濃香型白酒酒醅風(fēng)味物質(zhì)和微生物菌群結(jié)構(gòu)隨發(fā)酵時(shí)間的變化規(guī)律
2019-11-14 06:45:12高江婧任聰劉國(guó)英葛向陽(yáng)王慧琳葉方平徐巖
食品與發(fā)酵工業(yè) 2019年20期
高江婧,任聰,劉國(guó)英,葛向陽(yáng),王慧琳,葉方平,徐巖*
1(江南大學(xué) 生物工程學(xué)院,釀造微生物學(xué)與應(yīng)用酶學(xué)研究室,江蘇 無(wú)錫,214122) 2(安徽古井貢酒股份有限公司,安徽 亳州,236820) 3(教育部工業(yè)生物技術(shù)重點(diǎn)實(shí)驗(yàn)室(江南大學(xué)),江蘇 無(wú)錫,214122)
在濃香型白酒釀造生產(chǎn)實(shí)踐中,通常采用“雙輪底發(fā)酵工藝”來(lái)提高酒質(zhì),雙輪底糟發(fā)酵實(shí)質(zhì)是延長(zhǎng)發(fā)酵期的一種工藝方法[1-2]。受夏季氣溫過(guò)高因素的影響,酒醅發(fā)酵溫度較難控制,很多名優(yōu)酒廠采取暫停生產(chǎn)措施以安全度夏,這勢(shì)必會(huì)造成酒醅發(fā)酵周期的延長(zhǎng)。而長(zhǎng)期的生產(chǎn)經(jīng)驗(yàn)表明,延長(zhǎng)酒醅發(fā)酵時(shí)間后,所釀造出的濃香型原酒品質(zhì)更佳。
近年來(lái),國(guó)內(nèi)學(xué)者逐步重視對(duì)濃香型白酒酒醅中的微生物菌群演替規(guī)律進(jìn)行解析。多數(shù)研究表明,在發(fā)酵過(guò)程中,酒醅中的乳酸菌成為絕對(duì)優(yōu)勢(shì)菌群,乳酸菌在發(fā)酵初期快速繁殖,發(fā)酵中后期相對(duì)豐度呈緩慢上升趨勢(shì)[3-5];劉凡等[6]通過(guò)宏基因組測(cè)序解析了濃香型白酒發(fā)酵過(guò)程中的微生物與主要有機(jī)酸合成的關(guān)系,發(fā)現(xiàn)窖內(nèi)發(fā)酵初期(0~14 d)與主要有機(jī)酸(乳酸、乙酸、丁酸和己酸)合成相關(guān)的微生物數(shù)量顯著高于發(fā)酵后期(15~60 d)。上述有關(guān)酒醅微生物菌群的研究均針對(duì)的是2個(gè)月的常用發(fā)酵周期,尚未揭示出延長(zhǎng)發(fā)酵時(shí)間后窖內(nèi)酒醅微生物菌群和風(fēng)味物質(zhì)的變化規(guī)律。
窖泥在濃香型白酒生產(chǎn)中占據(jù)重要地位,隨著測(cè)序技術(shù)的快速發(fā)展,目前對(duì)窖泥的微生物菌群結(jié)構(gòu)認(rèn)識(shí)逐步清晰,研究發(fā)現(xiàn)地域、窖齡、窖內(nèi)分布位置、窖泥質(zhì)量均對(duì)窖泥微生物菌群結(jié)構(gòu)具有較大的影響,如新窖泥以乳桿菌為主,而老窖泥以梭菌綱、擬桿菌綱和產(chǎn)甲烷菌為主[7-12]。