豬Toll樣受體研究進展
2019-12-23 06:44:58王雅春
福建畜牧獸醫 2019年6期
李 鵬 王雅春
(1.中國農業大學動物科技學院 北京 100193;2.福建順鑫鑫源食品有限公司 福建南平 354004)
Toll樣受體(Toll-like receptors,TLRs)是生物進化過程中較為保守的I型跨膜蛋白,也是目前在哺乳動物中研究最多的模式識別受體,可識別細菌或病毒 的多種抗原成分,如鞭毛蛋白、脂多糖、病毒RNA等[1-3]。
TLRs作為重要的模式識別受體,一直受到研究者們的重視,隨著對豬TLRs研究的深入,很多病原的致病機制、益生菌保護作用的分子基礎得到了合理的解釋。另外,豬的TLRs基因中普遍存在非同義單核苷酸多態性(Single nucleotide polymorphism,SNP),并且主要集中在胞外編碼區,這在疫苗設計上的應用也具有重要意義[4]。
1 豬TLRs組織和細胞分布
目前,在哺乳動物細胞中發現了13種TLRs,其中10種在豬細胞上得到了克隆(TLR1-10)。根據TLRs在豬體內細胞上的分布及其接頭分子的種類,豬TLRs可分為兩大類:第一類表達于細胞表面,主要識別微生物或其特定成分,包括TLR1、TLR2、TLR4、TLR5、TLR6 和 TLR10[4];第二類分布在細胞器膜表面,主要識別核酸及其衍生物,包括TLR3、TLR7、TLR8 和 TLR9[5-6]。
在不同組織或細胞上TLRs的表達水平具有明顯差異[7-8],免疫相關的組織或細胞上的表達水平相對較高,如脾臟、腸系膜淋巴結以及與外界環境接觸的腸和氣管等粘膜組織[9]。在豬生殖系統中不同器官TLRs的表達水平也明顯不同,如TLR3和TLR5在豬睪丸、附睪、卵巢和輸卵管中表達水平較高,而TLR9的表達水平較低,在胚胎中TLR1表達水平較高[10]。另外,TLRs在細胞上的表達水平還與豬的品種、生長日齡及飼料種類有關,如大多數TLRs在藏豬細胞上的表達量顯著多于約克夏豬,且隨著豬日齡的增長其表達量也隨之增加[11];成年豬腸系膜淋巴結TLR1和TLR6、胃TLR3的表達量顯著高于新生仔豬;在12周齡的豬體內,約克夏豬和藏豬脾臟、血液、胸腺的TLR5及其脾臟的TLR6、TLR7,約克夏豬脾臟的TLR3、TLR4,藏豬扁桃體的TLR9均具有較高的表達水平[11];蘆薈多糖可以提高豬肝臟和脾中TLR2和TLR4的表達水平,這有助于提高斷奶仔豬的免疫功能[12]。這些TLRs表達差異可能與豬的抗病能力密切相關。
2 豬TLRs的進化分析
通過物種間進化分析發現,哺乳動物TLRs可大致分為6組,且豬與人的TLRs進化關系比較接近,見圖 1。 TLR1、TLR2 和 TLR6(也包含 TLR10,但其配體尚不明確)為一組,主要識別革蘭氏陽性菌的復合物,TLR1、TLR6和 TLR10與TLR2來源于同一分支[13],而 TLR1、TLR6(可能包含 TLR10)與 TLR2形成二聚體發揮作用[14-15]。與其它TLRs相比,TLR3、TLR4、TLR5在進化上比較特殊,分別各為一組,TLR4主要識別革蘭氏陰性菌的脂多糖成分,TLR3、TLR5分別識別不同病毒或細菌的抗原組分[16]。TLR7、TLR8和TLR9分為一組,主要分布在宿主細胞的細胞器膜上,在病毒成分的識別上發揮主要作用。鼠TLR11、TLR12和TLR13在人和偶蹄獸動物中還沒有發現,其中TLR11主要識別非病原菌,TLR12和TLR13還沒找到相應的配體[17]。
3 豬TLRs各家族成員研究現狀
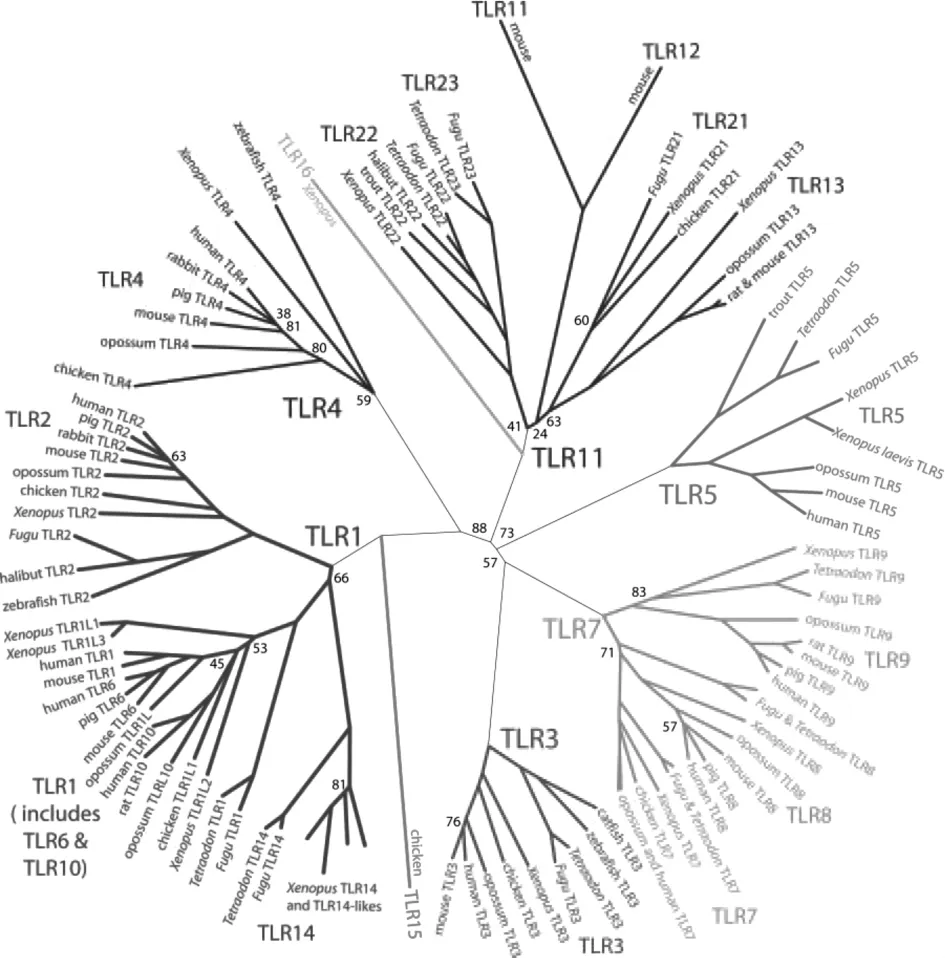
圖1 哺乳動物TLRs進化樹
