小麥面團粉質特性的遺傳分析
2019-11-13 03:39:12于紅彩胡鐵柱李淦
江蘇農業科學 2019年17期
于紅彩 胡鐵柱 李淦

摘要:以6個不同品質類型的小麥品種為親本,按GriffingⅡ雙列雜交方法配置15個雜交組合,分析研究了面團粉質特性的遺傳規律。結果表明,所有面團粉質參數的一般配合力(GCA)方差均達到顯著水平,除弱化度和粉質質量指數外,另外2個參數的特殊配合力(SCA)方差也達到顯著水平;面團粉質參數的廣義和狹義遺傳力均較高,加性方差明顯大于顯性方差,表明弱化度和粉質質量指數主要受加性效應影響,而面團形成時間和穩定時間同時受加性和非加性效應控制,且以加性效應為主。綜合來看,鄭2441×濟麥20屬于強優勢組合。
關鍵詞:小麥;面團粉質特性;雙列雜交;遺傳分析
中圖分類號:S512.103 ? 文獻標志碼: A ?文章編號:1002-1302(2019)17-0082-03
面團粉質特性是衡量小麥品質的重要指標之一[1],與面包、饅頭、面條、餅干等加工品質密切相關,其檢測指標主要包括吸水率、形成時間、穩定時間、弱化度和粉質質量指數等[2]。其中,面團穩定時間是評價面包品質最重要的指標之一[3],與面條的韌性、硬度和粘彈性呈顯著正相關,與煮面的外觀評價呈顯著負相關[4];另外,李蓓蓓等研究表明,吸水率、形成時間、穩定時間與酥性餅干評分呈顯著負相關[5]。因此,對面團粉質特性的研究可預測和控制專用粉的品質。鑒于當前我國食品加工專用粉的需求量較大,因此對小麥面團粉質特性的研究顯得尤為重要。
迄今,國內外學者已對小麥產量、穗粒數、株高等農藝性狀[6-10]以及淀粉含量、面筋含量等品質性狀[11-13]的遺傳研究做了大量報道,得出很多具有指導意義的結論,但有關小麥面團粉質特性遺傳研究報道相對較少,因此亟需對其遺傳規律進行深入探討。
雙列雜交一直是進行植物數量性狀研究的主要方法之一,該方法主要是通過分析組合材料的特點,進而篩選出符合目標性狀的優良親本組合[14],在棉花[15]、玉米[16]、水稻[17]等農作物育種上也均有廣泛應用。該遺傳統計分析方法主要分為2類:一是Griffing的配合力分析法,二是Hayman的遺傳效應分析法[18-19]。這2種方法至今仍在廣泛應用,并推動著數量性狀遺傳和育種的深入研究和發展。
本試驗選用6個不同品質類型的小麥品種為試驗材料,分析研究了面團粉質特性的遺傳規律,旨在為小麥品質育種提供理論依據。
1 材料與方法
1.1 試驗材料
小麥6個親本分別為濟麥20、SN01-24、百農64、周麥16、鄭麥004、鄭2441,按GriffingⅡ雙列雜交方法配置15個雜交組合。親本和雜交種F1(共21份材料)于2013—2014年度種植于河南科技學院試驗基地(河南省新鄉縣朗公廟鎮),完全隨機設計,每份材料種植4行,行長4 m,行距25 cm,每行播種80粒,肥水管理同一般大田。成熟時按組合收獲,脫粒,晾曬,常溫儲藏備用。
1.2 試驗方法
