大鼠脊髓背角神經元TRESK變化對lncRNA表達譜的影響*
2020-01-03 01:00:30王文康許立新阮祥才鄭彬林濤應彥璐李恒昌
中國醫(yī)學創(chuàng)新 2020年33期
王文康 許立新 阮祥才 鄭彬 林濤 應彥璐 李恒昌
雙孔鉀離子通道TRESK 在脊髓背根神經節(jié)廣泛表達,除了維持神經元靜息膜電位及神經遞質的釋放,還介導了脊髓水平神經病理性疼痛的發(fā)生,但具體機制不明確,TRESK 可被谷氨酸激活,但對脊髓神經元的作用不詳[1-2]。長鏈非編碼RNA(long non-coding RNA,lncRNA)是長度大于200 個核苷酸的非編碼RNA,在表觀遺傳、細胞周期和細胞分化等調控中發(fā)揮重要作用[3-4]。本研究觀察谷氨酸對大鼠脊髓神經元TRESK 的效應,通過基因芯片技術,檢測脊髓神經元TRESK 變化對lncRNA 表達譜的影響,探討TRESK 是否通過lncRNA 作用于下游信號通道發(fā)揮生物功能,為闡明脊髓神經元TRESK 在神經病理性疼痛中的作用機制奠定基礎。
1 材料與方法
1.1 動物和材料 動物為原代SD 大鼠,體質量180~250 g。主要材料:大鼠脊髓背角神經元(ScienCell,M1580-57)、神經元培養(yǎng)基(ScienCell,1521)、倒置光學顯微鏡(奧林巴斯,CKX41)、細胞恒溫培養(yǎng)箱(賽默飛,HERAcell150i)、lncRNA定量試劑盒(日本Takara 公司)。
1.2 方法
1.2.1 大鼠脊髓神經元培養(yǎng) 在無菌解剖液的平皿內分離脊髓,0.25%胰蛋白酶經37 ℃消化并吹打,吸取細胞懸浮液移置培養(yǎng)皿中。過濾及沉淀后加入3 mL 的培養(yǎng)基,稀釋計數(shù)后以5×105個/mL將細胞接種于經多聚賴氨酸處理的1×106個/孔,500 μL/ 孔,置于37 ℃、5%CO2培養(yǎng)箱中,1 d后更換飼養(yǎng)培養(yǎng)液。4 d 后于培養(yǎng)液內加入阿糖胞苷4 μg/mL,用于抑制非神經細胞增殖,培養(yǎng)7 d后進行實驗。
1.2.2 實驗分組和處理 培養(yǎng)原代大鼠脊髓神經元細胞,接種于培養(yǎng)皿中,采取隨機分組法,將其分為4 組:空白組(不進行任何處理)、谷氨酸(Glu)組(谷氨酸100 μmol/L 處理24 h)、Glu+NC siRNA組(siRNA 空白質粒轉染后同Glu 組方法處理)與Glu+TRESK siRNA 組(TRESK siRNA 轉染后同Glu組方法處理)。各組按照上述方法處理結束后,免疫印跡法和RT-PCR 分別檢測各組神經元的TRESK蛋白和mRNA 表達。