王雪山等[13]分析了濃香型酒醅微生物的來(lái)源,發(fā)現(xiàn)大曲是酒醅中好氧和兼性厭氧菌的主要來(lái)源(占比74%),而窖泥是酒醅厭氧菌群的主要來(lái)源(占比14%),但窖泥微生物菌群對(duì)酒醅微生物菌動(dòng)態(tài)變化的影響卻鮮有研究。另一方面,濃香型白酒發(fā)酵窖池深達(dá)2 m,對(duì)發(fā)酵過(guò)程酒醅取樣,尤其是對(duì)底層酒醅取樣困難,發(fā)酵后期較多的黃水也對(duì)酒醅取樣造成困難,目前尚缺少對(duì)濃香型白酒發(fā)酵窖池底層酒醅中微生物與風(fēng)味物質(zhì)產(chǎn)生動(dòng)態(tài)規(guī)律的長(zhǎng)發(fā)酵周期追蹤研究。本文跟蹤了發(fā)酵周期長(zhǎng)達(dá)5個(gè)月的窖池,并重點(diǎn)關(guān)注了窖池下層酒醅中的主要風(fēng)味物質(zhì)和微生物菌群的變化規(guī)律,為解析延長(zhǎng)發(fā)酵時(shí)間與原酒品質(zhì)關(guān)系提供基礎(chǔ)數(shù)據(jù)支撐。
1 材料與方法
1.1 材料
在濃香型白酒釀造車間,選取正常生產(chǎn)班組的2口老窖池,窖齡均為27年。采用自制酒醅取樣器分別取樣不同發(fā)酵時(shí)間點(diǎn)的酒醅樣本,連續(xù)跟蹤取樣直至156 d發(fā)酵結(jié)束。本文取樣的酒醅樣本均為窖池下部的酒醅。發(fā)酵結(jié)束后,采用五點(diǎn)取樣法取窖泥100 g并混合均勻。酒醅和窖泥樣本均凍存于-20℃冰箱待用。
1.2 主要試劑
己酸乙酯、乳酸乙酯、乙酸、乳酸、丁酸、己酸、叔戊醇、乙酸正戊酯和2-乙基丁酸等標(biāo)準(zhǔn)品(色譜純):購(gòu)于美國(guó)Sigma-Aldrich試劑公司;無(wú)水乙醇(色譜純):購(gòu)于上海安譜公司;DNA提取試劑盒:購(gòu)于美國(guó)Qiagen公司。
1.3 揮發(fā)性組分分析
稱取酒醅(窖泥)樣本10 g,加入60%的無(wú)水乙醇20 mL渦旋混勻,浸提30 min后,10 000 r/min離心10 min,取上清液過(guò)膜,并加入混合內(nèi)標(biāo)(叔戊醇、乙酸正戊酯和2-乙基丁酸)后進(jìn)行氣相色譜分析。色譜柱為CP-wax57CB毛細(xì)管柱(50m×0.25mm×0.25μm),氣相進(jìn)樣口溫度為230℃,檢測(cè)器溫度為230℃,載氣為高純氮,流速為1.0 mL/min,進(jìn)樣量為1 μL,分流比為30∶1。程序升溫條件為:初始溫度35℃,保持0 min,以4 ℃/min升到60℃,保持4 min,再以6 ℃/min升到195 ℃,保持20 min。
1.4 乳酸分析
酒醅(窖泥)樣本用超純水浸泡離心,并經(jīng)適當(dāng)稀釋后,采用超高效液相色譜法[14]測(cè)試上清液的乳酸含量。色譜柱為Waters Acquity UPLC HSS T3(100 mm×2.1 mm×1.8 μm),流動(dòng)相為0.02 mol/L KH2PO4,流速為0.1 mL/min,進(jìn)樣量為1 μL,二極管陣列檢測(cè)器,檢測(cè)波長(zhǎng)為208 nm。
