miR-29c通過調控SIRT1表達介導動脈粥樣硬化氧化應激與炎癥反應
2020-04-01 04:54:10李貞貞唐海濤朱華慶
安徽醫科大學學報 2020年2期
李貞貞,唐海濤,王 怡,周 青,汪 淵,朱華慶
動脈粥樣硬化(atherosclerosis, AS)是一種脂質堆積在動脈壁進而發生粥樣病變并導致血管管腔狹窄的慢性炎癥性疾病,嚴重危害人類健康。AS所致的冠狀動脈疾病、中風、外周動脈疾病等心血管疾病是人類高發病率和死亡率的主要原因[1-2]。沉默信息調節因子2相關酶類1(silent mating type information regulation 2 homolog-1, SIRT1)是一種煙酰胺腺嘌呤二核苷酸依賴性的組蛋白去乙酰化酶。研究[3]表明,SIRT1可通過去乙酰化下游靶分子來調節細胞代謝、減緩衰老,抑制氧化應激,減少炎癥反應。
微RNA(microRNA, miRNA)是一種內源性的、長約22 nt、單鏈非編碼的小RNA,通過與靶基因mRNA的3’-非編碼區(3’-UTR)結合,降解或抑制其翻譯,從而調控基因表達[4]。近年來,越來越多的研究表明miRNA在AS的發生發展過程中起關鍵作用。其中,文獻[5]報道miR-29可參與多種疾病或病理狀態的發生發展過程,并且miR-29c在AS形成中可能發揮多方面的重要作用,但具體的作用機制尚不清楚。因此研究miR-29c在AS形成過程中的作用機制具有重要意義。
1 材料與方法
1.1 主要材料與試劑清潔級雄性C57BL/6J小鼠與C57BL/6J ApoE-/-小鼠各6只,體質量(14±1.8)g,購自北京協和醫學院基礎研究所;人臍靜脈內皮細胞(human umbilical vein endothelial cells,HUVEC)購自美國ATCC細胞庫;DMEM高糖培養基干粉與胎牛血清(FBS)購自美國Gibco公司;二甲亞砜(dimethyl sulfoxide,DMSO)購自美國Sigma公司;氧化型低密度脂蛋白(oxidized low density lipoprotein,oxLDL)購自廣州Yiyuan Biotechnologies;Mimic與TransMessenger Transfection Reagent試劑盒購自德國Qiagen;Antagomir購自上海GenePharma;活性氧(reactive oxygen species, ROS)檢測試劑盒購自上海BestBio公司;TNF-α檢測試劑盒購自美國Adipo Bioscience公司;BCA試劑盒購自上海碧云天生物技術公司;還原型煙酰胺腺嘌呤二核苷酸磷酸(nicotinamide adenine dinucleotide phosphate,NADPH)氧化酶活性化學發光法定量檢測試劑盒購自上海宸功生物技術有限公司;總RNA提取試劑盒購自北京天根生化科技有限公司;逆轉錄試劑盒購自日本Takara公司;一抗SIRT1購自英國Abcam公司,內參甘油醛-3-磷酸脫氫酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)購自美國Santa Cruz公司;二抗購自北京中杉金橋公司。