1.2.1 磨粉 利用實驗磨粉機(型號LRMM 8040-3-D,江蘇無錫錫糧機械制造有限公司)磨粉,出粉率約為60%。
1.2.2 粉質參數測定 利用粉質儀(型號810101,德國布拉本德公司)測定面團形成時間、穩定時間、弱化度、粉質質量指數等粉質參數,每份材料測3次平行。
1.3 數據分析
利用DPS 7.05軟件進行數據分析處理,遺傳分析參照孔繁玲和劉來福等的方法[18-19]。
2 結果與分析
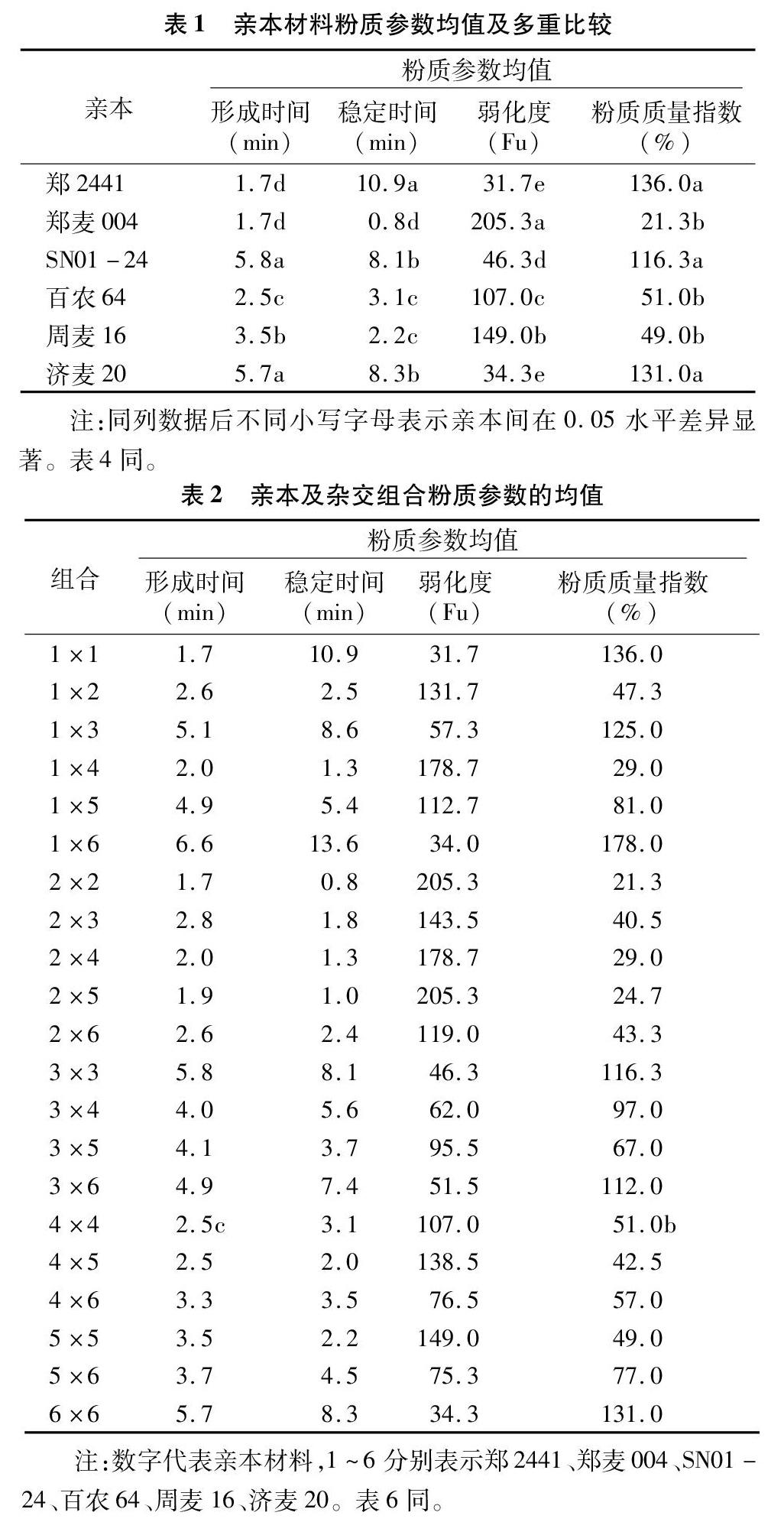
2.1 親本及F1面團粉質參數的平均表現
所有粉質參數在6個親本材料間的差異均達到顯著水平。
所有雜交組合的形成時間均介于鄭2441和SN01-24之間,穩定時間介于鄭2441和鄭麥004之間,粉質質量指數介于鄭麥004和鄭2441之間,主要表現規律為親本值高,后代值也較高。而弱化度在組合中的數值介于鄭麥004和鄭2441之間,表現為親本的值越低其F1的值越低(表2)。整體來看,未發現超親優勢現象。
2.2 親本及F1面團粉質參數配合力的方差分析
由表3可知,4個粉質參數的一般配合力方差均達到了顯著或極顯著水平,特殊配合力方差除弱化度和粉質質量指數外,其余參數也達到顯著或極顯著水平,表明形成時間和穩定時間同時受加性和非加性效應的影響,而弱化度和粉質質量指數主要受加性效應影響。
2.3 面團粉質參數的一般配合力效應分析及親本評價
基因加性效應用一般配合力來衡量,能夠穩定遺傳給后代,其一般配合力效應值與性狀遺傳成正比。一般配合力高的品種(系),將某一性狀傳遞給后代的能力強,所以親本的一般配合力在育種中有非常重要的意義。
一般配合力在6個親本中既有正向效應,又有負向效應。同一親本在不同參數的一般配合力效應差異較大,同一參數不同親本的一般配合力效應差異也很大(表4)。親本鄭2441、SN01-24和濟麥20形成時間的一般配合力效應值表現為正向效應,表明與其組配的后代形成時間可能會長;而鄭麥004、百農64和周麥16形成時間表現為負向效應,表明與其組配的后代形成時間可能會短。鄭2441和濟麥20穩定時間的一般配合力效應為正向效應,兩者之間差異不顯著,其余親本則為負向效應。鄭2441、SN01-24和濟麥20的粉質質量指數的一般效應值為正向效應,且三者之間差異不顯著,其余親本為負向效應。綜合上述,如果選擇優質強筋的小麥時,鄭2441和濟麥20這2個親本的一般配合力較好,可以顯著提高后代的面團粉質參數。
表5為6個親本面團粉質參數GCA效應的相對位次,不同品種間面團粉質參數GCA存在明顯差別,表現最為突出的是鄭2441和濟麥20。其中,鄭2441除形成時間外,其余,參數的GCA效應都是第1位,總體位次也排第1;濟麥20為第2位,兩者面團粉質參數的GCA均較好。鄭麥004的GCA效應較低。
2.4 雜交組合面團粉質參數特殊配合力的效應分析