1.5 酒醅和窖泥微生物DNA的提取
對(duì)于酒醅樣本,采用滅菌的PBS緩沖液多次提取酒醅中的微生物細(xì)胞,以去除雜質(zhì),收集細(xì)胞沉淀,然后采用DNeasy UltraClean Microbial Kit提取酒醅微生物總基因組DNA。窖泥微生物總基因組DNA利用DNeasy Power-Soil Kit進(jìn)行提取,提取方法按照試劑公司提供的實(shí)驗(yàn)操作指南進(jìn)行。利用瓊脂糖電泳對(duì)提取的微生物DNA進(jìn)行質(zhì)量檢測(cè),合格后進(jìn)行建庫(kù)、測(cè)序。
1.6 高通量測(cè)序與數(shù)據(jù)分析
建庫(kù)與測(cè)序由上海美吉生物技術(shù)有限公司完成。高通量測(cè)序針對(duì)細(xì)菌16S rRNA基因V3-V4區(qū)域,擴(kuò)增引物為338F(5′-ACTCCTACGGGAGGCAGCAG-3′)和 806R(5′-GGACTACHVGGGTWTCTAAT -3′)。測(cè)序平臺(tái)為Illumina MiSeq。測(cè)序原始序列采用FLASH和Trimmomatic軟件進(jìn)行拼接及質(zhì)控過(guò)濾,序列的質(zhì)檢標(biāo)準(zhǔn)為:(1)去除序列尾部質(zhì)量值20以下的堿基,去除質(zhì)控后50 bp以下的序列;(2)引物最大錯(cuò)配數(shù)為2,barcode允許的錯(cuò)配數(shù)為0,去除模糊堿基;(3)最小overlap序列長(zhǎng)度為10 bp,且拼接序列的overlap區(qū)允許最大錯(cuò)配比率為0.2。采用Uchime軟件去除嵌合體序列。使用Usearch(vsesion 7.0 http://drive5.com/uparse/)將序列按照97%相似度聚類,進(jìn)行OTU(operational taxonomic units,操作分類單元)劃分,提取代表序列,得到OTU表。將細(xì)菌OTU代表序列與Silva(Release132,http://www.arb-silva.de)數(shù)據(jù)庫(kù)比對(duì),進(jìn)行分類學(xué)注釋。細(xì)菌菌群熱圖(heatmap)和冗余分析(redundancy analysis,RDA)通過(guò)R語(yǔ)言(vegan包)分析。
2 結(jié)果與分析
2.1 酒醅主要風(fēng)味物質(zhì)隨發(fā)酵時(shí)間的變化趨勢(shì)
