ARID1A、E-cadherin和EphA2在胃癌及癌前病變中的表達及臨床意義
2020-01-08 09:04:24
國際消化病雜志 2019年6期
人ARID1A基因,即AT豐富結合域1A,屬于酵母交換型轉換/蔗糖不發酵(SWI/SNF)復合物的非催化亞基,其功能障礙可引起染色質重塑異常,導致基因表達調節紊亂,進而引發多種疾病。人源性E-鈣黏蛋白(E-cadherin)由CDH1基因編碼,主要分布于上皮細胞表面,可促進細胞間的連接,與細胞的黏附功能密切相關,而該功能對于多種組織中不同細胞的細胞間隙及細胞相互作用的穩態至關重要[1]。EphA2基因,即促紅細胞生成素產生肝細胞A2受體,是首個被篩選出來的 Eph 受體酪氨酸亞家族成員,其在細胞膜上有磷酸化和非磷酸化兩種狀態,在正常上皮細胞中,EphA2受體與其配體結合,構象發生改變,導致自身磷酸化而被降解;而腫瘤細胞的EphA2受體主要以非磷酸化狀態存在,并且表達增高。本文探究了三者在胃黏膜癌變過程中的作用、三者之間的相互關系及臨床意義,為胃癌的早期診斷及治療提供參考。
1 對象和方法
1.1 研究對象
收集2017年6月至2018年12月在內蒙古醫科大學第三附屬醫院消化內科內窺鏡中心行胃鏡檢查,且同時行活組織檢查的150例患者的組織石蠟標本,分為慢性非萎縮性胃炎組、慢性萎縮性胃炎伴腸化組、低級別上皮內瘤變組、高級別上皮內瘤變組和胃癌組,每組各30例。所有標本均由兩位內鏡專家及兩位病理學專家根據內鏡形態學和病理組織學特征確認。入組標準:(1)簽署入組知情同意書;(2)胃癌患者檢查前未接受化學治療或放射治療;(3)近4周內未服用質子泵抑制劑、糖皮質激素類藥物、非甾體類抗炎藥(NSAID);(4)近4周內無飲酒史。排除標準:(1)存在胃鏡檢查禁忌證;(2)身體其他部位存在腫瘤;(3)存在嚴重的肝腎功能不全、肝硬化、糖尿病及凝血功能異常。
1.2 實驗分組情況
慢性非萎縮性胃炎組30例,其中男性17例,女性13例,平均年齡(38.47±11.90)歲;慢性萎縮性胃炎伴腸化組30例,其中男性15例,女性15例,平均年齡(38.80±11.84)歲;低級別上皮內瘤變組30例,其中男性20例,女性10例,平均年齡(41.83±9.83)歲;高級別上皮內瘤變組30例,其中男性20例,女性10例,平均年齡(41.50±8.38)歲;胃癌組30例,其中男性22例,女性8例,平均年齡(43.10±9.72)歲。各組患者的性別及年齡相比較,差異無統計學意義(P>0.05),具有可比性。
