神經科學視角下小腦功能研究新進展及其對運動技能學習與表現的啟示
2020-03-21 08:32:44劉展
中國體育科技 2020年1期
劉 展
近年來,腦成像技術和計算機科學的革命性進步使研究人員能夠以不尋常的方式深入研究小腦的結構、神經元活動,以及對小腦疾病干預后的功能變化。這些新技術包括功能核磁共振成像(functional magnetic resonance imaging,fMRI)、彌散張量成像(diffusion tensor Imaging,DTI)、超分辨率徑跡密度成像(super resolution track-density imaging,TDI)、多光子成像( multi-photon microscopy,MPM)、激光雙光子顯微鏡(two-photon laser scanning microscopy,LSM)和功能性近紅外光譜技術(functional near infrared spectroscopy,fNIRS)等。新技術的使用促進了小腦研究的新成果和新發現。例如,fMRI結果顯示,小腦中網絡的大小是大腦皮層中網絡大小的兩倍。涉及高級思維過程的額頂神經網絡所占的小腦比例比大腦皮層的比例更大(Marek et al.,2018)。這一發現暗示了小腦在高級認知中的作用。fMRI是一種掃描腦部和身體的成像技術。由于腦細胞消耗的能量與血流變化有關,當人腦某個區域使用時,流向該區域的血液也會增加。因此,基于腦血流和神經元激活耦合的事實,fMRI可通過檢測血流變化來測量腦部活動(Skipper et al.,2014)。fMRI也可用于繪制腦部或脊髓的神經活動圖,從而開展對人類或動物的研究。另一種新興成像技術是DTI,也被稱為彌散張量成像核磁共振(DT-MRI)技術,是一種基于磁共振成像的神經影像技術。其主要機制是依賴于平行束中軸突的結構及其髓鞘屏蔽,促進水分子沿其主要方向彌散的原理,通過測量彌散量來比較白質纖維束中平行于軸突和垂直于軸突纖維的方向,并進行顏色編碼,繪制出各區域的位置、方向和各向異性的圖像(圖1)。DTI被視為細胞結構的敏感探針,允許直接體內檢查組織的微觀結構。該技術利用彌散各向異性來提供大腦細節,可用于識別大腦的主要白質區,估算大腦白質神經纖維束的位置、方向和各向異性(Manenti et al.,2007),有助于研究和揭示腦不同區域之間的連接,掃描和診斷深層的腦結構變化,以及評估許多疾病過程(包括創傷和腦瘤)對大腦的損害(Basser et al.,2000;Charlton et al.,2008;Karavasilis et al.,2019)。超分辨率徑跡密度成像(super resolution track-density imaging,TDI)作為一種新穎的腦成像技術,被廣泛應用于神經科學和臨床研究中。TDI可以產生高質量和高清晰度的腦白質圖像,具有高空間分辨率和出色的解剖對比度(Calamante et al.,2011),有助于對白質在腦和精神疾病中的作用進行深入研究(Dai et al.,2018)。
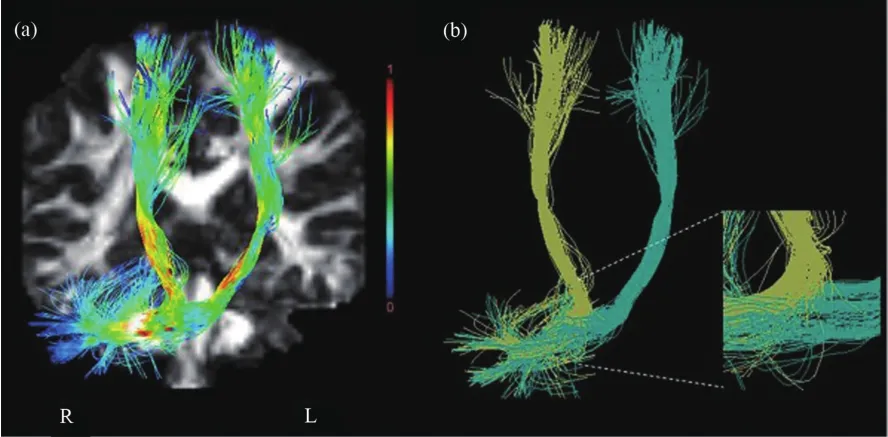
圖1 皮層-小腦通道彌散張量成像(Karavasilis et al.,2019)Figure 1. Diffusion Tensor Imaging of Cerebro-Cerebellar Pathways
實驗室的研究結果和臨床的觀察數據,使人們越來越意識到小腦系統可能參與多種類型的學習以及情緒反應,在身體運動、認知、情感、語言和社交方面均發揮重要的作用(羅程 等,2017;Stoodley,2012;Stoodley et al.,2009)。本文試圖通過對神經科學研究文獻的查閱和結果分析,介紹小腦的基本結構和功能,總結近年來有關小腦運動與非運動功能研究的新發現,并闡述這些小腦功能新發現在運動技能學習應用中的啟示。
1 小腦的基本結構與功能
人腦由大腦、小腦、間腦和腦干組成,共有860億多個神經元(細胞),不均衡地分布在大腦皮層、小腦和皮層下核(Azevedo et al.,2009)。由于大腦皮層含有豐富的較大神經元、樹突、軸、突觸、神經膠質細胞和龐大的貫穿腦白質的軸突分布,盡管其體積約占人腦總體積的82%,但大腦皮層只含有160億個神經元,約占人腦神經元總數的19%。小腦位于顱骨的底部,僅占人腦總體積的10%,但卻容納人腦總神經元的80%,約690億個神經元(圖2),其中大部分神經元是微小的顆粒細胞(Lent et al.,2012)。小腦由3個不同的功能葉組成:絨球小結葉、前葉和后葉。絨球小結葉(flocculonodular lobe)被稱為“前庭小腦”或“原小腦”,主要處理來自前庭系統的信息,調節軀體平衡。前葉(anterior lobe)也被稱為“脊髓小腦”或“舊小腦”,占據了頂骨區(小腦的中央)和中間區。前葉負責處理來自頭部、頸部、上肢、軀干和下肢的感覺信息,主要功能是抑制和調節肌緊張。后葉(posterior lobe)也被稱為“新小腦”,占據小腦的外側半球。后葉負責處理來自脊髓和上位腦結構傳入的信息,調節身體運動的計劃和起始,協調隨意運動。小腦的神經元和組織密集,小腦神經元是通過電信號傳輸信息的專用細胞,并且細胞結構均勻,具有無氧糖酵解和適應性可塑性的特征(Van Essen et al.,2018)。
