脂多糖致大鼠發熱特點與機制研究進展*
2020-04-19 07:12:16胡玉梅劉文君王振中
世界科學技術-中醫藥現代化 2020年7期
關鍵詞:劑量
楊 彪 ,胡玉梅 ,劉文君 ,姜 濤 ,耿 婷 ,曹 亮 ,王振中 ,肖 偉 **
(1. 江蘇康緣藥業股份有限公司 連云港 222001;2. 中藥制藥過程新技術國家重點實驗室 連云港 222001)
發熱是一種機體面對病原體入侵時產生的一種復雜的防御性反應,是生物歷經幾百萬年進化而來的一種生存策略[1-2]。機體每升高1℃需增強10%~12.5%的代謝速度[3],因此發熱通常伴隨一系列的“疾病行為”,如體溫升高、虛弱、疲倦等,而這些行為是為讓機體更快地恢復健康[4]。所以,發熱是一種生理反應,而不是病理反應。脂多糖(Lipopolysaccharide,LPS)是革蘭陰性菌外壁的類脂質多糖,其可通過與細胞表面Toll樣受體4(TLR-4)結合引發炎癥因子的大量表達,導致炎癥相關癥狀如紅、腫、熱、痛等,故常被用于誘導炎癥動物模型及細胞模型的建立[5]。自Szekely 以來,LPS 致大鼠發熱模型被廣范運用到發熱機制研究中,但由于多方面因素的影響,研究者得到的結果有所不同。因此,本文對LPS 引發大鼠發熱的特點與機制展開論述,為廣大實驗人員在建立大鼠發熱模型及解熱作用研究等方面提供一些幫助。
1 LPS致大鼠發熱的特點及影響因素
1.1 特點
LPS 致大鼠發熱的時間-體溫曲線會隨LPS 劑量、外界環境溫度的變化呈單相、雙相乃至多相變化,每一相也被稱為發熱的不同階段[6]。通常LPS 致發熱的產生可分為兩個階段。一是發熱的起始階段,也稱第一階段,此階段具有發熱迅速、持續時間較短等特點。二為發熱的持續階段,這個階段包括除發熱起始階段之外后續的發熱過程,通常包括第二及第三階段,具有持續時間長、多相變化等特點。此外,兩個階段有著截然不同的機制。
1.2 影響因素
1.2.1 LPS的劑量
LPS 劑量對大鼠發熱過程中的體溫調節有著重要作用[7]。在外界環境溫度適中的情況下,LPS 濃度在 100~101μg·kg-1時引起單相體溫升高,LPS 濃度在101.5μg·kg-1時,體溫呈現雙相波動,當 LPS 濃度達到101.75~104μg·kg-1時,發熱呈現出三相波動,且隨 LPS濃度的升高,發熱持續時間延長、幅度增大[8]。
在一定劑量范圍內LPS 致熱作用呈現劑量依賴性,但超出一定范圍后增加劑量并無作用[9]。劑量范圍跨度大:腹腔或靜脈注射10-1 000 μg/kg 不等,視模型需要做預實驗調整劑量[10]。通常注射劑量范圍20-250 μg/kg,引發雙相熱或三相熱,降低劑量呈單相熱,反復注射會致耐受[11]。內毒素緩釋,例:LPS 膠原水凝膠注射可克服反復注射LPS導致的耐受,形成SD大鼠從高熱到持續低熱[12]。常見造模劑量有三個:20 μg/kg、80 μg/kg、100 μg/kg,80 μg/kg劑量出現最多。20 μg/kg引發雙相熱,80 μg/kg、100 μg/kg引發三相熱。動物品系、生理狀態以及實驗室環境等均的會影響峰值出現時間、最大峰值,80 μg/kg 和 100 μg/kg LPS 致發熱比20 μg/kg致發熱模型發熱的熱勢高,發熱時間長,利于觀察以及進一步其他實驗操作,例如給藥觀察等。常見劑量20 μg/kg、80 μg/kg、100 μg/kg LPS 致大鼠發熱模型體溫變化趨勢見表1[13-23]。
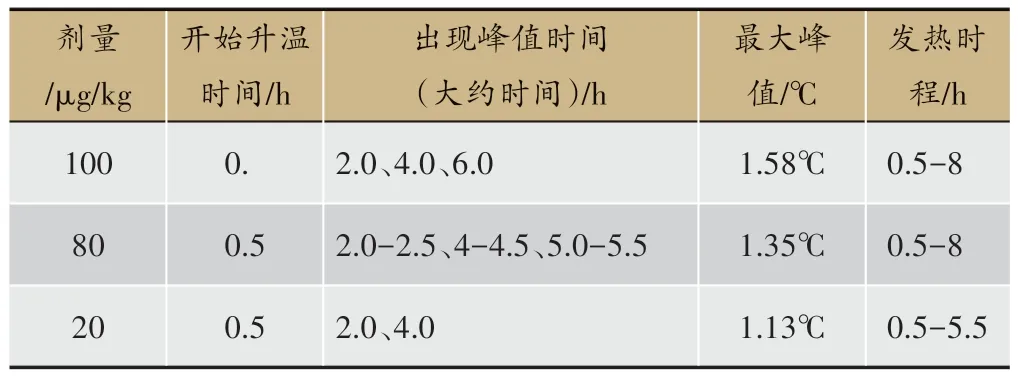
表1 常見劑量LPS致大鼠發熱模型體溫變化趨勢表
1.2.2 環境溫度
環境溫度對LPS誘導產生的發熱過程有著重要的影響,尤其是對體型較小的物種,如大鼠,很小溫度的差別均能產生重大影響,甚至可使體溫降低變成發熱。而且,不同溫度環境下,LPS致大鼠發熱過程有著明顯的區別。在29-31℃環境中,靜脈注射LPS 引起明顯的發熱,在低濃度時,LPS引起一個單相的體溫升高,當劑量升高后,LPS 引起幾個連續的體溫升高過程。而在一個溫度較低的環境中(26℃),同樣低濃度的LPS 引起雙相的發熱,隨著LPS 濃度的升高會導致低體溫現象出現[24]。