1.3 主要材料和試劑
ARID1A兔抗人多克隆抗體、E-cadherin兔抗人多克隆抗體、EphA2兔抗人多克隆抗體均購自武漢博士德生物工程有限公司。即用型免疫組織化學超敏SP檢測試劑盒、DAB顯色試劑盒、PBS磷酸鹽緩沖液(粉劑)、檸檬酸鹽抗原修復液均購自福州邁新生物技術公司。
1.4 實驗方法
將標本組織進行固定、脫水、浸蠟、包埋和切片處理后,按5 μm厚度切取3張切片,分別進行ARID1A、E-cadherin、EphA2的免疫組織化學染色。實驗步驟:將選擇好的組織石蠟切片進行常規脫蠟、脫水→高壓鍋中進行抗原熱修復→加入內源性過氧化物酶→加入非免疫性動物血清封閉液→分別滴加1∶100的一抗稀釋液蓋住組織,4 ℃過夜→加入生物素標記的二抗→加入鏈霉菌抗生物素蛋白-過氧化物酶→滴加DAB工作液進行顯色→蘇木素復染,梯度酒精中脫水、透明、封片,玻片晾干后鏡下觀察,隨后進行圖片采集及結果匯總。
1.5 染色結果
ARID1A、E-cadherin和EphA2的免疫組織化學陽性產物為棕色或棕褐色顆粒,其中ARID1A定位于細胞核中,E-cadherin定位于細胞膜和細胞質中,EphA2定位于細胞質中。Bresalier半定量公式用于判斷各組的染色情況,隨機選取高倍鏡下的10個視野區,根據細胞染色強度對每個高倍視野區進行分級,共分為4級:陰性(-)、弱陽性(+)、強陽性(++)和強陽性(+++);細胞未著色計0分,淡黃色計1分,棕黃色計2分,棕褐色計3分。通過計數已著色的視野數,按照下列公式計算切片的平均染色強度:IS(intensity store)=∑{(0×F0)+(1×F1)+(2×F2)+(3×F3)},F= %×10視野。
1.6 統計學處理
采用SPSS 21.0軟件進行數據分析及處理。采用Krusral-wallisH非參數檢驗,P<0.05為差異有統計學意義。對慢性非萎縮性胃炎組、慢性萎縮性胃炎伴腸化組、低級別上皮內瘤變組、高級別上皮內瘤變組、胃癌組間進行Bonferroni法多重比較(組間采用Mann-WhitneyU檢驗),以校驗P*<0.005(P<0.05)為差異有統計學意義。通過Kendall′stau-b等級相關分析法分析ARID1A、E-cadherin和EphA2之間表達的相關性,以P<0.05為差異有統計學意義。
2 結果