在濃香型白酒生產(chǎn)過(guò)程中,靠近窖池底部的下層酒醅質(zhì)量最佳,一般用于生產(chǎn)優(yōu)質(zhì)濃香型原酒。對(duì)窖池下層酒醅的4種主要有機(jī)酸進(jìn)行測(cè)試分析,結(jié)果表明乳酸含量在總酸中占有絕對(duì)優(yōu)勢(shì)(>85%),其次是乙酸、己酸和丁酸。在整個(gè)156 d長(zhǎng)的發(fā)酵周期內(nèi),乳酸含量在發(fā)酵初期0 ~14 d快速增加,之后乳酸含量增速變緩,在發(fā)酵120 d后乳酸含量有略微下降的趨勢(shì)(圖1-A)。乙酸則在發(fā)酵初期0~21 d快速增加,之后略有緩慢增加趨勢(shì)。而丁酸和己酸的含量在發(fā)酵50 d后持續(xù)增加,直至發(fā)酵結(jié)束(圖1-B)。由此可知,對(duì)于較短的發(fā)酵周期(60 d以內(nèi)),酒醅中的丁酸、己酸并未大量積累,這與相關(guān)文獻(xiàn)的報(bào)道基本一致[6]。
與此同時(shí),酒醅中的乳酸乙酯和乙酸乙酯兩種風(fēng)味組分與乳酸、乙酸的變化趨勢(shì)基本一致,在發(fā)酵初期,乳酸乙酯和乙酸乙酯呈現(xiàn)顯著增加趨勢(shì),20 d后則逐漸趨于平穩(wěn),而酒醅中己酸乙酯的含量則在長(zhǎng)期發(fā)酵的后期迅速增加(圖1-C),表明在濃香型白酒釀造工藝中,延長(zhǎng)發(fā)酵周期,可以顯著提高酒醅中己酸乙酯的含量。己酸乙酯含量的提高,相應(yīng)提高了己酸乙酯與乙酸乙酯的比值,因而延長(zhǎng)發(fā)酵時(shí)間,也有利于促進(jìn)白酒中酯類物質(zhì)的平衡。
2.2 酒醅原核微生物隨發(fā)酵時(shí)間的變化趨勢(shì)
為進(jìn)一步認(rèn)識(shí)窖池下層酒醅在長(zhǎng)期發(fā)酵過(guò)程中的微生物變化規(guī)律,對(duì)不同發(fā)酵時(shí)間的酒醅細(xì)菌菌群結(jié)構(gòu)進(jìn)行分析。酒醅16S rRNA基因測(cè)序表明,酒醅中的細(xì)菌主要以芽孢桿菌綱的乳桿菌科(Lactobacillaceae)為主,乳桿菌在整個(gè)發(fā)酵過(guò)程中占有絕對(duì)優(yōu)勢(shì),因而乳桿菌的代謝產(chǎn)物乳酸在總酸中也占有絕對(duì)優(yōu)勢(shì)。在發(fā)酵14 d時(shí),乳桿菌科的相對(duì)豐度達(dá)到99%以上,這與發(fā)酵初期酒醅中的乳酸含量急劇增長(zhǎng)相對(duì)應(yīng),表明乳桿菌科在發(fā)酵初期即大量繁殖,代謝產(chǎn)生高含量的乳酸。但隨著發(fā)酵時(shí)間的延長(zhǎng),乳桿菌科的相對(duì)豐度逐漸下降至85%左右(圖2)。相反,梭菌綱微生物的相對(duì)豐度則逐漸增加至8.94%。
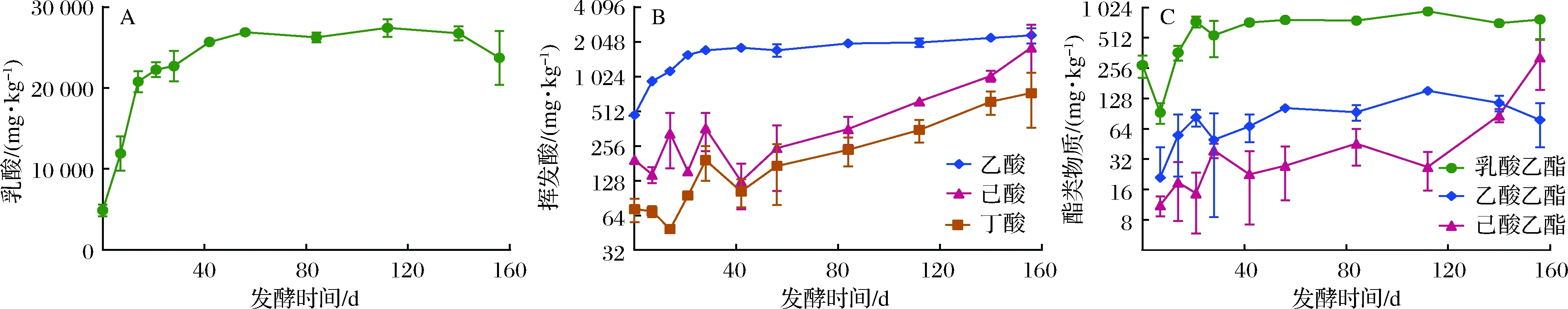
A-乳酸;B-揮發(fā)酸;C-酯類物質(zhì)圖1 酒醅發(fā)酵過(guò)程中主要風(fēng)味物質(zhì)含量變化Fig.1 Dynamics of key flavor compounds in fermented grains during fermentation