1.2 主要儀器酶標儀(Thermo MultiskanGO,美國Thermo-Fisher公司);低溫離心機(Sorvall Legend Micro 21R,美國ThermoFisher Scientific);電泳儀(DYY-11型,北京六一儀器廠);-80 ℃冰箱(MDF-U73V,日本sanyo);CO2細胞培養箱(MCO-20AIC,日本Panasonic公司);超凈工作臺(SW-CJ-1F型,蘇州凈化);正置顯微鏡(DM4000B,德國Leica公司);立式壓力蒸汽滅菌器(BXM-30R,上海博訊公司);制冰機(AF100,意大利SCOTSMAN公司);電子天平(DC-120AS,南京蘇測計量儀器有限公司);搖床(TS-1型,海門其林貝爾儀器制造有限公司);全自動數碼凝膠成像系統(ChemiQ 4600,上海歐翔公司);恒溫水浴鍋(DK-8D型,上海躍進醫療器械廠);PCR儀(T-Gradient Thermoblock,德國Biometra)。
1.3 方法
1.3.1建立模型 清潔級ApoE-/-小鼠與C57BL/6J小鼠各6只,常規方法建立AS模型,即小鼠于4周齡斷奶后,采用明暗(12 h ∶12 h),溫度(22±1) ℃,濕度50%~60%條件下以普通飼料(4%脂肪和0%膽固醇)進行適應性飼養,AS組ApoE-/-小鼠于6周齡時采用高脂飼料(21%脂肪和0.15%膽固醇)喂養。以C57BL/6J為正常對照組(正常組),給予普通飼料進行飼養。12周后處死分離主動脈進行相關實驗。
1.3.2細胞的培養與分組 復蘇HUVEC細胞后,以高糖DMEM培養基(含10% FBS、1% 青鏈霉素混合液)于37 ℃、5% CO2恒溫培養箱中培養,當細胞密度達80%時用0.25%胰酶進行消化傳代。采用oxLDL (50 μg/ml)刺激HUVEC細胞進行后續細胞實驗。細胞分組:空載體轉染組(對照組)、Antagomir組、Mimic組、oxLDL組、oxLDL+Antagomir組、oxLDL+Mimic組。
1.3.3細胞轉染miR-29c的過表達與抑制 HUVEC細胞轉染miR-29c mimic和antagomir。轉染前1 d,消化對數生長期HUVEC細胞,以1.0×106~2.0×106/孔接種于6孔板中,加入培養基(含10% FBS),搖勻后放置37 ℃培養箱。根據TransMessenger Transfection Reagent試劑盒說明書進行細胞轉染,3~4 h后換成完全培養基,于CO2恒溫培養箱中繼續培養。
