漢黃芩素對LPS-ATP誘導(dǎo)的巨噬細(xì)胞氧化應(yīng)激的抑制作用①
2020-02-20 12:10:10周思瑤游雷鳴張海麗趙夢繁蘇日娜
中國免疫學(xué)雜志 2020年3期
周思瑤 游雷鳴 張海麗 趙夢繁 蘇日娜 劉 慧 郝 鈺
(北京中醫(yī)藥大學(xué)生命科學(xué)學(xué)院免疫與微生物學(xué)系,北京 100029)
漢黃芩素是從中藥黃芩、半枝蓮等植物中分離得到的一種重要的黃酮類化合物。現(xiàn)代藥理研究發(fā)現(xiàn),漢黃芩素具有明顯抗炎作用[1]。且相關(guān)研究表明,漢黃芩素可通過抑制NF-κB和MAPK信號通路抑制LPS誘導(dǎo)的大鼠背根神經(jīng)節(jié)神經(jīng)元炎癥反應(yīng)[2]。漢黃芩素能通過抗炎及抗氧化作用來減輕糖尿病性心肌病[3]。且漢黃芩素還可通過抑制氧化應(yīng)激誘導(dǎo)的MAPK和NF-κB途徑來減輕大鼠體內(nèi)鎘誘導(dǎo)的腎毒性[4]。還有文獻(xiàn)表明,漢黃芩素可通過激活Nrf2/HO-1-SOD2-NQO-1-GCLC信號轉(zhuǎn)導(dǎo)軸來抑制IL-1β刺激引起的MAPK信號通路活化,從而抑制炎癥反應(yīng)[5]。我們從大量參考文獻(xiàn)中得出,漢黃芩素具有明顯的抗氧化應(yīng)激及抗炎作用。
我們課題組已有前期研究證明,漢黃芩素能抑制 LPS-ATP 聯(lián)合刺激巨噬細(xì)胞炎癥反應(yīng)的發(fā)生,與 LPS 和 ATP 聯(lián)合刺激組相比,漢黃芩素干預(yù)組能明顯降低胞內(nèi) IL-1β、IL-18、IL-6、TNF-α、NLRP3和 Caspase-1 的 mRNA 水平,減少NF-κB核轉(zhuǎn)位,并能下調(diào)胞外TNF-α和胞內(nèi)ROS水平[6]。ROS是氧化應(yīng)激的重要產(chǎn)物,大量的ROS又可以激活巨噬細(xì)胞的核轉(zhuǎn)錄因子 NF-κB使其發(fā)生核轉(zhuǎn)位,誘導(dǎo)炎癥反應(yīng)[7]。由此,我們認(rèn)為漢黃芩素可能是通過降低胞內(nèi)的ROS水平,減少NF-κB核轉(zhuǎn)位,抑制NF-κB介導(dǎo)的炎癥基因轉(zhuǎn)錄繼而減輕巨噬細(xì)胞的炎癥反應(yīng)。那么,是什么機(jī)制使胞內(nèi)ROS水平降低的?是漢黃芩素直接減少ROS產(chǎn)生和釋放還是通過激活抗氧化信號通路降解過多的ROS?Nrf2信號通路是細(xì)胞內(nèi)氧化應(yīng)激最重要的防御系統(tǒng)之一,當(dāng)細(xì)胞發(fā)生氧化應(yīng)激,Nrf2 信號通路首先被激活,進(jìn)而引發(fā)抗氧化物及相關(guān)酶類的大量表達(dá),以抵御氧化應(yīng)激引發(fā)的細(xì)胞損傷,減少ROS產(chǎn)生[8]。為此,本研究以脂多糖和三磷酸腺苷聯(lián)合刺激小鼠巨噬細(xì)胞為炎癥模型,從抗氧化的角度探究漢黃芩素抑制巨噬細(xì)胞炎癥反應(yīng)的作用機(jī)制,并探究漢黃芩素對Nrf2信號通路的影響。
