂內外·初中版(科學少年)(2023年10期)2023-12-10 00:43:06
全科護理(2022年10期)2022-12-26 21:19:15
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
國際放射醫學核醫學雜志(2021年10期)2021-02-28 08:41:58
學苑創造·A版(2020年9期)2020-10-13 09:41:02
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