A-綱水平;B-科水平圖2 酒醅細(xì)菌菌群隨發(fā)酵時(shí)間變化規(guī)律Fig.2 Dynamic of bacterial community in fermented grains during fermentation注:圖中數(shù)據(jù)為相同發(fā)酵時(shí)間點(diǎn)酒醅樣本測(cè)序結(jié)果的平均值。下同。
由于乳桿菌科在整個(gè)發(fā)酵過(guò)程中占有絕對(duì)優(yōu)勢(shì),其相對(duì)豐度均大于80%,當(dāng)扣除乳桿菌科后,可觀測(cè)到酒醅中梭菌綱微生物的相對(duì)豐度隨發(fā)酵時(shí)間的延長(zhǎng)呈現(xiàn)出顯著遞增趨勢(shì)。在科水平上,梭菌綱的瘤胃菌科(Ruminococcaceae)和梭菌科(Clostridiaceae_1)的相對(duì)豐度隨發(fā)酵時(shí)間的延長(zhǎng)顯著增加。除乳桿菌科的相對(duì)豐度隨發(fā)酵時(shí)間延長(zhǎng)下降外,r-變形菌綱的腸桿菌科(Enterobacteriaceae)也隨發(fā)酵時(shí)間的延長(zhǎng)而顯著降低(圖3)。
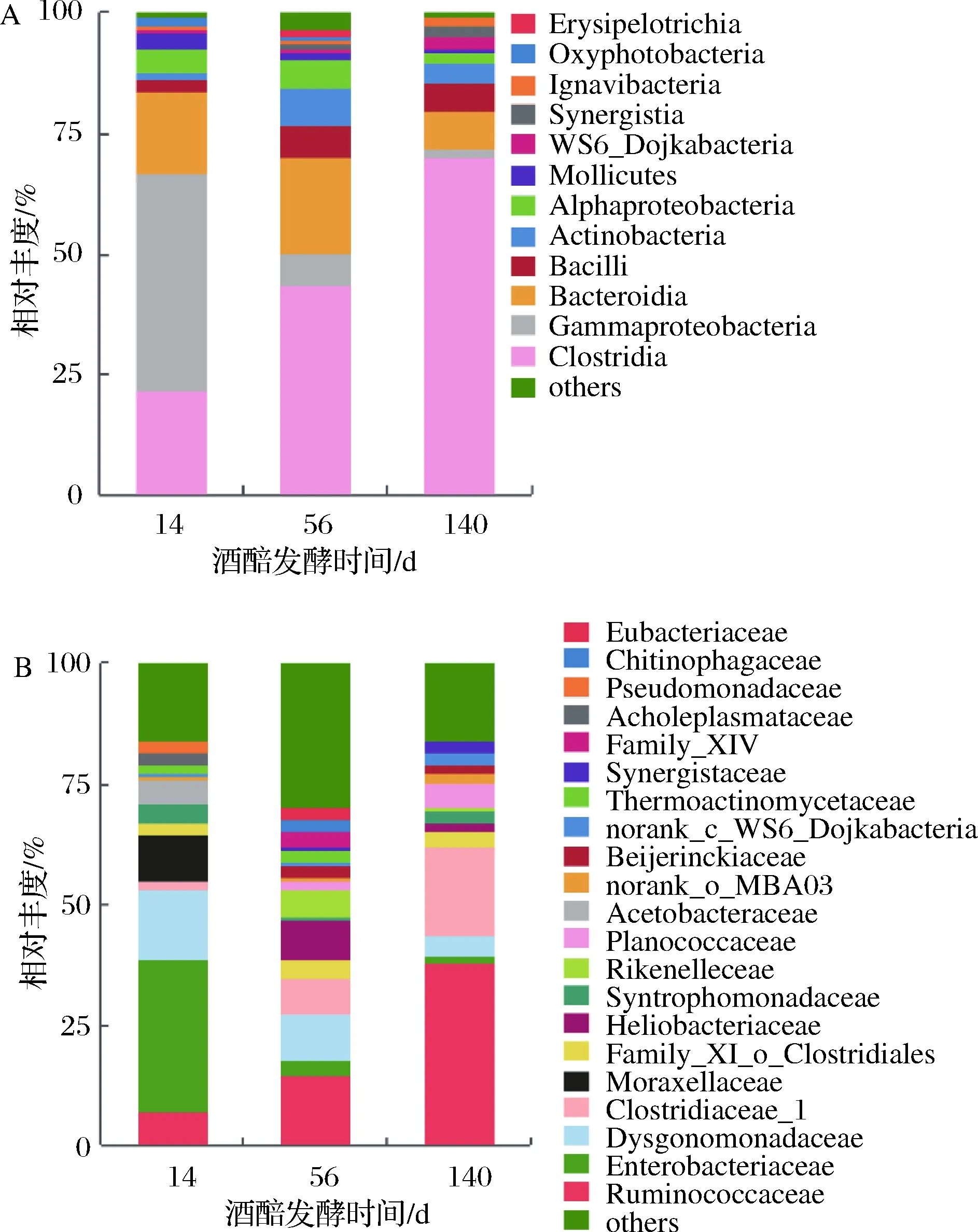
A-綱水平;B-科水平圖3 酒醅扣除乳桿菌科后的細(xì)菌菌群隨發(fā)酵時(shí)間變化規(guī)律Fig.3 Dynamic of bacterial community expect for Lactobacillaceae in fermented grains during fermentation
2.3 不同發(fā)酵時(shí)間酒醅微生物與窖泥微生物的相關(guān)性
由于窖泥是酒醅厭氧微生物菌群的重要來(lái)源,王雪山等通過(guò)微生物來(lái)源(SourceTracker)分析酒醅微生物的來(lái)源,發(fā)現(xiàn)窖泥來(lái)源厭氧菌占到酒醅原核菌群的14%[13]。將不同發(fā)酵時(shí)間的酒醅微生物與窖泥微生物的種類分別進(jìn)行比較,發(fā)現(xiàn)隨著發(fā)酵時(shí)間的延長(zhǎng),酒醅與窖泥中檢測(cè)到的共有微生物種類逐漸增多,在科水平上,從發(fā)酵14 d的28個(gè)共有微生物增加至140 d的82個(gè)共有微生物(圖4)。
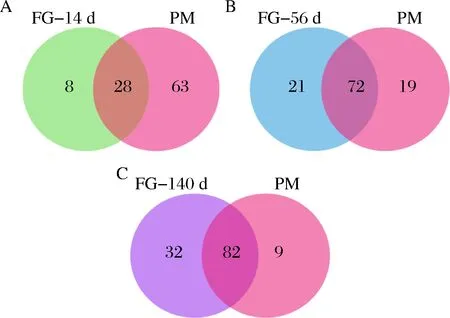
A-發(fā)酵14 d;B-發(fā)酵56 d;C-發(fā)酵140 d圖4 不同發(fā)酵時(shí)間酒醅與窖泥共有微生物(科水平)比較分析Fig.4 Comparison of communal microorganisms (at family level) between pit mud and fermentedgrains at different fermentation time注:FG-14d表示發(fā)酵14 d的酒醅;FG-56d表示發(fā)酵56 d的酒醅;FG-140 d表示發(fā)酵140 d的酒醅;PM表示窖泥。下同。