1.2.3 免疫印跡法 裂解細胞,離心后取上清液,用BCA 法測定蛋白濃度。蛋白上樣后電泳完畢,將蛋白轉移到PVDF 膜上(4 ℃、400 mA 恒流條件下120 min),室溫封閉PVDF 膜1 h(含5%脫脂牛奶的TBST 溶液),加抗4 ℃過夜;TBST 洗膜,加入辣根過氧化物酶偶聯(lián)二抗(1∶5 000),室溫下孵育2 h,TBST 洗膜3 次。采用ECL 試劑進行顯色反應后暗室內曝光顯影。
1.2.4 RT-PCR 檢測 谷氨酸處理24 h 后檢測脊髓神經元TRESK mRNA 表達。脊髓神經元離心勻漿,棄細胞上清液,用PBS 液潤洗細胞,采用Trizol 試劑盒提取總RNA,RNA 逆轉錄獲得cDNA。0.5 μg/μL寡脫氧核苷酸(Oligo dT)1 μL 和總RNA 2.0 μg加入PCR 小管中,補充焦碳酸二乙酯(DEPC)水至9 μL;混勻后離心,70 ℃溫浴10 min 后,立即置于0 ℃冰水混合物中冰浴,使Oligo dT 和模板退火,反應體系為:5×RT 緩沖液4.0 μL,10 mmol三磷酸脫氧核苷。
1.2.5 基因芯片檢測 檢測谷氨酸+TRESK siRNA與谷氨酸+NC siRNA 兩組lncRNA 表達譜變化,從總RNA 中移除rRNA 后,得到mRNA;使用隨機引物方法將每個樣品放大并轉錄成帶熒光的cRNA;使用純化標記的cRNAs,并用NanoDrop ND-1000 檢測濃度和活性。洗滌、固定并掃描雜交芯片。lncRNA譜包含探針原始信號的txt 文件導入到GeneSpring GX v12.1 軟件,標準化后,去除低質量探針,得到lncRNA 表達信息。利用Fold change 進行差異lncRNA 篩選,默認Fold change≥2.0,并根據(jù)lncRNA與蛋白編碼基因在基因組上的相對位置關系。
1.2.6 基因本體(GO)分析及KEGG 通路富集分析 GO 分析是采用Gene Ontology 數(shù)據(jù)庫對差異表達lncRNA 進行功能注釋,統(tǒng)計每個基因功能中存在的基因數(shù)量。KEGG 通路富集分析是基于京都基因與基因組百科全書(KEGG)數(shù)據(jù)庫來了解差異表達的lncRNA 富集的生物信號通路。分析后計算每個基因功能中差異基因的富集值,富集值越大,表示該基因功能或信號通路受到實驗影響越大。
1.3 統(tǒng)計學處理 采用SPSS 20.0 軟件對所得數(shù)據(jù)進行統(tǒng)計分析,計量資料用()表示,兩兩比較采用t 檢驗,多組比較采用方差分析。以P<0.05為差異有統(tǒng)計學意義。
2 結果
2.1 谷氨酸對TRESK siRNA 轉染大鼠脊髓神經元TRESK 表達情況 與空白組比較,Glu 組和Glu+NC siRNA 組TRESK 蛋白及mRNA 表達均明顯上調(P<0.05);與Glu+NC siRNA 組相比,Glu+TRESK siRNA 組脊髓神經元TRESK 蛋白及mRNA 表達均明顯下調(P<0.05)。見表1。