1 材料與方法
1.1實(shí)驗(yàn)材料 小鼠巨噬細(xì)胞系RAW264.7(ATCC:TIB-71),由北京中醫(yī)藥大學(xué)免疫與微生物學(xué)系實(shí)驗(yàn)室保存。漢黃芩素購自中國藥品生物制品檢定所,反轉(zhuǎn)錄試劑盒、ATP購自TaKaRa公司,RNA提取試劑TRIZOL購自Invitrogen公司,SYBGreen熒光定量PCR試劑盒購自ToYoBo公司,LPS購自Sigma公司,SOD、MDA檢測試劑盒購自凱基生物公司,胎牛血清購自HyClone公司,Anti-Nrf2抗體、山羊抗兔IgG購自Abcam公司,4%組織細(xì)胞固定液購自索萊寶公司,山羊血清購自中杉金橋公司,NO試劑盒購自南京建成生物工程研究所。引物采用Primer5.0設(shè)計(jì),由生工生物工程(上海)股份有限公司合成(表1)。
1.2實(shí)驗(yàn)方法
1.2.1細(xì)胞培養(yǎng) RAW264.7細(xì)胞用含 10%胎牛血清的 DMEM 高糖培養(yǎng)基在37℃、5%CO2的無菌培養(yǎng)箱中培養(yǎng)。鏡下觀察細(xì)胞密度達(dá)80%左右予以傳代。
1.2.2檢測試劑盒測定巨噬細(xì)胞SOD、MDA、NO的分泌水平 將對數(shù)生長期的巨噬細(xì)胞RAW264.7接種到6孔板中(1×106個(gè)細(xì)胞/孔),設(shè)對照組(正常培養(yǎng)不予以刺激)、模型組(先加LPS刺激4 h后再加ATP刺激30 min,LPS和ATP終濃度均為 1 μg/ml)和漢黃芩素(56 μmol/L、28 μmol/L、14 μmol/L)干預(yù)組(加入LPS的同時(shí)加入漢黃芩素,4 h后加ATP刺激30 min),每組4個(gè)復(fù)孔。取培養(yǎng)皿中的上清,按照SOD、MDA、NO檢測試劑盒說明書,分別對各組細(xì)胞上清中的SOD、MDA、NO水平進(jìn)行檢測。
1.2.3免疫熒光法檢測巨噬細(xì)胞Nrf2核轉(zhuǎn)位 將巨噬細(xì)胞RAW264.7制成細(xì)胞濃度為1×104個(gè)/ml的細(xì)胞懸液,接種1 ml到20 mm激光共聚焦培養(yǎng)皿中。設(shè)對照組、模型組、漢黃芩素干預(yù)組,各組處理方式同方法1.2.2。待細(xì)胞生長至50%~60%,吸凈培養(yǎng)基,用PBS清洗細(xì)胞(3次,3 min/次)。每皿加2 ml 4%多聚甲醛溶液,室溫固定細(xì)胞15 min,用PBS清洗細(xì)胞(3次,3 min/次)。