由表6看出,不同組合同一參數的特殊配合力差異較大。其中,形成時間特殊配合力效應值的變幅為-0.53~1.48,穩定時間為-1.72~3.16,弱化度為-14.84~13.50,粉質質量指數為-17.00~38.53。形成時間特殊配合力表現為正值的組合共有7個,表現為負值的組合有8個,其中,正值較大的組合為鄭2441×濟麥20,負值較大的組合為鄭麥004×濟麥20和周麥16×濟麥20。濟麥20參與的組合正值和負值均較大,雖然其一般配合力具有正向作用,但在一些組合中特殊配合力的負向效應也很明顯,說明某些組合非加性效應占主導作用。在15個組合中,穩定時間表現正值的組合有5個,表現負值的組合有10個。其中,正值較大的組合為鄭2441×濟麥20,負值較大的組合為鄭2441×鄭麥004和鄭2441×百農64。鄭2441參與的組合既有較大的正值又有較大的負值,雖然鄭2441的一般配合力具有正向作用,但在一些組合中特殊配合力的負向效應也很明顯,說明某些組合非加性效應占主導作用。
粉質質量指數表現為正值的組合有6個,正值較大的組合為鄭2441×濟麥20和SN01-24×百農64, 負值較大的組合為鄭2441×鄭麥004和百農64×濟麥20。綜合上述參數的特殊配合力來看, 鄭2441×濟麥20是強優勢組合,且組合中有一般配合力較高的親本。
2.5 面團粉質參數遺傳參數估計
在隨機模型下對6個親本及15個雜交組合的遺傳參數進行估計(表7),4個粉質參數的廣義遺傳力和狹義遺傳力均較高,說明粉質參數的遺傳同時受加性和非加性效應控制;加性方差明顯大于顯性方差,說明主要以加性效應為主。因此,面團粉質性狀適合在育種早期世代進行選擇。而環境方差和總遺傳方差較小,說明粉質參數受環境影響相對較小。
3 討論與結論
親本選配和強優勢組合篩選是雜交育種的關鍵,在小麥品質育種中也尤為重要。本試驗對小麥面團粉質參數進行配合力及遺傳參數估計,結果表明,4個粉質參數的一般配合力方差均達到了顯著或極顯著水平,特殊配合力方差參數也達到了顯著或極顯著水平(弱化度和粉質質量指數除外);4個粉質參數的廣義遺傳力均較高,均大于79%,且狹義遺傳力也高,另外,加性方差明顯大于顯性方差,表明弱化度和粉質質量指數主要受加性效應影響,而形成時間和穩定時間同時受加性和非加性效應控制,且以加性效應為主。綜合一般配合力和特殊配合力來看,鄭2441×濟麥20是強優勢組合,組合中有一般配合力較高的親本,小麥品種選育時可著重考慮該組合。這與劉建軍等的結果[20-23]基本一致。詹克慧等運用Hayman遺傳效應分析方法對面團粉質特性進行遺傳分析,結果發現穩定時間為加-顯性模型,且為部分顯性,而形成時間、斷裂時間、弱化度存在上位性效應[24],本研究結果與之存在一定差異, 原因可能是分析方法以及試驗材料等不同造成的。小麥面團粉質參數的遺傳規律需要多年多點的數據進行研究,本試驗僅選用1個環境,今后還需加大研究力度。
參考文獻:
[1]李寧波,王曉曦,于 磊,等. 面團流變學特性及其在食品加工中的應用[J]. 食品科技,2008(8):35-38.