1.3.4ROS檢測 組織ROS檢測:取50 mg動脈,加入1 ml勻漿緩沖液A,置于研磨器中研磨,充分勻漿后于4 ℃離心機中離心(1 000 r/min,10 min),棄去沉淀,吸取上清液。在96孔板中加入190 μl勻漿上清液和10 μl探針,混勻后37 ℃孵育30 min后置于熒光酶標儀在激發波長488 nm、發射波長520 nm下進行檢測。細胞ROS檢測:根據試劑盒說明書進行操作,熒光探針DCFH-DA用無血清培養液進行稀釋(1 ∶1 000),收集細胞后加入稀釋好的DCFH-DA中制成懸液,于37 ℃培養箱中孵育20 min,用無血清培養液洗滌3次,洗去未進入細胞的熒光探針,在激發波長488 nm、發射波長525 nm條件下測定熒光強度。采用BCA蛋白定量法進行蛋白定量。
1.3.5腫瘤壞死因子(tumor necrosis factor-α,TNF-α )測定 取細胞培養的上清液,于4 ℃離心機(2 000 r/min、5 min)進行離心,收集上清液于-80 ℃保存待測。根據TNF-α 檢測試劑盒說明書進行實驗,測定樣品的吸光度值,根據所繪標準曲線計算出樣品的相應濃度。
1.3.6NADPH氧化酶活性檢測 準備組織及細胞,分別按組織與細胞NADPH氧化酶活性化學發光法定量檢測試劑盒說明書實驗步驟進行實驗,背景對照測定后檢測樣品總活性以及樣品非特異性活性,最后計算出樣品中NADPH氧化酶的實際活性。
1.3.7Real time PCR 根據總RNA提取試劑盒說明書進行操作,提取總RNA后,用逆轉錄試劑盒進行逆轉錄。GAPDH為內標,逆轉錄的cDNA為模板,采用SYBR green對各基因進行實時PCR。各基因相對表達量用2-ΔΔCt表示,每個樣品重復3次。
1.3.8Western blot法檢測 已處理細胞達80%后,PBS洗3次,加入100 μl含蛋白酶抑制劑的蛋白裂解液,冰上裂解30 min后收集細胞蛋白,4 ℃、14 000 r/min離心30 min,取上清液,用BCA法測定蛋白濃度,用蛋白裂解液配成同體積同濃度液體,再加入蛋白上樣緩沖液,混勻,然后煮沸使蛋白變性,-80 ℃保存。配制SDS-PAGE凝膠,等量上樣進行電泳,然后轉移到PVDF膜上。置搖床室溫封閉2 h(5%脫脂牛奶),一抗SIRT1(1 ∶10 000)4 ℃搖床孵育過夜,二抗兔抗(1 ∶5 000)室溫孵育2 h后進行顯影。用QuantityOne軟件分析各條帶的灰度值,以GAPDH為參照,計算相對灰度值。
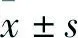
2 結果
2.1 AS中 miR-29c、TNF-α mRNA、ROS、NADPH氧化酶水平升高與正常組相比,miR-29c在AS小鼠主動脈中的表達明顯升高;TNF-α mRNA水平顯著升高,并且ROS水平和NADPH氧化酶活性也明顯提高,即炎癥和氧化應激水平明顯提高。見表1。

表1 主動脈miR-29c水平以及TNF-α mRNA水平、ROS水平和NADPH氧化酶活性
2.2 miR-29c可增加oxLDL誘導的HUVEC細胞中TNF-α表達、ROS生成與NADPH氧化酶活性與對照組比較,oxLDL組細胞內的TNF-α水平、ROS水平與NADPH氧化酶活性均明顯提高;轉染Mimic和Antagomir后細胞的TNF-α水平、ROS生成與NADPH氧化酶活性較對照組無明顯變化。此外,oxLDL刺激細胞后,Antagomir組細胞的TNF-α水平、ROS生成與NADPH氧化酶活性較對照組均明顯降低,而Mimic組細胞的TNF-α水平、ROS生成與NADPH氧化酶活性較對照組均明顯升高。見表2。