接著,用0.1%Triton X-100處理細(xì)胞15 min(室溫透化),PBS清洗細(xì)胞(3次,5 min/次),然后用山羊血清封閉細(xì)胞1 h(每皿1 ml),吸凈封閉液,加入兔抗Nrf2抗體工作液(山羊血清1∶500稀釋),37℃孵育2 h。PBS洗細(xì)胞(3次,5 min/次),加入羊抗兔IgG的二抗工作液(山羊血清1∶500稀釋),37℃孵育1 h。PBS洗細(xì)胞(3次,5 min/次)。加入DAPI對細(xì)胞核染色(藍(lán)色),室溫避光孵育5 min。PBS洗細(xì)胞(3次,5 min/次),在激光共聚焦顯微鏡下觀察Nrf2(紅色)的定位情況。
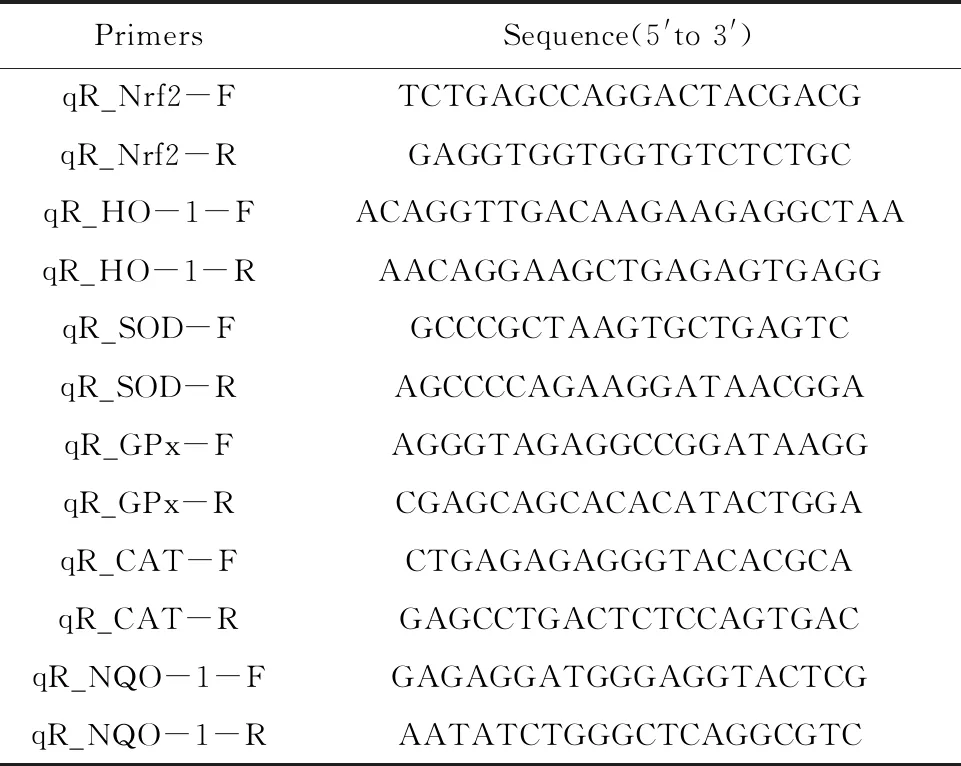
1.2.4RT-qPCR檢測巨噬細(xì)胞Nrf2和抗氧化酶mRNA水平 將巨噬細(xì)胞RAW264.7接種到6孔板中(2×106個(gè)細(xì)胞/孔),設(shè)對照組、模型組、漢黃芩素(56 μmol/L、28 μmol/L、14 μmol/L)干預(yù)組,每組3個(gè)復(fù)孔,各組處理方法與1.2.2相同。棄去各孔培養(yǎng)基后用PBS清洗各孔細(xì)胞,加入TRIZOL試劑裂解細(xì)胞(500 μl/孔),參照說明書的操作要求分別提取各孔細(xì)胞的總RNA。參照反轉(zhuǎn)錄試劑盒操作說明,各取1 μg RNA配制10 μl反轉(zhuǎn)錄體系合成相應(yīng)的cDNA,得到的cDNA用DEPC水做10倍稀釋后備用。取1 μl稀釋后的cDNA作為模板,以小鼠GAPDH基因作為內(nèi)參,加入相應(yīng)的引物(表1),利用實(shí)時(shí)熒光定量PCR(qPCR)法檢測各組樣品中相應(yīng)基因的mRNA水平。定量分析基因包括Nrf2、HO-1、SOD、GPx、CAT以及NQO-1,qPCR數(shù)據(jù)采用2-ΔΔCt法進(jìn)行處理和分析。
表1 設(shè)計(jì)并合成的引物
Tab.1 Primers designed and synthesized in this study
PrimersSequence(5'to 3')qR_Nrf2-FTCTGAGCCAGGACTACGACGqR_Nrf2-R GAGGTGGTGGTGTCTCTGCqR_HO-1-FACAGGTTGACAAGAAGAGGCTAAqR_HO-1-RAACAGGAAGCTGAGAGTGAGGqR_SOD-FGCCCGCTAAGTGCTGAGTCqR_SOD-RAGCCCCAGAAGGATAACGGAqR_GPx-FAGGGTAGAGGCCGGATAAGGqR_GPx-RCGAGCAGCACACATACTGGAqR_CAT-FCTGAGAGAGGGTACACGCAqR_CAT-RGAGCCTGACTCTCCAGTGACqR_NQO-1-FGAGAGGATGGGAGGTACTCGqR_NQO-1-RAATATCTGGGCTCAGGCGTC