表1 谷氨酸對TRESK siRNA轉染大鼠脊髓神經元TRESK表達情況()
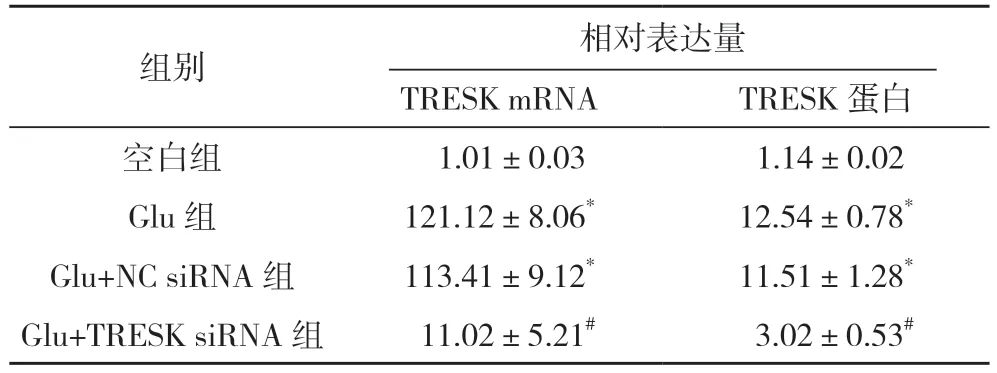
表1 谷氨酸對TRESK siRNA轉染大鼠脊髓神經元TRESK表達情況()
注:* 與空白組相比,P<0.05;# 與Glu+NC siRNA 組相比,P<0.05。
2.2 谷氨酸誘導神經元TRESK 中l(wèi)ncRNA 差異表達結果 本次實驗中共檢測lncRNA 基因經聚類分析得出,與Glu+NC siRNA 組比較,Glu+TRESK siRNA 組出現(xiàn)lncRNA 差異表達基因322 條,其中212 條基因表達上調,110 條基因表達下調。所有表達差異的lncRNA 基因中,uc007pgs.1 基因表達上調倍數(shù)最多,約為41.61 倍;而表達下調倍數(shù)最多基因為ENSMUST00000161848,約為6.97 倍。其中表達上調及下調差異倍數(shù)最大的前5 條lncRNA差異表達。見表2。
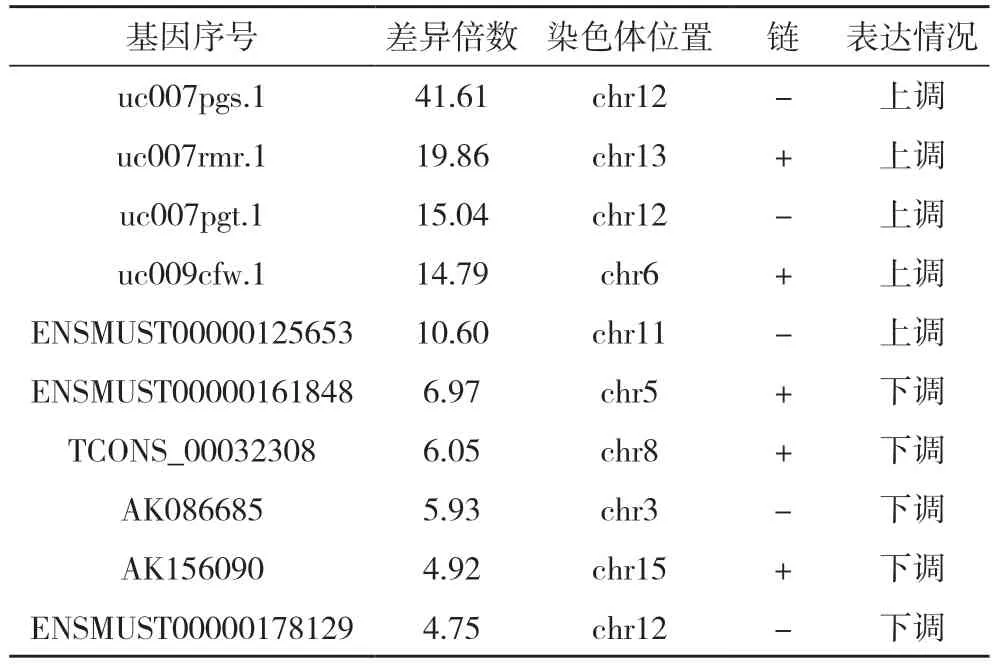
表2 Glu+NC siRNA組與Glu+TRESK siRNA組部分lncRNA基因差異表達
2.3 lncRNA 差異表達基因生物信息分析 GO 分析結果顯示,lncRNA 差異表達的靶基因功能更多與炎癥反應、凋亡過程及生物過程等有關,見圖1。KEGG 富集通路分析結果顯示,差異顯著的lncRNA 基因調控的靶基因主要參與mTOR 信號通路、蛋白激酶信號通路和GRCP 信號通路等下游通路,見圖2。
3 討論
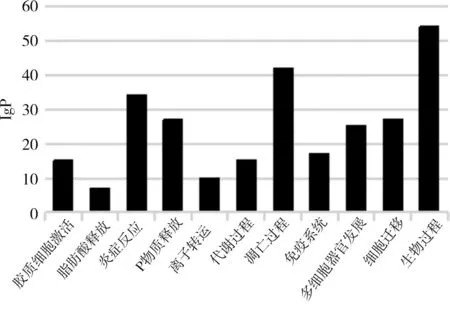
圖1 GO富集分析
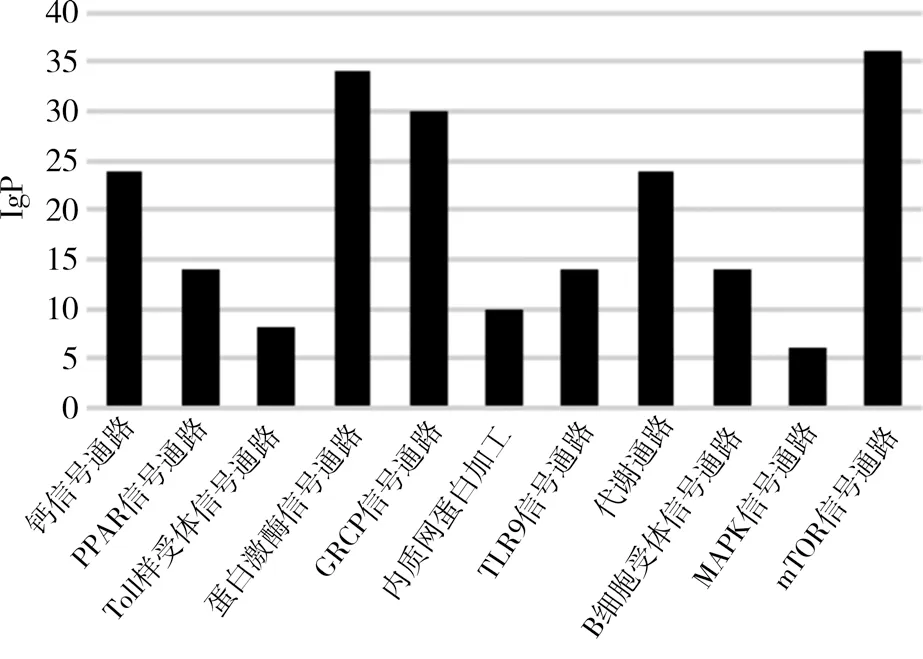
圖2 KEGG富集通路分析
神經病理性疼痛是臨床中常見慢性疼痛,其形成及病程發(fā)展機制或經多種神經遞質參與,發(fā)病機制較為復雜。文獻[5-6]報道,谷氨酸可誘導脊髓神經元TRESK 凋亡,且與慢性疼痛的發(fā)生機制密切相關。而神經病理性疼痛的病理過程涉及大量細胞分子的變化,許多研究證實了lncRNA 可調節(jié)編碼基因的表達,從而影響神經發(fā)育,且富集于大鼠中樞神經,密切參與許多神經系統(tǒng)性疾病[7-8]。因此,通過研究脊髓神經元TRESK 的變化及其lncRNA 差異表達情況,分析差異基因參與機制和信號通路分析,進一步闡明神經病理性疼痛的病理過程。