2.1 ARID1A在各組中表達水平的比較
采用Krusral-wallisH方法檢驗ARID1A在各組中的表達秩均值,結果分別為105.65、95.87、79.33、53.03和43.62,表達呈逐漸遞減趨勢,差異具有統計學意義(χ2=49.188,P<0.01)。對各組間進行Bonferroni法多重比較,結果顯示ARID1A在慢性非萎縮性胃炎組與慢性萎縮性胃炎伴腸化組中的表達水平差異無統計學意義(P>0.05),在慢性非萎縮性胃炎組、慢性萎縮性胃炎伴腸化組與低級別上皮內瘤變組中的表達水平差異具有統計學意義(P<0.05),在低級別上皮內瘤變組與高級別上皮內瘤變組、胃癌組中的表達水平差異具有統計學意義(P<0.05)。見圖1。
2.2 E-cadherin在各組中表達水平的比較
采用Krusral-wallisH方法檢驗 E-cadherin在各組中的表達秩均值,結果分別為104.38、96.43、76.68、55.93和44.07,表達呈逐漸遞減趨勢,差異具有統計學意義(χ2=46.670,P<0.01)。對各組間進行Bonferroni法多重比較,結果顯示E-cadherin在慢性非萎縮性胃炎組與慢性萎縮性胃炎伴腸化組中的表達水平差異無統計學意義(P>0.05),在慢性非萎縮性胃炎組、慢性萎縮性胃炎伴腸化組與低級別上皮內瘤變組中的表達水平差異具有統計學意義(P<0.05),在低級別上皮內瘤變組與高級別上皮內瘤變組、胃癌組中的表達水平差異具有統計學意義(P<0.05)。見圖2。
2.3 EphA2在各組中表達水平的比較
采用Krusral-wallisH方法檢驗EphA2在各組中的表達秩均值,結果分別為34.35、62.32、78.05、99.37和103.42,表達呈逐漸遞增趨勢,差異具有統計學意義(χ2=57.488,P<0.01)。對各組間進行Bonferroni法多重比較,結果顯示EphA2在慢性萎縮性胃炎伴腸化組與低級別上皮內瘤變組中的表達水平差異無統計學意義(P>0.05);在高級別上皮內瘤變組與胃癌組中的表達水平差異無統計學意義(P>0.05);在慢性萎縮性胃炎伴腸化組、低級別上皮內瘤變組中與高級別上皮內瘤變組、胃癌組中的表達水平差異具有統計學意義(P<0.05),與慢性非萎縮性胃炎組中的表達水平差異具有統計學意義(P<0.05)。見圖3。
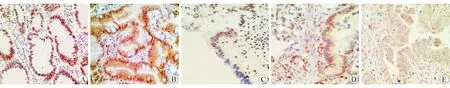