2 結(jié)果
2.1漢黃芩素干預(yù)能明顯降低LPS和ATP聯(lián)合刺激的巨噬細(xì)胞MDA、NO分泌水平 模型組(LPS-ATP聯(lián)合刺激)與對照組相比,巨噬細(xì)胞分泌的NO、MDA顯著增多;與模型組相比,漢黃芩素干預(yù)組能夠明顯降低巨噬細(xì)胞NO、MDA的分泌水平(表2)。
2.2漢黃芩素能促進(jìn)LPS和ATP聯(lián)合刺激的巨噬細(xì)胞Nrf2 mRNA表達(dá)及胞內(nèi)Nrf2的核轉(zhuǎn)位 熒光定量PCR結(jié)果顯示,與對照組相比,模型組(LPS-ATP聯(lián)合刺激)巨噬細(xì)胞中的Nrf2 mRNA水平顯著升高;與模型組相比,漢黃芩素低劑量干預(yù)組(14 μmol/L)巨噬細(xì)胞中的Nrf2 mRNA水平明顯升高(圖1A)。免疫熒光法觀察巨噬細(xì)胞內(nèi)Nrf2核轉(zhuǎn)位發(fā)現(xiàn),對照組巨噬細(xì)胞內(nèi)Nrf2幾乎不表達(dá),模型組Nrf2明顯表達(dá),但多在胞漿內(nèi),漢黃芩素干預(yù)組巨噬細(xì)胞內(nèi)Nrf2明顯表達(dá),且細(xì)胞核內(nèi)的Nrf2表達(dá)量較模型組明顯增多(圖1B、C)。
2.3漢黃芩素能夠上調(diào)LPS和ATP聯(lián)合刺激的巨噬細(xì)胞抗氧化酶HO-1、SOD、GPx、CAT以及NQO-1的mRNA水平 熒光定量PCR結(jié)果顯示,與對照組相比,模型組巨噬細(xì)胞中抗氧化酶HO-1、SOD、GPx、CAT、NQO-1的mRNA水平顯著降低。在LPS-ATP聯(lián)合刺激時(shí)加入漢黃芩素干預(yù),細(xì)胞中這些抗氧化酶的mRNA水平顯著升高(圖2)。
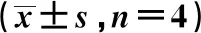
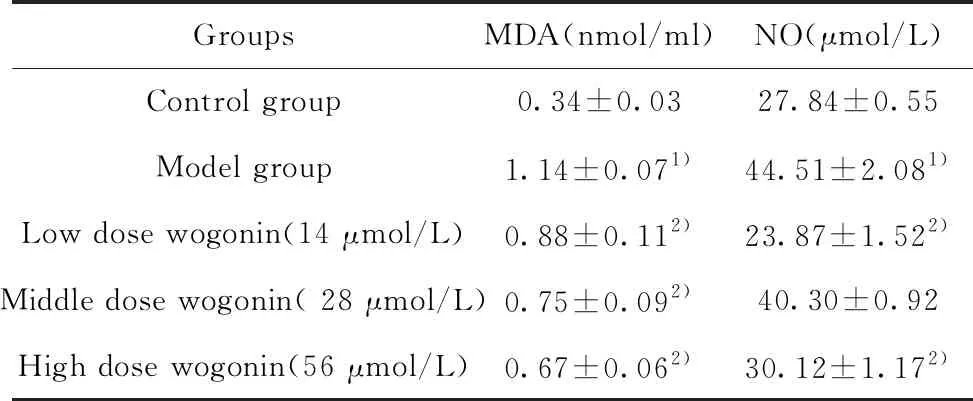
GroupsMDA(nmol/ml)NO(μmol/L)Control group0.34±0.0327.84±0.55Model group1.14±0.071)44.51±2.081)Low dose wogonin(14 μmol/L)0.88±0.112)23.87±1.522)Middle dose wogonin( 28 μmol/L)0.75±0.092)40.30±0.92High dose wogonin(56 μmol/L)0.67±0.062)30.12±1.172)
Note:Compared to Control group,1)P<0.01;compared to Model group,2)P<0.01.