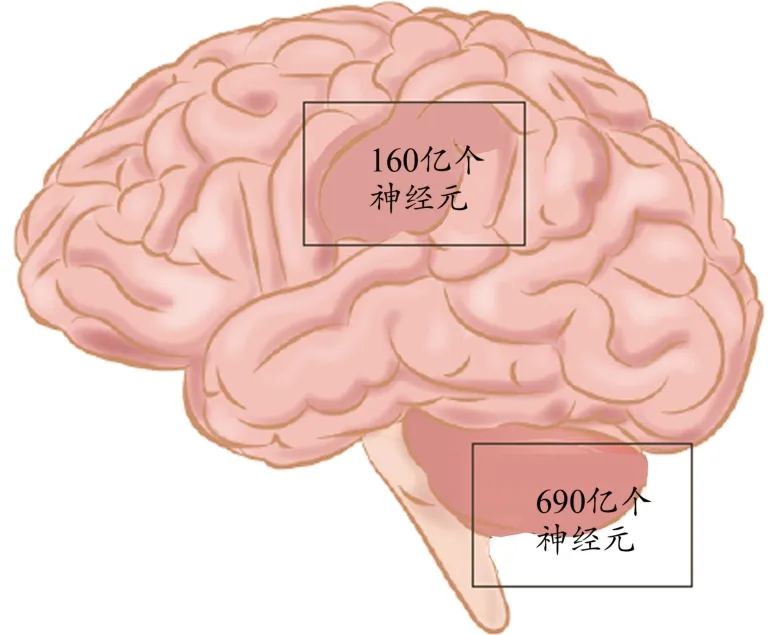
圖2 小腦中與大腦皮層中的神經元數量(Lent et al.,2012)Figure 2. The Number of Neurons in Cerebellum and Cerebral Cortex
小腦最著名的功能是協調身體活動和學習新的運動技能。小腦從感覺系統、脊髓和大腦的其他部位接收信息,然后調節和控制諸如姿勢、平衡、言語之類的自愿運動。小腦還可調節平衡,協調精細動作的完成,使人們能夠快速而平穩地移動。雖然小腦不被認為可以引發或啟動身體運動,但其有助于組織參與特定運動的肌肉群的所有動作,以確保身體能夠平衡,保持一定的體姿,進而產生流暢、協調的運動。一個簡單的例子為大腦的運動皮層告訴人的腿開始行走,而小腦使其步幅平穩、速度均衡。小腦損害可能會導致身體不平衡,運動緩慢和顫抖,甚至中斷。此外小腦還參與順序性、適應性以及粗大和精細運動技能的學習(Buckner et al.,2013),在運動技能學習中起著至關重要的作用。當一個人學習騎自行車或打羽毛球等新運動技能時,通常會經歷反復嘗試-錯誤的過程。小腦通過不斷地調控身體運動,協調粗大和精細動作,最終實現完美地執行動作,這種運動學習是小腦與大腦不同區域結構可塑性變化以及功能上連接增強的結果。需要特別注意的是,小腦與同側身體功能有關。因此,無論何時小腦連接到具有交叉功能的大腦部分(如大腦皮層),連接都必須交叉。前庭小腦即絨球小結葉是前庭系統的重要調節器。前庭系統的損害會導致眩暈和眼球震顫。然而,前庭小腦最重要的功能是允許適應前庭損傷。如果前庭小腦也受傷,則不會發生此補償。前庭眼反射一直是小腦可塑性研究的基礎,因為這種反射在運動過程中保持眼睛穩定并適應前庭損傷方面具有精確作用(James,1971)。
2 小腦運動功能研究新發現
2.1 小腦的同側遷移效果
神經科學的研究已表明,小腦參與程序性工作記憶和運動記憶。這些記憶功能都是空間事項信息處理和程序性運動學習的重要特征(Leggio et al.,1999;Mandolesi et al.,2011;Spampinato et al.,2017a)。眾所周知,小腦在適應性運動技能學習中起著重要的作用。然而,小腦與大腦運動皮層(Motor Cortex-M1)之間的連接是確保正常身體運動的關鍵。小腦的齒狀核通過腹側丘腦以興奮性突觸投射至大腦運動皮層,小腦通過齒狀核丘腦皮質回路來降低大腦運動皮層的活性,被稱為小腦抑制(Spampinato et al.,2017b)。Spampinato 等(2017b)對小腦在適應性視覺運動學習時的抑制變化,以及這種變化是否是皮層特定區域性的進行了創新研究。經顱磁刺激(TMS)技術常用于研究人們學習新運動技能時腦中神經連接的變化。在Spampinato等研究中,TMS技術被用來測量從小腦到齒狀核的抑制性投射。該研究共招募了32名志愿者作為被試,志愿者分別單獨坐在計算機屏幕前,學習操作綁在右手中指的無線控制電子筆來完成一系列需要眼手協調配合的運動任務。研究發現,當被試學習這種用手移動的視覺運動任務時,小腦的抑制水平發生了改變。有趣的是,測量同側未進行視覺運動的小腿肌肉(脛骨前肌肉)的小腦活動情況,驚奇地觀察到了類似的小腦抑制變化。也就是說,這種小腦抑制變化效應還遷移到了沒有進行過學習和訓練的同側小腿肌肉上,用手進行的適應性學習不僅會影響訓練手的小腦-大腦運動皮層的連接,還會影響到未經過訓練的腿部肌肉的小腦-大腦運動皮層的連接。該研究結果表明,同側身體的運動效應器(肌肉)之間存在學習遷移的現象,小腦可以將運動技能學習的效果從身體的一部分轉移到另一部分。該研究結果為小腦與大腦運動皮層之間存在密切的聯系和可塑性提供了直接的證據。