通過(guò)比較窖泥和不同發(fā)酵時(shí)間酒醅的微生物差異,進(jìn)一步發(fā)現(xiàn),隨發(fā)酵時(shí)間在酒醅中相對(duì)豐度逐漸升高的微生物,如瘤胃菌科(Ruminococcaceae)、Dysgonomonadaceae、梭菌科(Clostridiaceae_1),這些微生物也是窖泥中的高豐度微生物,它們?cè)诮涯嘀械钠骄鄬?duì)豐度分別為24.79%、22.35%、13.01%(圖5)。表明在長(zhǎng)期發(fā)酵過(guò)程中,窖泥為酒醅持續(xù)不斷提供其主體的厭氧微生物菌群。值得說(shuō)明的是,瘤胃菌只是細(xì)菌分類學(xué)名稱,最初命名源于從牛瘤胃中發(fā)現(xiàn)的能產(chǎn)生丁酸的某種新類型厭氧菌,而窖泥中的瘤胃菌屬于該科下的新成員,與定義該科源自于瘤胃的細(xì)菌并不是同屬細(xì)菌,它們只是在科水平上的親緣關(guān)系上較為接近。
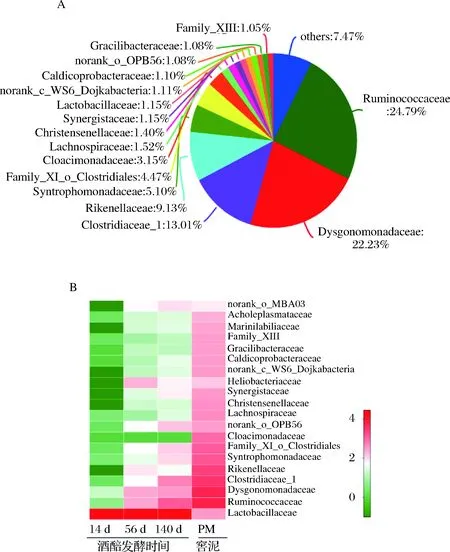
A-菌群結(jié)構(gòu);B-變化規(guī)律圖5 窖泥菌群結(jié)構(gòu)和對(duì)應(yīng)的酒醅菌群隨發(fā)酵時(shí)間變化規(guī)律Fig.5 The bacterial community of pit mud and dynamic of bacterial community in fermented grains during long-termfermentation注:數(shù)據(jù)為樣本測(cè)序結(jié)果的平均值,菌群為科水平。
此外,窖泥中平均相對(duì)豐度較高的一些微生物,如Rikenellaceae (9.13%)和Cloacimonadaceae(3.15%),它們?cè)诰契械南鄬?duì)豐度并未同比例增加,后者甚至在酒醅中檢測(cè)不到;而窖泥中平均相對(duì)豐度較低的Heliobacteriaceae(0.47%)在酒醅中的相對(duì)豐度卻較高。雖然窖池下部酒醅在發(fā)酵過(guò)程中呈現(xiàn)半固態(tài)(由于原料水解和微生物的代謝活動(dòng),酒醅中的水分含量較高),窖泥微生物可以通過(guò)遷移或擴(kuò)散進(jìn)入酒醅,但本文研究結(jié)果表明,從窖泥遷移至酒醅中的微生物并非是同比例增加的,酒醅的高酸高醇環(huán)境也可能影響窖泥微生物向酒醅的遷移速率和生長(zhǎng)代謝。而與此相對(duì)的是,酒醅中雖富含有80%以上高豐度的乳桿菌,但其在窖泥中的相對(duì)豐度卻僅有1%左右,表明即使在長(zhǎng)期的發(fā)酵過(guò)程中,酒醅中占有絕對(duì)優(yōu)勢(shì)的乳桿菌并未能大量入侵到窖泥微生物體系中,正常窖泥微生物菌群具有較好的魯棒性,能夠抵抗酒醅中乳酸菌的入侵。