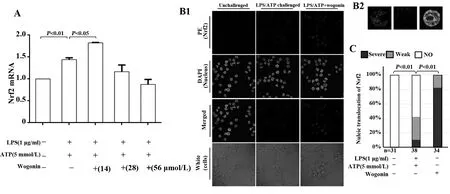
圖1 漢黃芩素對LPS和ATP刺激的巨噬細(xì)胞(RAW264.7)Nrf2 mRNA水平及Nrf2核轉(zhuǎn)位的影響

圖2 漢黃芩素對LPS和ATP聯(lián)合刺激的RAW264.7細(xì)胞抗氧化酶mRNA水平的影響
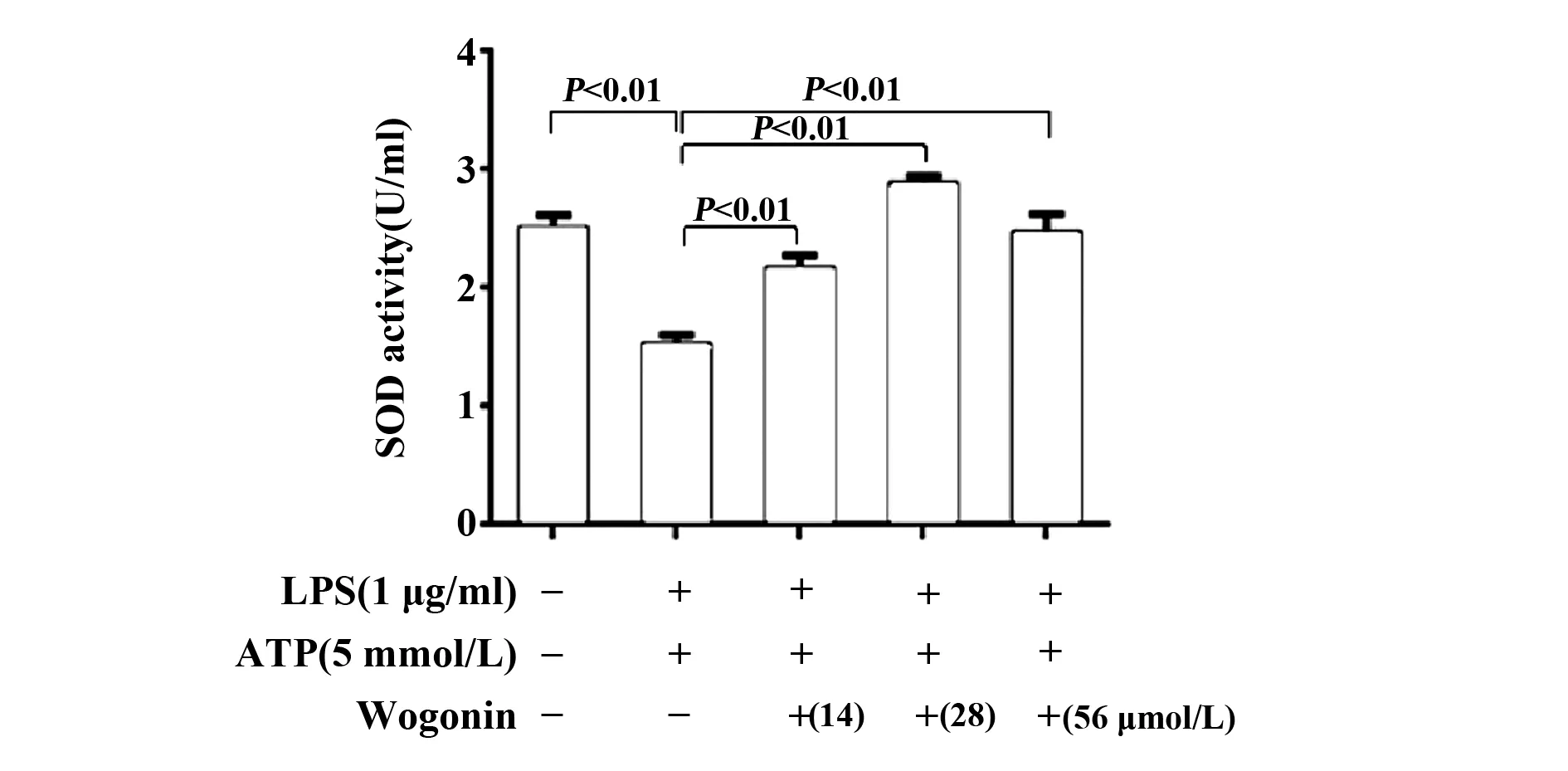
圖3 漢黃芩素對LPS和ATP聯(lián)合刺激的RAW264.7細(xì)胞SOD活性的影響
2.4漢黃芩素能明顯提高LPS和ATP聯(lián)合刺激的巨噬細(xì)胞SOD活性 模型組(LPS-ATP聯(lián)合刺激)與對照組相比,SOD活性顯著降低;與模型組相比,漢黃芩素干預(yù)組能顯著逆轉(zhuǎn)SOD活性被抑制的情況(圖3)。
3 討論
炎癥是機(jī)體對各種致炎因素引起組織損害而產(chǎn)生的一種基本病理過程,也是許多疾病的重要發(fā)生機(jī)制。適當(dāng)?shù)难装Y反應(yīng)作為機(jī)體的一種自我保護(hù)機(jī)制是有益的,但過度炎癥反應(yīng)則會(huì)造成組織器官嚴(yán)重的病理損傷。我們課題組的前期研究表明,漢黃芩素能顯著抑制LPS和ATP聯(lián)合誘導(dǎo)的巨噬細(xì)胞RAW264.7的炎癥反應(yīng),能明顯降低胞內(nèi)IL-1β、IL-18、IL-6、TNF-α、NLRP3和Caspase-1的mRNA水平,減少NF-κB核轉(zhuǎn)位,并能下調(diào)胞外TNF-α和胞內(nèi)ROS水平[6]。ROS又稱活性氧,其產(chǎn)生與炎癥密切相關(guān)。有研究表明,ROS可以激活巨噬細(xì)胞的核轉(zhuǎn)錄因子NF-κB核轉(zhuǎn)位,誘導(dǎo)炎癥反應(yīng)[7]。還有研究表明,細(xì)胞應(yīng)激后ROS濃度增加導(dǎo)致硫氧還蛋白(TXNIP也稱為VDUP1)從氧化硫氧還蛋白-1(Trx-1)中解離,TXNIP隨后與NLRP3相結(jié)合并激活NLRP3炎性小體[9]。這些都說明漢黃芩素對巨噬細(xì)胞炎癥的抑制作用可能與其下調(diào)胞內(nèi)ROS水平有關(guān),那么漢黃芩素又是通過什么機(jī)制下調(diào)胞內(nèi)ROS水平的呢?