入侵的細菌、病毒等微生物或其特定成分被TLRs識別后,TLRs下游信號通路將被激活,進而上調促炎細胞因子或干擾素的表達[4](見圖2)。髓樣分化因子(myeloid differentiation factor88,MyD88)是TLR信號通路中一個重要的接頭分子,根據TLR信號通路對MyD88的依賴性,TLRs信號通路分為兩種:MyD88-dependent和 MyD88-independent signaling pathways,在傳遞上游信息和疾病發生發展中具有重要的作用[13]。
3.1 TLR1、TLR2和TLR6 它們在進化關系上比較接近,且TLR2與TLR1或TLR6形成異質二聚體發揮功能,參與識別多種抗原分子,如革蘭氏陽性菌的肽聚糖和脂磷壁酸、分枝桿菌的脂阿拉伯甘露糖、錐形蟲的糖基磷脂酰肌醇以及酵母或其它真菌的酵母多糖[18]。有研究表明受體和配體的相互作用能有效促進抗原遞呈細胞識別抗原的能力,進而促進細胞的吞噬及下游信號通路過程,豬TLR2就發揮這種受體功能,可高效促進豬抗原遞呈細胞識別并吞噬詹氏乳酸桿菌[19]。抗體封閉豬肺泡巨噬細胞(Porcine alveolar macrophages,PAMs)TLR2 和 TLR6可顯著降低豬肺炎支原體(Mycoplasma hyopneumoniae)誘導的TNF-α的表達水平,說明豬 TLR2和TLR6在宿主識別和抵御M.hyopneumoniae感染中起重要作用。真菌產生的脫氧雪腐鐮刀菌烯醇(Deoxynivalenol,DON)可破壞腸上皮細胞的屏障保護功能,可能還與腸道炎癥有關,而豬腸上皮細胞的TLR2相關信號通路可抑制DON對腸道粘膜屏障的破壞[20]。鼠李糖乳桿菌(Lactobacillus rhamnosus)CRL1505可以通過TLR2信號通路提高Th1型免疫反應,上調豬腸上皮細胞IL-6和TNF-α的表達[21]。
3.2 TLR4 TLR4在豬各種組織中廣泛存在,主要識別革蘭氏陽性菌的LPS或脂質A,可誘導產生多種促炎細胞因子的分泌,在抵抗革蘭氏陽性菌的感染上發揮重要作用,然而LSP過度上調促炎因子的表達可能會導致嚴重的炎癥反應。用LPS刺激豬小腸上皮細胞可顯著上調TLR4和炎性因子TNF-α、IL-1β的表達,說明在豬體內TLR4及其相關信號通路參與了LPS引起的炎癥反應[22]。嗜酸乳酸桿菌(Lactobacillus acidophilus)是一種非常重要的益生菌,可調節腸道菌群平衡,有效抑制腸道不良微生物的繁殖,研究發現L.acidophilus可通過TLR4/NF-κB信號通路降低豬外周血淋巴細胞促炎細胞因子的表達水平緩解LPS引起的炎癥反應,進而增加豬日進食量和日增重[23]。另外,豬TLR4可能還在抗病毒感染中起作用,如圓環病毒2型(Porcine circovirus type 2,PCV2)可上調TLR4的表達水平,進而激活下游信號通路,上調相關抗病毒細胞因子的表達[24]。
3.3 TLR5 TLR5特異地識別細菌的鞭毛蛋白,被激活后可誘導下游復雜的級聯反應,通過NF-κB信號通路,上調促炎細胞因子及趨化因子的表達。通過單核苷酸多態性分析發現民豬(Min pig)TLR5含有4個非同義單核苷酸多態性(Single nucleotide polymorphisms,SNPs)位點,分別為 c.176C>T(p.R59M)、c.902C>T(p.S301F)、c.959T>A(p.F320Y)和 c.1796C>T(p.T599M,reference sequence:GenBank No.AB208697),其中 SNP c.1796C>T(p.T599M)位于TLR5胞外區的一個N端糖基化位點編碼區域,豬TLR5 SNPs可能與細胞識別細菌鞭毛蛋白的能力密切相關[25]。