2.4 風(fēng)味物質(zhì)與微生物的相關(guān)性分析
上述分析比較了不同發(fā)酵時(shí)間酒醅和發(fā)酵終點(diǎn)窖泥細(xì)菌菌群結(jié)構(gòu)的差異,發(fā)現(xiàn)隨發(fā)酵時(shí)間的延長(zhǎng),相對(duì)豐度逐漸增高的物種亦是窖泥中的主體功能菌群。為進(jìn)一步分析這些微生物與發(fā)酵過(guò)程中的風(fēng)味物質(zhì)生成是否相關(guān),我們進(jìn)行了冗余分析(RDA)(圖6)。結(jié)果表明,乳桿菌科的相對(duì)豐度與乳酸含量呈強(qiáng)正相關(guān),而丁酸、乙酸、己酸含量則與梭菌科、瘤胃菌科、Dysgonomondaceae的相對(duì)含量呈現(xiàn)正相關(guān)關(guān)系。因此,推測(cè)這些微生物對(duì)發(fā)酵后期己酸、丁酸、乙酸的合成具有重要貢獻(xiàn)。
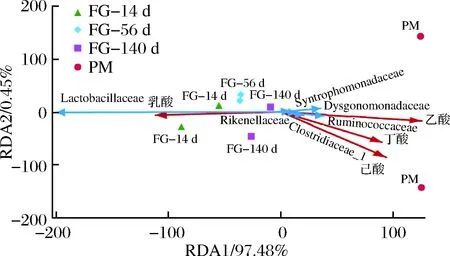
圖6 微生物與風(fēng)味物質(zhì)相關(guān)性分析Fig.6 Correlation between microbes and flavor compounds
3 討論
本文對(duì)長(zhǎng)達(dá)5個(gè)月發(fā)酵周期的窖池下層酒醅進(jìn)行分析,發(fā)現(xiàn)在發(fā)酵初期(0~14 d),由于乳桿菌的大量繁殖,酒醅乳酸含量迅速增加,此后乳桿菌在整個(gè)發(fā)酵過(guò)程中占有絕對(duì)優(yōu)勢(shì)。乳桿菌在發(fā)酵初期即大量繁殖,其代謝生成的高濃度乳酸抑制了無(wú)耐酸性能的微生物或有害菌的生長(zhǎng),避免了發(fā)酵過(guò)程受到這些微生物的污染。相對(duì)占絕對(duì)優(yōu)勢(shì)的乳桿菌而言,梭菌綱微生物占比較小,雖然梭菌綱微生物的相對(duì)豐度隨發(fā)酵時(shí)間呈現(xiàn)顯著遞增趨勢(shì),但即使到發(fā)酵140 d時(shí),其平均相對(duì)豐度也僅占總細(xì)菌的8.94%。雖然酒醅中的梭菌綱微生物在數(shù)量上不占優(yōu)勢(shì),但其對(duì)酒醅風(fēng)味物質(zhì)貢獻(xiàn)卻巨大。
任聰?shù)萚15]通過(guò)對(duì)新、老窖泥的微生物菌群結(jié)構(gòu)和進(jìn)化關(guān)系,發(fā)現(xiàn)窖泥中的主體己酸菌為梭菌綱瘤胃菌科的己酸菌屬(Caproiciproducens)微生物,且以乳酸和葡萄糖利用型己酸菌為主。本文通過(guò)跟蹤長(zhǎng)達(dá)5個(gè)月的發(fā)酵過(guò)程中窖池下層酒醅微生物的變化規(guī)律,發(fā)現(xiàn)隨著發(fā)酵時(shí)間的延長(zhǎng),梭菌綱瘤胃菌科的微生物在酒醅中的相對(duì)豐度逐漸增加,并觀測(cè)到在長(zhǎng)期發(fā)酵的后期(120 d后),酒醅中的乳酸含量亦有略微的下降趨勢(shì),而丁酸、己酸含量則隨發(fā)酵期的延長(zhǎng)持續(xù)增加。推測(cè)窖泥中的主體產(chǎn)己酸菌進(jìn)入酒醅后,可將乳酸底物逐漸轉(zhuǎn)化為丁酸和己酸,從而為酯類物質(zhì)的合成不斷提供充足的前體物質(zhì)。