圖1ARID1A在不同胃黏膜組織中的表達水平 免疫組織化學染色 ×400A慢性非萎縮性胃炎組B慢性萎縮性胃炎伴腸化組C低級別上皮內瘤變組D高級別上皮內瘤變組E胃癌組
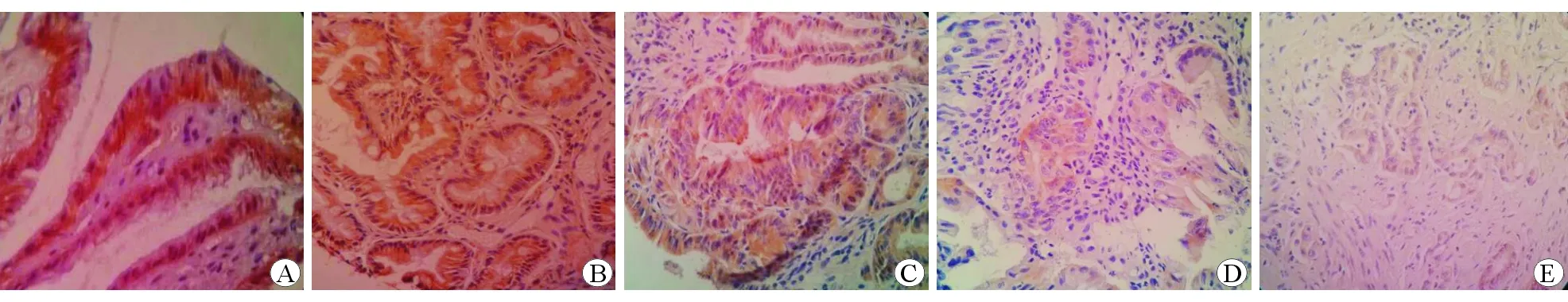
圖2E-cadherin在不同胃黏膜組織中的表達水平 免疫組織化學染色 ×400A慢性非萎縮性胃炎組B慢性萎縮性胃炎伴腸化組C低級別上皮內瘤變組D高級別上皮內瘤變組E胃癌組
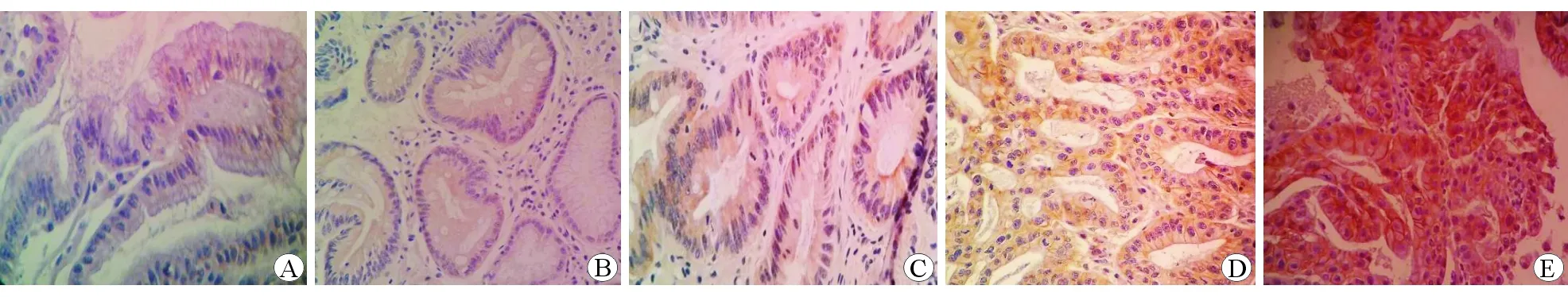
圖3EphA2在不同胃黏膜組織中的表達水平 免疫組織化學染色 ×400A慢性非萎縮性胃炎組B慢性萎縮性胃炎伴腸化組C低級別上皮內瘤變組D高級別上皮內瘤變組E胃癌組
2.4 ARID1A、E-cadherin和EphA2在各組中相關性的比較
采用Kendall′stau-b方法檢驗ARID1A、E-cadherin和EphA2在各組間表達的相關性。結果顯示,在慢性非萎縮性胃炎組中,ARID1A與E-cadherin表達呈顯著正相關,E-cadherin與EphA2表達呈顯著負相關,ARID1A與EphA2表達呈顯著負相關(P均<0.05)。在慢性萎縮性胃炎伴腸化組中,ARID1A與E-cadherin表達呈顯著正相關,E-cadherin與EphA2表達呈顯著負相關,ARID1A與EphA2表達呈顯著負相關(P均<0.05)。在低級別上皮內瘤變組中,ARID1A與E-cadherin表達呈顯著正相關,E-cadherin與EphA2表達呈顯著負相關,ARID1A與EphA2表達呈顯著負相關(P均<0.05)。在高級別上皮內瘤變組中,ARID1A與E-cadherin表達呈顯著正相關,E-cadherin與EphA2表達呈顯著負相關,ARID1A與EphA2表達呈顯著負相關(P均<0.05)。在胃癌組中,ARID1A與E-cadherin表達呈顯著正相關,E-cadherin與EphA2表達呈顯著負相關,ARID1A與EphA2表達呈顯著負相關(P均<0.05)。具體見表1、2、3。
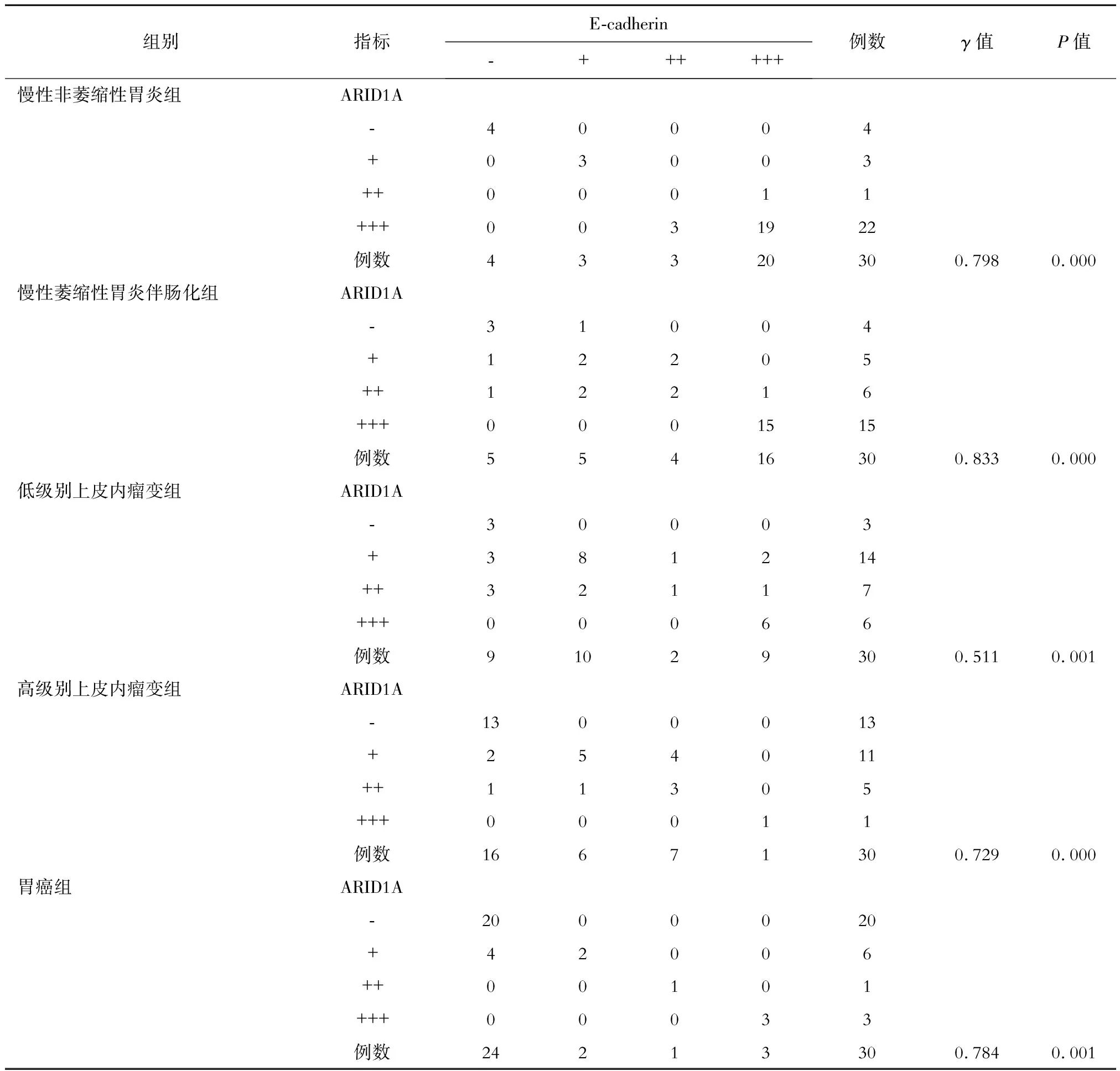
表1 各組中ARID1A與E-cadherin表達強度的相關性分析

表2 各組中EphA2與E-cadherin表達強度的相關性分析
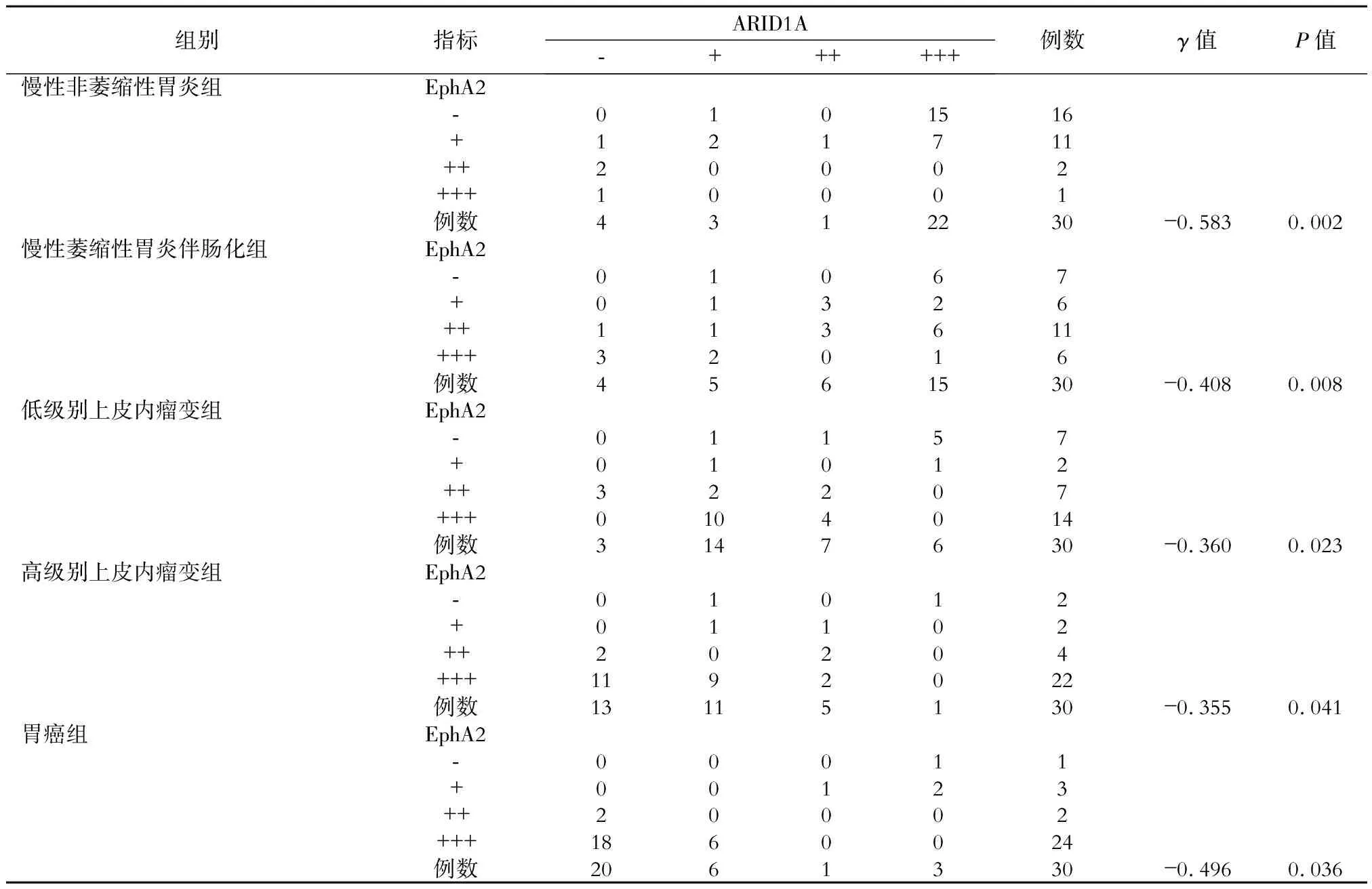