眾所周知,ROS是氧化應(yīng)激的重要產(chǎn)物,而氧化應(yīng)激的發(fā)生是由于機(jī)體或細(xì)胞遭受內(nèi)源性或外源性的有害物質(zhì)刺激后,產(chǎn)生大量的自由基,釋放出過多的活性氧簇(ROS)、活性氮簇(RNS)等氧化物質(zhì),而機(jī)體或細(xì)胞本身的抗氧化能力下降,機(jī)體的氧化反應(yīng)大大超出抗氧化的清除能力,導(dǎo)致大量積累的 ROS等[11]氧化物不能被清除,從而誘導(dǎo)細(xì)胞損傷,并促進(jìn)炎癥反應(yīng)發(fā)生[10]。反過來,炎癥細(xì)胞在炎癥部位釋放出許多活性物質(zhì),也會(huì)導(dǎo)致氧化應(yīng)激加劇。薛嘉虹等[11]研究表明,炎癥因子 TNF-α 可增強(qiáng) NADPH 氧化酶的表達(dá),并抑制抗氧化酶SOD的表達(dá),導(dǎo)致氧化應(yīng)激反應(yīng)的發(fā)生,而抑制 TNF-α 的表達(dá),可有效地抑制氧化應(yīng)激反應(yīng)。由此可見,氧化應(yīng)激和炎癥是密切相關(guān)的病理生理過程,常相伴而生。據(jù)此,本研究以LPS和ATP聯(lián)合刺激的RAW264.7小鼠巨噬細(xì)胞為炎癥細(xì)胞模型,從抗氧化應(yīng)激的角度探究漢黃芩素下調(diào)ROS水平的作用機(jī)制,從而證明漢黃芩素能否通過抑制氧化應(yīng)激下調(diào)ROS水平,進(jìn)而抑制NF-κB核轉(zhuǎn)位,減少NLRP3活化,最終抑制炎癥反應(yīng)的發(fā)生。
NO是活性氮簇(RNS)中的一種,是氧化應(yīng)激的重要產(chǎn)物,其分泌水平能反映氧化應(yīng)激的程度。氧化應(yīng)激生成過多的ROS會(huì)引起細(xì)胞發(fā)生過氧化脂質(zhì)反應(yīng),使細(xì)胞膜受到損傷,破壞細(xì)胞的功能和結(jié)構(gòu),而丙二醇(MDA)作為過氧化脂質(zhì)產(chǎn)生的終末產(chǎn)物是公認(rèn)的氧化應(yīng)激標(biāo)志物。本實(shí)驗(yàn)結(jié)果發(fā)現(xiàn),與對照組相比,模型組巨噬細(xì)胞NO、MDA的分泌水平顯著提高,而漢黃芩素干預(yù)組相對于模型組其細(xì)胞NO、MDA的分泌水平顯著降低,說明炎癥反應(yīng)能伴隨氧化應(yīng)激的發(fā)生,漢黃芩素能抑制氧化應(yīng)激的發(fā)生。
Nrf2信號通路是細(xì)胞內(nèi)氧化應(yīng)激最重要的防御系統(tǒng)之一,當(dāng)細(xì)胞發(fā)生氧化應(yīng)激,Nrf2 信號通路首先被激活,Nrf2因子核轉(zhuǎn)位與抗氧化反應(yīng)元件(anti-oxidant response element,ARE)上的DNA 基序(GCTGAGTCA)識(shí)別并結(jié)合,啟動(dòng)抗氧化基因的轉(zhuǎn)錄,進(jìn)而引發(fā)抗氧化物及相關(guān)酶類的大量表達(dá),包括 NADPH、 NQO-1、 HO-1、 SOD、 GPx 和 GST 等,以抵御氧化應(yīng)激引發(fā)的細(xì)胞損傷[7]。