表2 各組細胞TNF-α含量、ROS水平與NADPH氧化酶活性
與對照組比較:*P<0.05;與oxLDL組比較:#P<0.05
2.3 oxLDL誘導下HUVEC 細胞的SIRT1活性和表達降低Real time PCR與Western blot法檢測結果顯示,與對照組相比,oxLDL組SIRT1 mRNA和蛋白表達均明顯降低,差異有統計學意義(t=4.260,P=0.013;t=4.905,P=0.008)。見圖1。
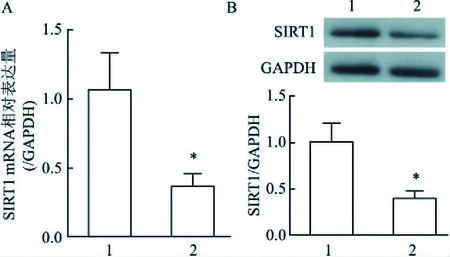
圖1 HUVEC中SIRT1 mRNA水平及蛋白表達A:細胞中SIRT1 mRNA水平;B:細胞中SIRT1蛋白表達;1:對照組;2:oxLDL組;與對照組比較:*P<0.05
2.4 miR-29c調控SIRT1表達Real time PCR與Western blot法檢測結果顯示,與對照組比較,Antagomir組和Mimic組細胞的SIRT1 mRNA水平并無明顯變化,差異無統計學意義(t=0.583,P=0.591;t<0.001,P=1.000),見圖2A、2B;然而,Antagomir組細胞的SIRT1蛋白表達顯著升高,而Mimic組細胞的SIRT1 蛋白表達則明顯降低,差異有統計學意義(t=12.422,P<0.001;t=4.691,P=0.009)。見圖2C、2D。
3 討論
miR-29c屬miR-29家族,在人類癌癥中大多下調。近年來有研究[6]表明,miR-29c可以通過靶向細胞外基質蛋白如膠原蛋白3A1和膠原蛋白4A5的mRNA來調節AS。此外,miR-29c可通過抑制胰島素樣生長因子1來抑制內皮細胞的遷移和血管生成[7]。最近,miR-29c被報道可作為鑒別AS患者的生物標志物[8]。以上研究表明,miR-29c在AS發展過程中發揮重要作用。
oxLDL可引起內皮細胞功能障礙,是AS發生的主要危險因素。本研究采用oxLDL 刺激HUVEC模擬高脂血癥損傷模型,研究miR-29c在AS發展中的潛在作用。數據顯示,與正常組相比,miR-29c在AS小鼠主動脈的表達明顯上調。此外,課題組對SIRT1下游的炎癥和氧化應激指標如TNF-α、ROS、NADPH氧化酶活性作了檢測。結果顯示,與正常組相比,AS小鼠主動脈的TNF-α mRNA水平、ROS生成與NADPH氧化酶活性明顯增高。在HUVEC培養中,經oxLDL刺激后,與對照組比較,轉染miR-29c antagomir的細胞中TNF-α的表達、ROS生成與NADPH氧化酶活性均明顯降低,而轉染miR-29c mimic的細胞中TNF-α的表達、ROS生成與NADPH氧化酶活性均明顯升高,差異均有統計學意義。由此可見,miR-29c水平與關鍵炎癥因子TNF-α水平、ROS、NADPH氧化酶活性顯著相關。