表3 各組中EphA2與ARID1A表達強度的相關性分析
3 討論
SWI/SNF復合物是重要的染色質重塑復合物之一,主要由核心亞單位、催化亞單位和調節單位組成,借助三磷酸腺苷(ATP)水解釋放的能量更改核小體與基因啟動子、增強子等區域的DNA結合位點,調控基因的轉錄表達。作為SWI/SNF復合物的調節亞單位之一,ARID1A可以特異性地調節該過程的基因表達[2]。近年來由于外顯子測序的廣泛應用,發現ARID1A在多種腫瘤中存在高頻失活突變,這是許多惡性腫瘤發生的重要原因[3]。另外,ARID1A突變與EB病毒(EBV)感染、微衛星不穩定性顯著相關[4]。Zhang等[5]的研究發現,ARID1A表達下調會促進胃癌細胞增殖、細胞生長及營養消耗,同時還會直接作用于PIK3CA和PDK1靶基因,激活PI3K/Akt信號通路,參與胃癌的發生、發展。本研究結果表明,ARID1A在各組中的表達呈逐漸遞減趨勢,推測ARID1A在胃癌癌前病變進展至胃癌的過程中存在表達缺失,其可能作為一種抑癌基因在胃癌的發展過程中發揮重要作用。此外,本研究發現ARID1A在高級別上皮內瘤變中的表達水平明顯降低,雖通過多重比較發現高級別上皮內瘤變組與胃癌組間的表達差異無統計學意義(校驗前P*>0.005,校驗后P>0.05),但單獨比較ARID1A在兩組中的表達水平差異仍具有統計學意義(校驗前P*<0.05),提示該基因的表達失活參與了胃癌的形成過程。因此,ARID1A可能在胃癌的靶向性預防及治療中發揮重要作用。ARID1A的抑癌機制非常復雜,如果能進一步具體闡明其抑癌機制,將對腫瘤的形成、發展及診治的研究提供一個新的思路。
E-cadherin是一種鈣依賴性跨膜糖蛋白,主要介導同源細胞與細胞間的連接和黏附,是上皮細胞中維護細胞形態結構完整性和極性的重要黏附分子,具有抑制腫瘤細胞浸潤和轉移的功能。在正常組織中,E-cadherin表達下調可降低同種細胞間的黏附功能,細胞形態發生變化,導致功能障礙,不能正常發育;在腫瘤細胞中,E-cadherin表達下調可降低腫瘤細胞間的黏附作用,細胞骨架形態發生改變,有利于腫瘤細胞從原發灶脫落向遠處侵襲、轉移[6]。本研究結果表明,E-cadherin在各組中表達呈逐漸遞減趨勢,提示E-cadherin表達降低可升高胃癌的發生率。