值得注意的是,大腦對身體運動的控制是交叉性的,即大腦的左半球運動皮層控制著身體右側的運動。而Spampinato等(2017b)的研究結果表明,小腦對身體運動的控制則是同側的,即小腦的左半球控制身體左側的運動,與大腦對身體運動的交叉控制截然相反。該研究還調查了小腦變化是否是學習新運動技能時所特有的,測量小腦與大腦運動皮層之間的連接發現,執行熟悉的視覺運動任務時,小腦-大腦的連接活動并沒有發生改變,而學習新任務時卻發生了明顯變化。解釋其中原因,Spampinato等認為,學習新的運動技能時需要小腦與大腦之間更多的溝通與聯系,而在熟練掌握這些運動技能后,人腦會以更加簡單和有效的方式進行操作和控制,更依賴于儲存在小腦中的運動記憶來完成運動技能。另外,該研究后續對只進行手部和腿部動作預備時的小腦抑制水平變化進行了檢測,發現手部和腿部在動作預備期間的小腦抑制水平也發生了變化。該研究結果顯示,與運動學習有關的小腦抑制變化具有皮層特定區域性。前期神經影像學研究結果證明,小腦皮層具有調控上肢和下肢表達粗大運動的特定區域(Buckner et al.,2011;Stoodley et al.,2009)。該研究結果表明,小腦皮層具有調控上肢精細運動的特定區域。Spampinato等認為,小腦的不同部位專門對應特定的身體部位,遵循小腦皮層與執行器之間的對應關系。這項研究還表明,在運動學習出現和未出現的情況下小腦與大腦運動皮層M1連接反應的調控具有皮層特定區域性。這意味著對身體不同部位活動的控制不僅是大腦皮層特定區域的功能,而且也是小腦與大腦運動皮層連接反應調控的結果,反映了皮層特定區域的可塑性機制。而運動前的小腦抑制可能是由于浦肯野細胞和深部小腦核的活動模式所引起的(Celnik,2015)。在預備特定的肌肉運動時,小腦皮層可能通過小腦-丘腦-大腦運動皮層通路來降低其對大腦運動皮層的抑制作用,使大腦運動皮層在運動預備時興奮性增強。這些發現顯示,小腦的興奮性以及與大腦運動皮層的連接的改變與運動學習有關,而小腦在運動學習時發生的抑制性變化則表明了小腦可塑性的改變。已有研究結果表明,小腦皮層中苔狀纖維和中間神經元具有長時程增強(long term potentiation,LTP),而平行纖維和浦肯野細胞具有長時程抑制(long term depression,LTD),這兩種突觸可塑性對運動技能學習非常重要(D’Angelo,2005;Grasselli et al.,2014)。在運動學習中,小腦的突觸可塑性變化對運動行為的改變起到一定的作用。任何未經訓練效應器的小腦抑制變化都可能與運動學習遷移有關。
2.2 小腦顆粒細胞參與運動學習的新發現
小腦顆粒細胞形成小腦皮層的顆粒層,是腦中最小的神經元之一,也是數量最多的神經元。顆粒細胞從苔狀纖維接收信息傳入,并對信息進行組合編碼。傳統理論認為,由于每個顆粒細胞僅接收4~5個苔狀纖維所輸入的信息,顆粒細胞需要多個苔狀纖維輸入處于活躍狀態才會作出響應,并把組合編碼的信息通過輸出神經元——浦肯野細胞發送到小腦的其他部位,后者將信息處理后再發送到大腦的其他部位,包括支配和控制身體運動的運動皮層神經元(Albus,1971;Marr,1969)。根據這種理論,小腦顆粒細胞存在特定的輸入模式,其活動是稀疏而非集中群體性的,并且只對從外部世界收集的信息進行編碼。
普林斯頓神經科學研究所和貝勒醫學院的研究人員使用先進的激光雙光子顯微鏡(LMS)成像技術來解密小腦中的顆粒細胞信號(Giovannucci et al.,2017)。LMS成像技術是一種熒光成像技術,可以在常規(單光子)熒光或共聚焦顯微鏡無法實現的深度,對活組織進行更清晰、更深層次的觀察,從而使得能夠對較厚的組織標本(例如腦片、腫瘤、淋巴結外植體)以及小動物模型的活體進行觀察和研究。它依賴于雙光子吸收原理,可以產生針對單個突觸或突觸組的刺激模式,因此可用來研究神經連接和信號傳導。該研究是第一個探索活體動物學習新任務時小腦顆粒細胞活動的研究。研究人員使用LMS捕獲了小腦中單個顆粒細胞的圖像,并觀察到在學習新運動任務中密集的顆粒細胞群體被激活,而且隨著訓練的繼續,被激活的顆粒細胞數量也在增加。研究發現,在學習新任務時小腦中許多顆粒細胞同時活躍,具有整體活性模式,并且小腦顆粒細胞既對外部感覺信息進行編碼,又對來自大腦其他部位的信息進行編碼,不僅產生預測,而且在接受預測,它們與大腦其他區域保持持續的聯系與溝通,不斷處理信息并隨時更新信息。這項新發現與傳統的小腦功能理論相反,揭示曾經被認為只能控制和調節身體運動的小腦,實際上在學習新事物和技能的認知過程中起著復雜和重要的作用。
3 小腦非運動功能研究新發現
長期以來,人們認為小腦與腦的其余部分尤其是大腦幾乎是分開的,小腦控制和調節與認知無關的無意識或潛意識自動化身體運動,例如本體感覺、身體平衡和動作協調等。然而自20世紀80年代后期以來,研究證據表明,小腦與多種非運動功能有關,包括認知、情感和社交等(馬碧 等,2013;彭洪衛 等,2015;Carta et al.,2019;Schmahmann et al.,1998;Stoodley,2012;Stoodley et al.,2009;Swenson,2006)。