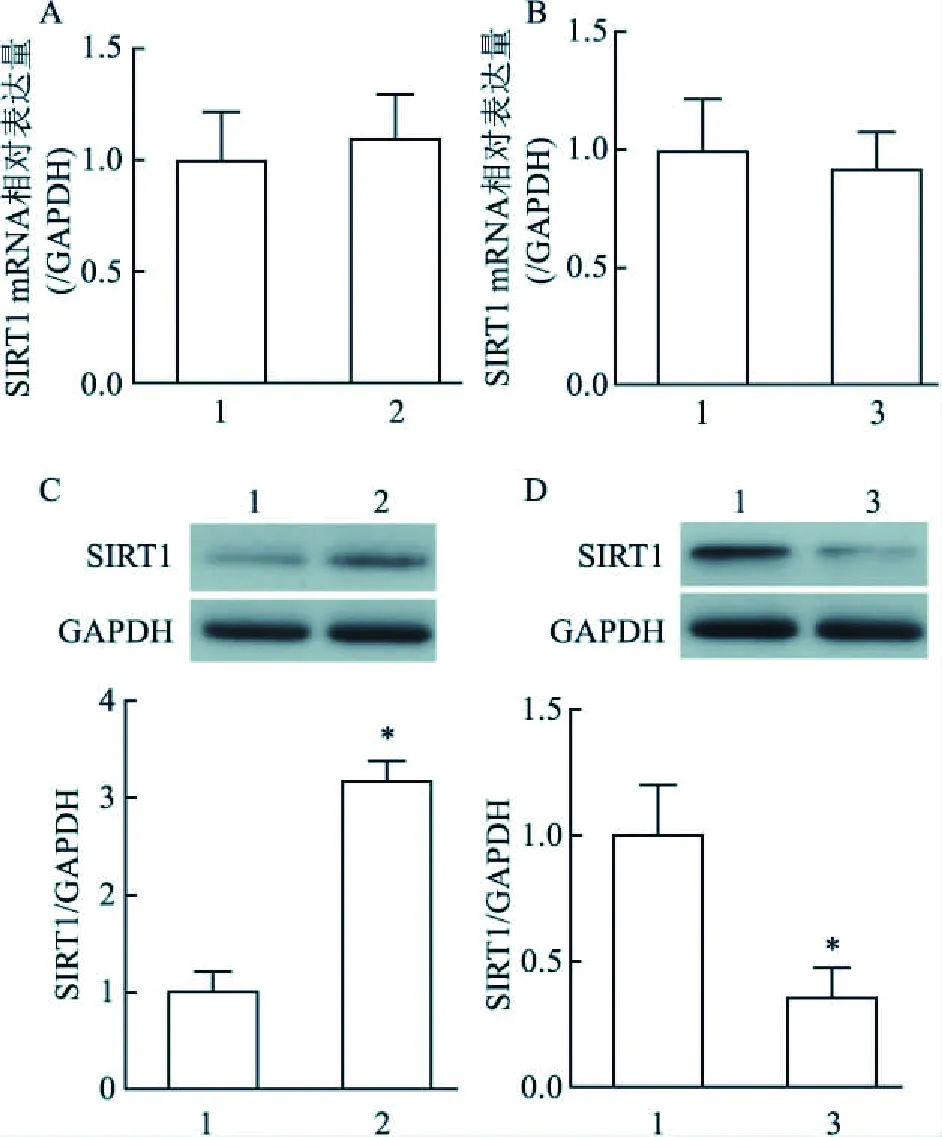
圖2 轉染miR-29c Mimic或Antagomir后細胞的SIRT1 mRNA水平以及蛋白表達A、B:細胞中SIRT1 mRNA水平;C、D:細胞中SIRT1蛋白表達;1:對照組;2:Antagomir組;3:Mimic組;與對照組比較:*P<0.05
SIRT1是sirtuin家族成員之一,在內皮細胞中高度表達,具有組蛋白脫乙酰酶的活性,可參與細胞的生長、凋亡等許多生物過程[9]。有研究[10]表明,SIRT1可抑制NADPH氧化酶活性,減少ROS產生,從而保護血管內皮功能。此外,SIRT1的特異性激活劑SIRT1720可通過抑制NF-κB信號傳導,減少血管緊張素Ⅱ誘導的多種促炎因子如TNF-α、單核細胞趨化蛋白-1(monocyte chemotactic protein 1,MCP-1)等的表達,減少炎癥細胞浸潤,從而發揮抗AS作用[11]。因此,SIRT1缺陷會增加炎癥和氧化應激,促進AS發生。有研究[12]報道,在肝癌中miR-29c通過直接靶向SIRT1抑制肝癌細胞生長從而起到腫瘤抑制劑的作用。此外,抑制miR-29可通過升高SIRT1的活性和表達來抑制脂肪的生成[13]。本研究顯示,oxLDL刺激后,與對照組相比,HUVEC的SIRT1 mRNA和蛋白表達水平均降低,并且在轉染miR-29c mimic細胞中SIRT1蛋白的表達明顯降低,轉染miR-29c antagomir細胞中SIRT1的表達顯著上升。然而,轉染miR-29c mimic和antagomir細胞中的SIRT1 mRNA水平較對照組差異無統計學意義,提示miR-29c對SIRT1的調控處于翻譯后水平。
綜上,miR-29c在AS發展過程中高表達,抑制miR-29c可上調SIRT1的表達,進而抑制NADPH氧化酶活性,減少ROS生成及炎性因子TNF-α表達,從而減弱AS氧化應激和炎癥反應。得出結論:miR-29c可下調SIRT1表達進而增強AS中氧化應激及炎癥反應。未來有必要進一步探討miR-29c調控SIRT1參與AS氧化應激和炎癥過程的具體作用機制,對防治AS有重要意義。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
學苑創造·A版(2020年9期)2020-10-13 09:41:02
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
人大建設(2019年12期)2019-05-21 02:55:32
西南軍醫(2016年6期)2016-01-23 02:21:19
西南軍醫(2015年2期)2015-01-22 09:09:37
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
食品科學(2013年15期)2013-03-11 18:25:48
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