對(duì)酒醅菌群和窖泥菌群比較可知,酒醅中占有絕對(duì)優(yōu)勢(shì)的乳桿菌在窖泥中的相對(duì)豐度卻較低,表明即使在長(zhǎng)達(dá)5個(gè)月的發(fā)酵過(guò)程中,酒醅中占有絕對(duì)優(yōu)勢(shì)的乳桿菌并未能入侵到窖泥微生物體系中。有研究表明,新窖池的窖泥以高含量的乳桿菌為主要特征[7, 15],新窖池的窖泥由于厭氧菌群物種多樣性低,尚未形成穩(wěn)定的菌群結(jié)構(gòu),推測(cè)在長(zhǎng)時(shí)間的發(fā)酵過(guò)程中,新窖池酒醅中的乳桿菌可能會(huì)入侵至窖泥體系中。本文所取樣的酒醅和窖泥均為老窖池的樣本,老窖池的窖泥具有豐富的厭氧菌群,在長(zhǎng)時(shí)間的酒醅發(fā)酵過(guò)程中,不僅能抵御酒醅中乳桿菌的入侵,而且老窖泥中的主體微生物能不斷遷移至酒醅層中。因此酒醅的發(fā)酵期越長(zhǎng),從老窖泥遷移至酒醅中的窖泥微生物越多,代謝生成的丁酸、己酸等有機(jī)酸含量也越多。
綜上所述,對(duì)于老窖池而言,延長(zhǎng)酒醅發(fā)酵時(shí)間后,一方面,窖泥中的主體厭氧微生物不斷遷移進(jìn)入酒醅,使得酒醅中的丁酸、己酸等有機(jī)酸含量持續(xù)積累;另一方面,也為有機(jī)酸與乙醇通過(guò)酯化反應(yīng)生成酯類風(fēng)味物質(zhì)提供了更為充足的時(shí)間,因而發(fā)酵后期(120 d后),在酒醅中檢測(cè)到了己酸乙酯的大量合成。此外,己酸作為濃香型白酒重要風(fēng)味物質(zhì)己酸乙酯的前體物質(zhì),其可作為衡量濃香型原酒品質(zhì)的重要指征性指標(biāo)。本文也主要對(duì)酒醅中高含量的有機(jī)酸(乳酸、乙酸、己酸、丁酸)和相應(yīng)的酯類物質(zhì)進(jìn)行了分析,然而很多其他微量風(fēng)味物質(zhì)(如戊酸、辛酸、丁酸乙酯、戊酸乙酯、辛酸乙酯、己酸丁酯等)對(duì)濃香型白酒風(fēng)格的形成也具有重要作用[16-18],因此在長(zhǎng)期發(fā)酵過(guò)程中,酒醅中其他微量風(fēng)味物質(zhì)的變化規(guī)律仍有待深入分析。
雖然延長(zhǎng)酒醅發(fā)酵時(shí)間對(duì)提高酒質(zhì)有諸多優(yōu)勢(shì),但也存在出酒率偏低、窖池周轉(zhuǎn)率偏低等問(wèn)題。另外,現(xiàn)有對(duì)窖泥微生物的研究表明,窖泥中亦存在一些產(chǎn)異嗅味的微生物[19-21],這些微生物在長(zhǎng)期發(fā)酵過(guò)程中對(duì)原酒品質(zhì)的影響有待深入研究。