分析人腦不同區域的發育時間與發展軌跡發現,其他腦區域的發育窗口跨越數周或數月,如參與高級大腦功能(認知,感官知覺和語言等)的新皮層細胞,當胎兒仍在子宮中妊娠時大部分已形成。與之不同的是,小腦的發育時間表較長,小腦的浦肯野細胞(Purkinje Cells)在妊娠第7周初開始與干細胞前體分化,但新細胞直到嬰兒快1歲才出現。這種較長的成熟窗口使小腦與整個腦的其他區域建立了完整的聯系,例如與大腦皮層的廣泛連接,而大腦的皮層在感知、注意力、驚醒、思想、記憶、語言和意識中起著關鍵的作用,因此擴大了對神經系統疾病易感性的范圍(Dobbing,1975;Sathyanesan et al.,2019)。Sathyanesan等(2019)認為,發育周期長和與整個腦區域的廣泛連接,這兩個特征是小腦參與多種神經發育障礙的根源。
3.1 小腦認知研究新進展
3.1.1 新的大腦圖暗示了小腦在認知和語言中的作用
近年,華盛頓大學Marek等(2018)使用一種特殊的“精確映射”的fMRI技術來研究腦連接,繪制了人類的神經網絡圖。該技術能夠量化小腦與其他腦區域之間的各種連接。他們發現,只有20%的小腦與身體運動有關的大腦區域相連接,而80%的小腦則與抽象思維、計劃、情感、記憶和語言等功能有關的大腦區域相連接。該項對腦部掃描的新分析,突出顯示了小腦神經元網絡水平上的變化,以及小腦區域與大腦區域活動的程度。基于這些相關性,研究人員將小腦區域分配給各種神經網絡,包括背側注意力網絡,默認模式網絡以及控制手、臉和腳的神經網絡。評估這些網絡在每個人中的相對規模和位置,他們發現小腦網絡組織存在較大的個體差異。另外,結果還揭示,小腦中網絡的大小是大腦皮層中網絡大小的兩倍,而且涉及高級思維過程的額頂神經網絡所占的小腦比例(23%)比大腦皮層的比例(10%)更大,表明小腦在高級認知尤其是對人們的思想有質量控制的作用。研究小組還分析了掃描腦活動的相對時間,發現小腦網絡中的信號比大腦皮層中的信號落后125~380 ms,表明小腦處理源自大腦皮層的信號,以便監督大腦的活動。研究認為,小腦不會直接執行諸如思考的任務,就像它不會直接控制運動一樣。它的主要功能在于監視正在工作的大腦區域,使它們表現地更好更有效。這種功能類似是一種編輯,可以不斷地審查和改善一個人的思想和決策,使運動控制、認知、情感、社會行為等過程更順暢、更快、更準確。小腦會自動執行和監控這些操作,從而使有意識的大腦專注于更重要的事情。
3.1.2 小腦參與視覺感知的調控
長期以來,人們認為視覺知覺完全屬于大腦的功能,大腦皮層和皮層下結構負責視覺感知信息的處理與分析,這種功能是以視網膜與大腦特定區域的連接而進行的。然而,有研究表明,大腦皮層的視覺區域與小腦在功能上緊密連接(Brissenden,et al.,2016;Buckner et al.,2011)。為了進一步確定小腦與視覺的關系,荷蘭研究人員對小腦的視覺空間功能區域圖進行了研究,發現小腦與視覺感知密切相關(van Es et al.,2019)。研究人員使用fMRI技術發現,在小腦的感覺運動區域,其功能通過身體同側特征而表現出來,小腦包含5種同側視覺空間功能區,對視覺感知有強烈的反應。研究人員還量化了視野覆蓋和功能區域分布。結果顯示,在眼球運動區域,認知功能例如注意力、工作記憶以及視覺運動信息的整合效應分別位于小腦的3個不同區域,而且小腦含有豐富的視覺空間信息,并可根據已知的視網膜特性對這些信息進行組織與處理。這項研究也揭示了小腦在視覺空間信號的選擇性方面起著一定的作用。結合先前的研究,表明小腦對于認知功能(例如感知和記憶)至關重要。當小腦不能發揮正常作用時,可能導致腦部疾病或功能障礙(Marek et al.,2018;Wang et al.,2014)。因此,要了解腦和治愈腦部疾病,還需要檢查小腦的功能。
3.1.3 小腦參與獎賞期望的編碼和調控
Strick等(2009)首次證明了小腦有助于控制大腦的獎賞回路。該研究的目的在于探究小腦與大腦腹側被蓋區(VTA)是否存在直接聯系。大腦腹側被蓋區長期以來被認為參與獎賞處理和編碼,是控制獎賞和激勵行為的大腦結構。一旦確定了小腦與大腦腹側被蓋區之間的神經聯系,便可以研究小腦到大腦腹側被蓋區的輸入是否能夠影響獎賞和社會偏好行為。研究人員使用光遺傳技術,激活連接到大腦腹側被蓋區域的小腦神經元發現,當小鼠按下操縱桿時,會激活一些顆粒細胞。當小鼠等待或獲得獎賞時,另一組顆粒細胞會被激活。而當取消獎賞時,小腦中又另一組顆粒細胞被激活。激活小腦神經元導致大腦腹側被蓋區域的激活增加,表明這兩個腦結構之間存在直接和有效的神經連接。
Wagner等(2017)使用雙光子鈣成像(two-photo calcium imaging)技術實時記錄神經元的活動發現,小腦內的特定顆粒細胞會學習并響應預期的獎賞或缺乏獎賞。Wagner等開始這項實驗時,最初只是期望發現顆粒細胞的活動與計劃和執行身體運動有關。同Strick等(2009)的研究結果相似,該研究發現,當小鼠獲得期望的含糖獎賞時,一些顆粒細胞被激活和發射信號。令人驚訝的是,當小鼠等待它所期望的含糖獎賞時,其他一些顆粒細胞被激活。而當研究人員悄悄地拿走了預期的含糖獎賞時,小腦又激活了另一批顆粒細胞。對幾天學習過程中相同的顆粒細胞進行跟蹤觀察,發現具有獎賞預期反應的顆粒細胞從學習開始時就對獎賞做出反應,對獎賞缺失反應隨著學習的進行而變得越來越強。再一次證實,除了協調和控制身體運動,小腦也有助于控制大腦的獎賞回路。