在本次研究中,大鼠脊髓神經元TRESK 經谷氨酸之后其mRNA 和蛋白表達量上調,TRESK siRNA 轉染后,Glu+TRESK siRNA 組相比較于Glu+NC siRNA 組,脊髓神經元的TRESK 蛋白及mRNA 表達明顯下調,說明谷氨酸對TRESK 具神經元突觸及功能調控作用,與文獻[9-10]研究報道結果相符。通過谷氨酸處理神經元TRESK 蛋白24 h之后的lncRNA 進行基因芯片高通量測序,以篩選差異倍數(shù)最大且差異顯著基因322 條,其中212 條基因表達上調,110 條基因表達下調,這些差異表達lncRNA 基因或與神經元TRESK 參與神經病理性疼痛的生物功能進程有關[11]。為進一步發(fā)掘lncRNA 顯著差異表達靶基因的潛在作用和效應功能,進行了GO 富集分析和KEGG 富集通路分析,其中當富集值越大是,其代表差異基因參與生物過程和涉及信號通路更密切[12-13]。結果在GO 富集分析中,差異顯著的靶基因更多富集于炎癥反應、凋亡過程及生物過程等,提示這些差異顯著的靶基因可促進分子結合與分化和信息傳導,當減少神經元凋亡可印制傷害信息傳導,致脊背根神經節(jié)向脊髓傳導痛覺傷害性神經元的興奮增高,進而產生痛覺反應,表明lncRNA 或參與神經功能行使[14-16]。KEGG 富集通路分析可以發(fā)現(xiàn),PI3K-AKT 信號通路有許多功能,差異顯著的lncRNA 靶基因主要涉及mTOR 信號通路、蛋白激酶信號通路和GRCP 信號通路等下游通路,表明神經元TRESK 中差異表達的lncRNA 可能通過以上信號通路直接或間接參與神經病理性疼痛病程發(fā)展[17-20]。然而對于差異lncRNA 在上述信號通路的調控機制仍有待進一步研究,且本研究僅探討lncRNA 表達譜的差異,對于共表達的mRNA 在神經元TRESK 的生物學功能未深入展開分析。
綜上,谷氨酸誘導神經元TRESK 中相關lncRNA 表達譜發(fā)生差異,從而引起細胞炎癥反應和神經元凋亡,其中mTOR 信號通路、蛋白激酶信號通路和GRCP 信號通路可能是主要的靶點信號通路,以期為闡明臨床中神經病理性疼痛的發(fā)病機制提供理論性參考依據(jù)。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
鴨綠江(2021年35期)2021-04-19 12:24:18
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
電子制作(2018年18期)2018-11-14 01:48:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
電子制作(2018年11期)2018-08-04 03:25:42
山東工業(yè)技術(2016年15期)2016-12-01 05:31:22
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