[2]呂軍倉. 面團流變學及其在面制品中的應用[J]. 糧油加工與食品機械,2006(2):66-68.
[3]劉建軍,何中虎,趙振東,等. 小麥面條加工品質研究進展[J]. 麥類作物學報,2001,21(2):81-84.
[4]孫 輝,吳存榮,李 玥,等. 我國小麥標準體系現狀及發展方向[J]. 糧食與飼料工業,2010(4):13-17.
[5]李蓓蓓,王鳳成,齊兵建,等. 國產小麥粉與酥性餅干品質的相關性研究[J]. 現代面粉工業,2011,25(4):36-43.
[6]吳同彥,楊 亮,謝令琴,等. 小麥主要農藝性狀配合力分析[J]. 河北農業大學學報,2009,32(5):12-17.
[7]Dagustu N. Genetic analysis of grain yield per spike and some agronomic traits in diallel crosses of bread wheat (Triticum aestivum L.)[J]. Turkish Journal of Agriculture and Forestry,2008,32(4):249-258.
[8]Inamullah,Ahmad H,Mohannad F,et al. Diallel analysis of the inheritance pattern of agronomic traits of bread wheat[J]. Pakistan Journal of Botany,2006,38(4):1169-1175.
[9]王翠玲,高海濤,王書子,等. 冬小麥品質與產量性狀的遺傳分析[J]. 麥類作物學報,2003,23(3):26-28.
[10]盧 超,高明博,焦小鐘,等. 幾個小麥親本主要農藝性狀的配合力評價及遺傳力分析[J]. 麥類作物學報,2010,30(6):1023-1028.
[11]Chung O K,Ohm J B,Caley M S,et al. Prediction of baking characteristics of hard winter wheat flours using computer-analyzed? mixogram parameters[J]. Cereal Chemistry,2001,78(4):493-497.
[12]Zhu J,Huang S,Khan K. Relationship of protein quantity,quality and dough properties with Chinese steamed bread quality[J]. Journal of Cereal Science,2001,33(2):205-212.
[13]田紀春,胡瑞波,陳建省,等. 小麥粉面團穩定時間的變化及其穩定性分析[J]. 中國農業科學,2005,38(11):22-29.
[14]余中偉,向長萍. 雙列雜交在作物遺傳育種中的應用[J]. 長江蔬菜,2008(8):1-4.
[15]袁有祿,張天真,郭旺珍,等. 棉花優異纖維品質性狀的雙列雜交分析[J]. 遺傳學報,2005,32(1):79-85.
[16]方繼友. 玉米自交系產量性狀和營養成分的配合力及遺傳參數分析[D]. 北京:中國農業大學,2005.
[17]鄭 軼,楊德衛,涂詩航,等. 我國雜交水稻相關性狀配合力、遺傳力研究進展[J]. 福建農業學報,2014,29(9):926-933.
[18]孔繁玲. 植物數量遺傳學[M]. 北京:中國農業大學出版社,2006.
[19]劉來福,毛盛賢,黃遠樟,等. 作物數量遺傳[M]. 北京:農業出版社,1984.
[20]劉建軍,趙振東,董進英,等. 冬小麥面團流變學特性的雜種優勢和配合力分析[J]. 作物學報,1996,22(5):577-582.
[21]張 利,田笑明,曹連莆,等. 冬小麥主要品質性狀配合力和雜種優勢研究[J]. 新疆農業科學,2005,42(2):65-69.
[22]張 利,田笑明,曹連莆,等. 冬小麥面粉品質性狀配合力和雜種優勢研究[J]. 麥類作物學報,2005,25(4):24-28.
[23]張 利. 冬小麥主要品質性狀和產量性狀雜種優勢及其相關性的研究[D]. 石河子:石河子大學,2005.
[24]詹克慧,王福亭,崔黨群,等. 小麥面粉品質性狀的遺傳分析[J]. 河南農業大學學報,1995,29(4):348-352.