Kostadinov等(2019)在對攀緣纖維輸入浦肯野細胞在產生和評估有關運動動作,以及感覺刺激與獎賞之間的關聯的預測中的作用進行研究發現,攀緣纖維將獎賞和期望信號投射到小腦微小區域,獎賞信號激活了一些微區并抑制了其他微區,而獎賞缺失則激活了獎賞中受抑制的微區。Strick等(2009)認為,小腦以某種方式參與觸發了受到獎賞刺激的大腦區域釋放神經遞質多巴胺,多巴胺被釋放到中腦邊緣途徑中,從而引起愉悅和報酬感。在學習過程中,對可預測獎賞的反應會逐漸受到抑制。因此,小腦在獎賞和社會偏好行為的創建和控制中可能具有主要作用(Kostadinov et al.,2019;Strick et al.,2009,Wagner et al.,2017)。這些研究結果證明,小腦以及小腦-大腦回路在控制獎賞和社會偏好行為方面起著重要的作用,并且對成癮行為的治療和康復提供了新策略。
3.2 小腦與大腦連接研究新發展
3.2.1 攀緣纖維向小腦傳遞感覺信息的新發現
小腦皮層中的浦肯野細胞具有長時程抑制,它可抑制引起錯誤運動的突觸的活動。單個浦肯野細胞上的興奮性突觸中實際上只有不到5%起作用(Bloedel et al.,1983;Ito et al.,1982b)。需要注意的是,在小腦所有類型細胞中,只有浦肯野細胞發出軸突離開小腦皮層,是小腦皮層中唯一的一種傳出神經元。其余類型的細胞僅在小腦皮層內建立連接。進入小腦的軸突有兩種:苔狀纖維和攀緣纖維,這兩種纖維在小腦皮層中具有不同的端接和作用。苔狀纖維末端突觸連接在顆粒細胞樹突上,激發顆粒細胞。攀緣纖維直接在浦肯野細胞上突觸并強烈激發它們(Bloedel et al.,1983;Ito et al.,1982b)。
在日常生活中,源源不斷的感覺信息不斷沖擊和挑戰著人腦。當學習一種新的運動技能時,人腦會不斷跟蹤周圍的感官提示。這些感官提示有助于指導運動技能學習,因此當再次遇到相同的環境時,人們有可能以更好的方式進行運動。已經證實,感覺信息有助于運動技能的學習。然而進入小腦的輸入途徑是影響感覺信息編碼的重要因素(Albus,1971;Ito et al.,1982a;Marr,1969)。通過接收來自大腦各個區域的信息輸入,小腦整合并通過浦肯野細胞發送精確的信息。浦肯野細胞的一個重要輸入是攀緣纖維的遠程投射(Ito,1984;Spampinato et al.,2017b)。
最近,Gaffield等(2019)進行了一項研究來檢驗攀緣纖維對感官信息編碼的影響。該研究認為,攀緣纖維與小腦形成了長距離連接,可以提供指導性的運動信號并傳遞感覺信息。研究對小鼠進行聽覺、視覺和本體感覺刺激的同時,使用雙光子鈣成像技術檢測小腦浦肯野細胞的活性。然后,分離并評估了浦肯野細胞活性的變化,這些變化直接對應于每個刺激和攀緣纖維輸入的時間。研究發現,每當出現一種感覺刺激時,浦肯野氏細胞的活性都會持續增強。但是,浦肯野細胞的這種活性增強作用在3種感覺類型中并不完全相同,它隨著感覺類型的變化而變化。接下來,研究人員又檢查了攀緣纖維以外的其他細胞是否直接促進了浦肯野細胞的活性。使用光抑制神經活動和藥物抑制神經活動技術,抑制局部小腦回路中的其他細胞類型,結果發現,浦肯野細胞的活性增強現象沒有發生變化。但是,直接抑制攀緣纖維的活性后,這種增強作用被消除了。結果表明,只有攀緣纖維負責向小腦傳遞感覺信息。該研究進一步開創新技術監測攀緣纖維軸突的活動發現,當提供感覺刺激時,攀緣纖維的突觸前活動的分級變化可在浦肯野細胞中準確地表示出來。攀緣纖維的活動越強,它所連接的浦肯野細胞的活性越強。傳統上認為,到達小腦的感覺信號是由浦肯野細胞通過局部連接進行整合和處理的(Albus,1971;Marr,1969)。研究表明,攀緣纖維參與感覺信息編碼,并通過浦肯野細胞把感覺信息傳遞給小腦。這意味著大腦的遠端區域對感覺信息進行處理,然后將其直接地傳遞到小腦進行整合,促進小腦調節身體運動和學習新運動行為(Zhang et al.,2019)。
3.2.2 小腦與額葉皮層連接對記憶功能的影響
眾所周知,腦的某些區域負責調控某些功能,例如小腦對控制運動很重要,而額葉皮層負責諸如短期記憶和決策等認知功能。但是,隨著腦中數十億個神經元相互作用謎團的相繼揭開,人們越來越清晰的認識到,腦特定區域的功能并不是簡單的區域劃分和功能界定的。已經確認,小腦與腦其他區域有廣泛的相互連接和回路(圖3)。其中,額葉皮層與小腦在解剖學上是相互連接的,并且臨床上發現小腦損傷引起記憶或計劃受損,也提示額葉皮層與小腦可能存在功能上的連接。
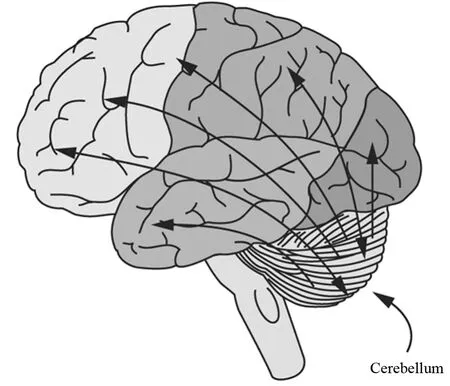