此外,本研究發現E-cadherin在高級別上皮內瘤變中的表達水平明顯降低,經多重比較發現高級別上皮內瘤變組與胃癌組間的表達差異無統計學意義(校驗前P*>0.005,校驗后P>0.05),推測可能有以下幾種原因:一方面,胃黏膜高級別上皮內瘤變實際上大部分已同時存在癌變[7],其浸潤和轉移的危險度明顯升高[8];另一方面,由于病理科醫生對早期胃癌不夠重視,在診斷尺度把握上也存在差異,對于無法看到的浸潤性癌變的活組織檢查樣本診斷保守,可能將惡性腫瘤誤判為高級別上皮內瘤變。因此,E-cadherin可能是胃黏膜癌變的早期分子事件,從低級別上皮內瘤變這一階段開始進行積極有效地監測或干預,可能成為防治胃癌的有效途徑之一。
EphA2屬于受體酪氨酸激酶(RTK)亞家族之一,可通過胞外配體結合區與其配體EphrinA1結合形成受體-配體復合物,使其構象發生改變,導致自身磷酸化被降解,通過一系列信號轉導參與細胞的生長、遷移和分化,在胚胎發育、血管生成、腫瘤形成等方面起重要作用。研究發現,EphA2通常在正常上皮細胞中低表達,而在膠質瘤、胃癌、肝細胞癌、結直腸癌等實體瘤中過表達,并且與腫瘤的轉移和預后密切相關[9-12]。翟道寬等[13]的研究表明,在肝細胞癌中EphA2可通過調節E-cadherin的表達來發揮腫瘤細胞侵襲、轉移作用,其機制可能是活化了Wnt/β-catenin通路,從而促進肝癌細胞發生上皮-間質轉化(EMT),因此EphA2可能是腫瘤EMT過程中的調控分子。本研究結果表明,EphA2在各組中表達呈逐漸遞增趨勢,提示EphA2的異常活化可能是胃癌的早期事件,對胃癌的形成和發展具有重要作用。因此,如果能盡早地在分子水平上抑制EphA2表達,有望阻止胃癌的形成或發展。
本研究結果顯示,ARID1A、E-cadherin和EphA2在各組中的表達具有相關性。其中,ARID1A與E-cadherin在各組中的表達呈正相關,提示兩者可能協同促進胃癌的發生、發展,這也證實了Yan等[14]的研究結果,他們通過在胃癌細胞中構建E-cadherin啟動子區域的報告基因,結果顯示敲低ARID1A后,含CDH1啟動子區域的報告基因數值顯著下降,E-cadherin的表達水平也隨之降低,細胞遷移、侵襲能力變強,提示ARID1A與CDH1的轉錄相關,由此得出ARID1A可能通過基因轉錄調控E-cadherin的表達,是腫瘤EMT過程中的調控分子。在上皮細胞中,E-cadherin與EphA2共同定位于上皮細胞的外側緣,特別是細胞接觸的部位[15],因此,兩者可以相互調節。