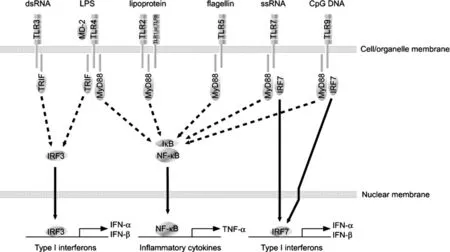
圖2 TLRs誘導的不同信號通路示意圖
3.4 TLR3、TLR7、TLR8 和 TLR9TLR3、TLR7、TLR8和TLR9分布在細胞器膜表面,主要通過上調抗病毒細胞因子I型干擾素(IFN-α/β)的表達發揮抗病毒作用。TLR3可識別病毒雙鏈RNA,在PAMs中TLR3在識別病毒RNA上起著重要作用;TLR7和TLR8識別單鏈RNA;TLR9識別非甲基化CpG DNA,這種核酸基序一般存在于細菌的基因組DNA中,也存在于病毒基因組中,如單純皰疹病毒2型[26]。病毒核酸被相應的TLRs識別后,下游的轉錄因子(包括IRF1、IRF3和IRF7)被激活,從而上調IFN-α/β的表達水平。如 H3N2豬流感病毒(Swine influenza virus,SIV)可 顯 著 提 高 PAMs 中TLR3、TLR7 的表達,進而上調 IFN-α 的表達[27];高致病性豬繁殖與呼吸綜合征病毒(Highly pathogenic porcine reproductive and respiratory syndrome virus,HP-PRRSV)可顯著上調TLR3、TLR7和TLR8的表達,進一步誘導 IL-1β、IL-6、TNF-α 和 IFN-γ 的分泌[28]。氧化劑和抗氧化劑可有效干預PRRSV對TLR3/NF-κB信號通路的激活,提示該信號通路的激活可能是通過細胞的氧化應激機制調節的[29]。然而目前很多研究表明PRRSV感染可下調宿主細胞IFN-α/β的表達,其分子機制也得到初步研究:PRRSV可阻止雙鏈RNA或仙臺病毒誘導的IRF3磷酸化及核轉位過程[30],與PCV2聯合感染可抑制TLR3、 TLR7 和 TLR9 的表達[31],進而抑制 IFN-α/β的表達。在腎臟的冷藏過程中,caspase-3小干擾RNA(Small interfering RNA,siRNA)灌注腎動脈可有效保存腎臟的活性,然而將此方法用于處理豬自體移植腎臟時,腎臟出現嚴重的炎癥反應和細胞凋亡,不能起到很好的保存效果,通過western blotting和quantitative PCR檢測發現,caspase-3 siRNA可上調豬自體移植腎臟TLR3、TLR7及其接頭分子TRIF和MyD88的表達水平,進而上調促炎細胞因子 IL-1、IL-6、TNF-α 和 IFN-α/β 的表達[32]。
4 總結與展望
TLRs是宿主識別入侵病原微生物的重要模式識別受體,同時也與自身免疫反應的調節及自身免疫病密切相關。因此,需要更加深入地了解TLRs在細胞內發揮的作用,以TLRs為靶標探索解決機體免疫失衡的方法。目前在藥物或病理研究方面,活體動物模型主要以小鼠為主[33],但是鼠和人的TLRs在功能上還存在較大的差異,比如用脂多糖(Lipopolysaccharides,LPS)處理小鼠和人的星狀膠質細胞,其TLR4發揮著不同的功能[34],這說明小鼠動物模型在人TLRs研究方面不能提供充分的數據。然而有證據顯示豬的TLRs在功能上與人更接近[35],越來越多的研究發現在人的某些傳染病和自然免疫病方面,豬可以作為較好的動物模型。因此,對豬TLRs深入系統地研究可為養豬業的健康發展和人類疾病研究提供重要參考。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
人大建設(2019年12期)2019-05-21 02:55:32
電子制作(2018年11期)2018-08-04 03:25:42
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