圖3 小腦與其他腦區域之間的回路與連接(Jensen,2005)Figure 3. Circuits and Connections Between Cerebellum and other Brain Regions
Gao等(2018)首次證實,小腦不僅能控制肌肉活動,而且在認知功能中發揮重要作用。研究人員對小鼠進行訓練,在特定位置向小鼠顯示了單個對象。一定時間延遲后,小鼠必須記住該物體的位置,并通過左右滑動來指示其位置。小鼠在執行正確的運動之前必須先使用短期記憶來回憶物體所在的位置。研究發現,在延遲期,額葉皮層的記憶活動可預測小鼠未來的運動,延遲期的記憶活動在額葉皮層和小腦均可觀察到。研究人員在延遲期使小鼠的小腦區域保持沉默,導致了錯誤的反應,但并未干擾身體運動。同時,額葉皮層的記憶活動遭到嚴重影響。然后,他們使額葉皮層區域保持沉默,從而停止了小腦的記憶活動。研究人員發現小腦的輸出是針對額葉皮層,小腦的活動與額葉皮層的記憶活動緊密聯系。當破壞這兩個區域之間的交流時,記憶活動就會被破壞。該研究結果表明,單個行為的活動是由腦多個區域協調的結果。小腦通過從錯誤中學習來指導身體的運動,并參與運動記憶的形成。例如,當我們學習籃球的投籃技能時,最初的準確率較低,但是小腦可以根據視覺信息反饋來調整投籃動作,最終提高投籃的準確性。
3.2.3 圖雷特綜合癥與小腦連接有關的新線索
圖雷特綜合癥是一種神經系統方面的障礙,常見的癥狀包括非自愿運動性抽動,如拍手、眨眼、嗅探、發聲失控等(Worbe et al.,2015)。圖雷特綜合癥的早期癥狀通常首先在兒童時期出現,平均發病年齡為3~9歲。以前人們認為,圖雷特綜合癥運動性抽動是人腦單個功能區域-基底神經節異常或障礙導致的。但是,最近對人、猴子和小鼠的研究表明,小腦、大腦皮層和丘腦也參與了這些抽動的產生(Ito,2001)。 Caligiore等(2017)使用先進的基于計算機的腦模擬技術,對圖雷特綜合癥運動性抽動是否是由于大腦皮層和皮層下區域之間的相互作用引起的進行了研究。計算機模擬發現,基底神經節-小腦-丘腦-大腦皮層系統功能障礙會在大腦中產生運動性抽動(圖4)。圖雷特綜合癥抽動新模型研究結果表明,基底神經節中異常的多巴胺活性與丘腦-大腦皮層系統的活性可能共同觸發了初始抽動。此外,基底神經節與小腦之間的雙向相互作用可能使小腦影響抽動的產生。研究人員認為,該新模型代表了首次計算機模擬嘗試,旨在研究最近發現的基底神經節與小腦結構聯系的作用。研究人員還發現,當連接基底神經節、丘腦、大腦皮層和小腦的神經回路功能異常時,該新計算機模型可用于預測抽動的次數。由此可見,越來越多的研究和臨床證據支持小腦-大腦的連接和相互作用。盡管目前還沒有藥物可以完全消除圖雷特綜合癥患者的癥狀,但認知行為干預法,例如意識訓練、競爭反應訓練、生物反饋療法等,可用于減少抽動和解決繼發性社交和情緒問題。該研究發現為探索更加有效的圖雷特綜合癥治療和康復方法提供了新的思路。
3.2.4 小腦與大腦連接在自閉癥譜系中的作用
自閉癥譜系患者由于腦缺乏有效過濾和調節信息的功能,通常會經歷一個完全未過濾的信息世界,不斷受到各種感覺刺激的沖擊。例如,燈光、聲音和氣味等都可能使患者感到難以忍受,甚至痛苦。自閉癥譜系患者還會在多種環境下表現出持續性的社會溝通和交往缺陷或障礙,并伴有某些重復性的行為、興趣或活動。不論是在身體、心理和社會層面上,自閉癥譜系患者都表現出了異于常人的低功能發展水平。
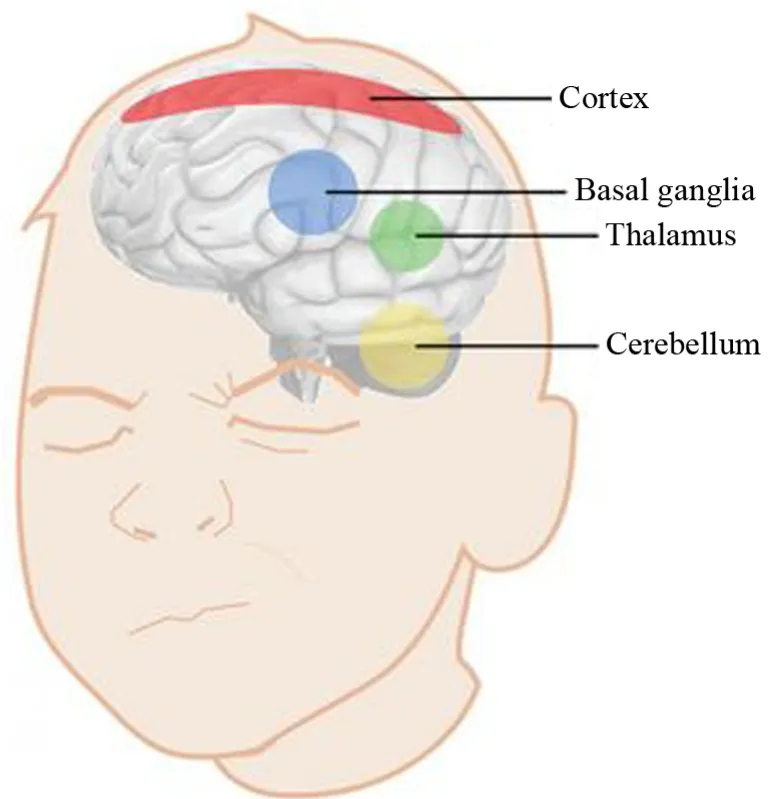
圖4 導致圖雷特抽動產生的關鍵腦區:基底神經節-小腦-丘腦-大腦皮層系統(Caligiore et al.,2017)Figure 4. Key Brain Regions that Cause Tourette’s Twitch:Basal ganglia-Cerebellum-Thalamus-Cortex
越來越多的研究表明,自閉癥譜系癥狀可能歸因于功能連接的變化,或大腦內部區域信息傳遞出現異常(Becker et al.,2013;Nayate et al.,2005)。 Khan 等(2015)對自閉癥譜系患者的大腦與小腦之間的連接進行了研究。使用靜息態功能磁共振成像(rs-fMRI)技術研究小腦和大腦7個不同區域之間的連接,其中4個大腦區域位于感覺運動區域,其他3個是超模態區域,即負責復雜、抽象心理過程的區域,例如語言、解釋社交情景和社會互動。他們對大腦與小腦之間的交叉連接研究發現,自閉癥患者大腦中每個感覺運動區域與小腦的連接都比正常人感覺運動區域與小腦的連接更多。相反,自閉癥譜系患者每個超模態區域與小腦的連接都比正常人較少。研究結果表明,大腦與小腦之間的連接異常可能是導致自閉癥譜系患者身體、心理和社交異常的主要原因,并且自閉癥譜系患者大腦與小腦之間的連接異常不是隨機的。針對交叉連接的測量與觀察也發現,正常人和自閉癥譜系患者的大腦具有相同數量的被認為是“規范”的連接,即大腦與小腦負責相同任務(感覺運動或超模態)的連接區域。然而,自閉癥譜系患者的大腦超模態區域與小腦之間的連接較弱,而大腦感覺運動區域與小腦的連接較強。這意味著自閉癥譜系患者在兩種不同類型的區域之間具有更多的“非規范”聯系。