一方面,E-cadherin表達缺失和功能改變會使細胞的黏附性降低,影響EphA2的磷酸化及其與膜結合配體的膜定位,從而升高EphA2的表達[16];另一方面,EphA2表達升高也會破壞黏附連接,加速細胞接觸的分離,導致細胞接觸處E-cadherin表達下調更快[17]。本研究結果顯示E-cadherin與EphA2在各組中的表達呈負相關,推測可能由于E-cadherin與EphA2黏附功能方面相互影響,形成負反饋環,因此,兩者在胃黏膜癌變過程中可能存在拮抗作用。此外,本研究還發現,ARID1A與EphA2在各組中的表達呈負相關,推測在胃癌發生過程中,由于ARID1A的失活突變而觸發了E-cadherin的低表達,而EphA2高表達的主要原因與E-cadherin表達降低有關,因而ARID1A在此過程中占據主導地位,EphA2可能受到ARID1A的間接調控作用。眾所周知,EMT被認為是腫瘤細胞侵襲和轉移的特征性事件,而E-cadherin表達下降或缺失是 EMT 的重要標志之一[18]。ARID1A和EphA2作為腫瘤EMT過程中的調控分子,異常表達后均可下調E-cadherin的表達,促進腫瘤細胞的侵襲、轉移。因此,ARID1A、E-cadherin和EphA2在胃黏膜癌變過程中可相互影響,且其中ARID1A起決定性作用。
綜上所述,ARID1A蛋白、E-cadherin蛋白的低表達,以及EphA2蛋白的高表達在胃癌的發生、發展過程中發揮著重要的作用,共同促進了胃癌的形成與發展,早期檢測三者在胃癌癌前病變中的表達水平,將有助于判斷病變的嚴重程度和進展趨勢,能夠更早地干預和治療早期胃癌、逆轉癌前病變,有望成為日后胃癌免疫治療的新亮點。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國組織化學與細胞化學雜志(2016年3期)2016-02-27 11:15:35
中國衛生標準管理(2015年3期)2016-01-14 03:41:46
醫學研究雜志(2015年6期)2015-07-01 17:40:49
醫學研究雜志(2015年9期)2015-07-01 17:28:27
中國當代醫藥(2015年20期)2015-03-01 02:04:29
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