本實(shí)驗(yàn)結(jié)果發(fā)現(xiàn),與模型組相比,漢黃芩素干預(yù)組能顯著提高巨噬細(xì)胞內(nèi)Nrf2 mRNA水平,促進(jìn)Nrf2的核轉(zhuǎn)位,并能顯著上調(diào)抗氧化酶HO-1、SOD、GPx、CAT、NQO-1的mRNA水平,增加SOD活性,說明漢黃芩素可激活轉(zhuǎn)錄因子Nrf2進(jìn)行核轉(zhuǎn)位,啟動(dòng)抗氧化酶基因的轉(zhuǎn)錄,促進(jìn)抗氧化酶蛋白的表達(dá),從而發(fā)揮抗氧化作用。
綜上所述,LPS與ATP聯(lián)合刺激巨噬細(xì)胞能夠引起細(xì)胞的氧化應(yīng)激,漢黃芩素可能通過激活Nrf2核轉(zhuǎn)位,啟動(dòng)抗氧化酶基因的轉(zhuǎn)錄,誘導(dǎo)抗氧化酶的表達(dá),清除過量的自由基,包括ROS、NO等,繼而抑制NF-κB的核轉(zhuǎn)位,減少NLRP3炎性體活化,從而發(fā)揮抑制炎癥反應(yīng)的作用。本研究在一定程度上豐富了漢黃芩素抗炎抗氧化的作用機(jī)制,然而,其抗氧化的具體分子機(jī)制,如漢黃芩素是如何激活Nrf2核轉(zhuǎn)位的,對Nrf2-ARE信號通路上的關(guān)鍵信號分子有何影響仍有待進(jìn)一步的研究。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
美與時(shí)代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年10期)2020-11-26 08:24:50
世界科學(xué)技術(shù)-中醫(yī)藥現(xiàn)代化(2020年2期)2020-07-25 02:05:56
數(shù)學(xué)物理學(xué)報(bào)(2020年2期)2020-06-02 11:29:24
人大建設(shè)(2019年12期)2019-05-21 02:55:32
光學(xué)精密工程(2016年6期)2016-11-07 09:07:19
西南軍醫(yī)(2016年6期)2016-01-23 02:21:19
西南軍醫(yī)(2015年2期)2015-01-22 09:09:37
食品科學(xué)(2013年15期)2013-03-11 18:25:48