該研究檢測整個小腦與大腦左、右側所有區域之間的整體連接性,獲得了不同的結果。在感覺運動區域和超模態區域,自閉癥譜系患者的大腦與小腦之間的整體連接更多。這意味著自閉癥譜系患者的同側大腦與小腦之間的聯系要多于正常人。但是,研究確實發現,左側大腦超模態區域與小腦之間具有更多連接的個體,對社會的反應更加強烈。這個結果并不令人感到意外,因為研究人員發現,超模態區域有助于復雜而抽象的思維過程,例如解釋社會狀況。左側大腦感覺運動區域與小腦連接更多的個體,對社會反應的得分較低。右側大腦感覺運動區域與小腦連接更多與較低的非語言智商有關。研究結果還顯示,大腦與小腦的連接與溝通技能水平之間的相關性較低。這項研究表明,大腦與小腦之間的連接確實在自閉癥譜系中起著一定的作用,提供了自閉癥譜系患者行為癥狀與大腦-小腦連接差異有關的證據。小腦連接差異的特異性表明它在自閉癥發展中的獨特作用,值得進一步的研究。由于此研究使用的fMRI技術要求參與者在有限的空間下進行合作,所以很難檢測到自閉癥嚴重程度與大腦連接之間的總體相關性。有研究提出,小腦損傷或功能障礙會干擾大腦其他部位的發育進程,影響對外部刺激進行處理與分析的能力(Wang et al.,2014)。根據該理論,在生長關鍵時期,小腦功能障礙可能會破壞新皮層回路的成熟,導致包括自閉癥譜系在內的認知和行為癥狀。眾所周知,小腦是信息處理器。大腦皮層負責更高程度的信息處理,但不會接收未經過濾的信息。從大腦檢測到外部信息再傳遞到大腦皮層之間,必須采取一些關鍵的過濾步驟。小腦就起著這種關鍵的信息過濾與處理的作用(Danny et al.,2017)。
4 對運動技能學習和表現的啟示
將大腦中的運動“思維”和小腦中的運動“操控”結合起來,通過采用基于這種“上下”模型的雙管齊下的方法,每個人都可以通過針對大腦和小腦的常規訓練來創造更好的運動思維和身體天賦,從而優化自己的行為或運動表現。考慮到大腦與小腦之間的關系,大腦可認為是“人腦上部”,小腦是“人腦下部”,以便對應“左腦-右腦”區域與功能的劃分,從而有助于重塑大腦與小腦的關系。任何一項運動中的成功表現最終都取決于小腦,協調、反射、快速的反應時間以及運動技能學習都依賴于小腦結構。它從脊髓和感覺系統接收信息,并使用此信息來調節和執行身體運動。小腦可以幫助職業羽毛球運動員以高的精準度擊球,并在短短的停頓時間內改變運動程序。
運動學習效果具有一定的遷移性,但遷移的效果因運動任務、技能等的類型而不同(Schmidt et al.,2011)。運動和認知技能的獲得通常取決于練習和使用的時間,以及努力提高的意志,尤其兒童更是如此(Carson et al.,2015)。腦不同的區域功能需要不斷地接受挑戰才能提高,而不是簡單的使用。也就是說,這個提高的過程需要大量的練習,需要有超過現有水平的心態和意志以及不懈的努力(Ericsson et al.,2009)。當學習新動作和技能時,需要大腦和小腦的共同參與。不斷學習新的東西,可以增強大腦與小腦的連接,促進腦功能的發展與提高(Danny et al.,2017)。當接受或學習新的事物時,能夠最大程度啟用大腦的人,就會有最佳的表現(Duncan et al.,2000;Poldrack et al.,2005)。然而當人們熟練掌握相關知識和擅長這種技能后,表現最佳者往往是那些啟用腦前回最少的人。因為此時任務的完成將下放給比腦前回進化更長時間,而且功能完善的腦下回區域包括小腦。這些區域在支配熟練技能完成方面比腦前回效率更高,負責自動化的行為。只有通過重復的行為(練習)才能使這些行為變成自動化行為,而不經過腦前回的控制。這是小腦的作用在學習過程中逐步提升,腦不同區域功能連接效果不斷優化的結果(Basser et al.,2000;Chein et al.,2005;Garavan et al.,2000;Temple et al.,2003)。
以身體活動為基礎的運動技能學習專注于前庭系統、小腦和大腦其余部分之間的相互作用,旨在通過利用人腦的自然生長,發展有效的運動和認知能力來改善人腦不同區域之間的神經元傳遞,從而避免或減少神經疾病與小腦之間的聯系。在設計練習內容和訓練環境時需要考慮動作與認知的有機搭配,并通過科學的手段和先進的器材來促進小腦不斷調節身體運動的速率、節奏、力量和準確性。同時還應根據年齡和運動能力的實際情況,適宜地增加粗大與精細運動技能相結合以及小腦參與調節認知過程的速度和連貫性的練習內容部分。另外,在運動技能學習過程中,除了不斷學習新的動作元素(基本和功能動作與動作模式),還要學習不同的動作技術與完成的方法,并且通過改變學習任務、學習環境以及學習者本身的限制條件來提升運動技能學習的效率(Newell,1986)。運動技能的學習是基于運動技能的元素(動作)、運動技能的完成方法(技術)和運動技能完成的支撐(能力)3者之間的有機搭配和協同發展(Liu,2018)。學習任務的多樣性和練習條件的多變性可增強小腦的運動記憶、動作協調和運動管控的功能,加強小腦與大腦不同區域之間的連接和信息交流,從而提升運動技能的學習效果和表現。根據小腦參與身體同側的感知、運動控制和學習效果遷移的特殊功能,在運動技術和技能學習過程中,應考慮增加身體同側運動的部分——手與腳、手臂與腿,例如橫行移動、跨步、同側弓箭步(羽毛球跨步接球)。還應考慮增加同側上下肢體協調練習和不同身體部位之間的協調練習,例如,用右手拍擊右腳內側或外側,用上肢的一部位觸碰軀干或下肢的某一部位。這些身體動作協調練習,尤其在學齡前和學齡期兒童的運動技能學習和發展中更為需要。
由于小腦發育和發展需要較長的時期,并且參與多種感知信息的過濾與分析,特別是參與對視覺-運動信息的處理。嬰幼兒需要進行適量的平衡、協調、各種視覺-運動的結合練習,例如觸摸和觀察各種顏色、形狀、空間、距離、方向的物體,適度地進行眼-手配合(單、雙手)、眼-腳配合(單、雙腳)練習等。還應為嬰幼兒提供完成多種基本動作模式的機會,例如翻身、爬行、跪立、起身、推、拉、側彎、轉動等(劉展,2016)。學齡前和學齡初期的兒童還要學習身體移動、身體控制和物體操控3種基本動作技能,增強多種感知與身體運動的有機結合,增強小腦對運動記憶、感知信息處理、身體動作協調等的功能參與,提升視覺運動與小腦連接效果,并加強小腦與大腦其他區域的連接和信息流通性。此外,還需要充分利用小腦在多種感知信息整合與處理、信息反饋的功能,安排與實施基于閉環運動控制系統的各種本體感覺和運動覺意識、身體平衡、關節穩定和反應性肌肉收縮的專門練習內容與方法,提高肌肉激活水平、剛韌性和反應性收縮的能力,增強關節在不同受力下的穩定性,建立神經對肌肉有效控制的功能。有組織的運動技能的練習還應逐步增加基于開環運動控制系統的練習內容與方法。由于小腦參與預備性動作和適應性學習,應增加動作預先計劃和預備性動作練習,并通過不斷變化環境和動作程序來增強小腦對身體運動的協調與控制,提升動作的流暢、平穩和準確性。逐步從閉環運動控制為主的身體練習(重復性動作、穩定性環境的肌肉力量、關節幅度以及身體平衡)轉換為以開環運動控制為主的身體練習(多變的動作、速度、力量、方向以及不可預測的運動環境等)。這些對功能動作和動作模式的獲得與發展,基本動作技能和專項運動技能的學習與掌握都非常重要。
5 結束語
綜述小腦功能研究最新發現與進展,小腦像調控肌肉運動一樣調控非運動功能。小腦負責協調身體運動和參與許多認知活動過程。可以通過優化腦區域之間的連接來實現腦功能和人類潛能的最大化。從歷史上看,小腦被認為與運動控制有關。但是,過去30年的研究清楚地表明,小腦還致力于許多認知、語言和社交功能,例如工作記憶、認知控制、動作觀察、獎賞期望等。從簡單的哺乳動物進化到人類的過程中,小腦沒有像其他大腦結構那樣擴大,這暗示著小腦在人類智力發展中的特殊作用(Diedrichsen et al.,2019)。先進的成像技術使研究人員能夠通過小腦神經元的活動、微回路和與大腦不同區域的連接來收集和分析小腦的數據,并以可視化的方法探索和發現小腦的結構與功能。最近,King等(2019)進行了一項大規模研究,通過創建一個多領域任務測試庫,其中包含26個任務,涵蓋人類心理活動的大多數方面,包括語言、記憶、數學、抽象思維、心理意象和運動控制,開發了一個全面的人類小腦新功能圖。該圖以前所未有的細節表征了每個小腦區域的功能。這個新功能圖可用于預測新任務和新參與者的小腦功能邊界,還可以用于研究小腦功能障礙與神經發展性疾病和精神障礙的關系。小腦已成為21世紀認知和運動神經科學、發育神經生物學以及精神病學研究的熱門領域,充分了解小腦在身體運動、認知、語言、社交、情緒調節和精神疾病中的潛在作用,具有深遠的意義,也是優化一個人身體、心智和社交情感功能的關鍵。
猜你喜歡
故事作文·高年級(2023年10期)2023-10-23 11:21:18
中華詩詞(2022年6期)2022-12-31 06:41:24
中國公路(2017年19期)2018-01-23 03:06:33
中國科技論壇(2017年7期)2017-07-25 08:49:53
學苑創造·A版(2017年6期)2017-06-23 14:10:46
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
Coco薇(2015年11期)2015-11-09 13:03:51
電測與儀表(2015年5期)2015-04-09 11:30:52
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