1.2.3 注射方法
通常實驗者采用腹腔或靜脈注射LPS 進行造模,而這種方法會讓大鼠產生嚴重的緊張和疼痛感,這種情況會造成大鼠的體溫波動1-3℃,并且持續30-120 min,對后續的體溫監測產生嚴重影響,并使正常的發熱起始階段被掩蓋或使單相發熱被認為是雙相發熱,具有嚴重的欺騙性。而且不同的給藥方法也會對發熱產生的時間有較大影響[25]。
1.2.4 其他
除上述三個主要因素之外,還有一些因素可能會造成影響,如大鼠的性別、品系、造模前禁食禁水與否、實驗者注射手法熟練程度等。其中,雌性大鼠的荷爾蒙水平會直接影響發熱反應。
2 發熱的不同階段及機制
2.1 發熱的起始階段
早期發熱理論認為發熱的起始階段是由內致熱源經血液循環通過血腦屏障作用于大腦發揮作用[26],后經實驗[27]證實,內致熱源如腫瘤壞死因子-α(TNF-α)等在LPS 注射后的30-60 min 才出現在血液循環中,而此時發熱已產生。隨后補體系統的發現以及迷走神經傳輸理論成為發熱起始階段的關鍵因素,并受到廣范認可。通常發熱的起始階段發生在LPS注射后15 min,并在1 h 左右達到峰值[28]。起始階段的潛伏期會隨著LPS劑量的增加而有所縮短,幅度也會有所增加。
2.1.1 起始階段的關鍵因素
2.1.1.1 Kupffer細胞(Kc)
Kc 是機體內最大的巨噬細胞群,存在于肝臟中,擔任著LPS 代謝清除的任務。同時,它也是TLR-4 的主要供體細胞以及補體因子5a(C5a)的作用細胞,在發熱的啟動過程和終止過程中起到重要的作用。L 等[29]采用異硫氰酸熒光素(FITC)標記LPS,觀察到發熱起始階段的體溫升高與Kc 攝取FITC 標記的LPS 密切相關。
2.1.1.2 補體系統的激活
補體系統是自身免疫系統的一個重要組成部分,它可被病原體及其代謝產物迅速激活[30]。靜脈注射LPS 可在2 min內引起補體的級聯反應,引起各種細胞因子的生成,且大多數炎癥因子白細胞介素-1(IL-1)、IL-6、前列腺素 E2(PGE2)和 TNF-α 等與發熱密切相關[31]。補體系統的激活過程主要發生在Kc 細胞中,如圖1 所示。補體C 代謝生成補體C3,補體C3進一步代謝生成C5a,C5a通過與補體因子5a受體(C5aR)結合啟動磷脂酶C(PLC)水解生成花生四烯酸(AA),接著AA在環氧酶-1/2(COX-1/2)的作用下生成PGE2和前列腺素D2(PGD2)等[32]。Li等[33]通過基因敲除及C5aR受體拮抗劑證實了C5a在發熱的起始過程中起到重要作用。
2.1.1.3 肝臟傳入迷走神經與去甲腎上腺素(NE)
肝臟傳入迷走神經元表面表達有前列腺素EP3受體[34],可與C5a誘導產生的PGE2結合,通過迷走神經將信號向下丘腦前區(POA)傳送。Simons 等[35]研究發現肝臟迷走神經切斷術可阻斷發熱產生及PGE2升高,證實了肝臟迷走神經在發熱起始過程中有著重要的作用。Kc-PGE2信號傳入POA 后會刺激NE 的釋放,引起棕色脂肪代謝及皮膚收縮,導致發熱。同時生成的NE 會刺激PGE2的生成,值得注意的是此處生成的PGE2是由神經膠質、神經元本身及樹突小棘產生[36-37]。

圖1 發熱起始階段信號傳導通路圖
2.1.1.4 激活內源性Kalikrein-kinin系統
研究[38]發現 LPS 激活內源性 Kalikrein-kinin 系統導致Bradykinin(BK)產生,其作用于POA 細胞的B2 受體導致前列腺素合成,進而產生發熱。
2.1.1.5 信號傳導過程
如圖1 所示,LPS 進入機體后隨血液循環至肝臟(Liver)中,通過激活補體系統產生PGE2,產生的PGE2通過與肝臟傳入迷走神經元(Hepatic vagus neuron)的EP3受體結合將信號傳入到非顫栗產熱(NST)細胞群A1和A2后,再通過腹側的NE能神經束進一步傳遞到POA,引起NE的釋放,NE通過與溫敏神經元上的腎上腺素受體α1(α1-AR)結合,引起棕色脂肪代謝及皮膚收縮,產生快速發熱。
2.2 發熱的持續階段
關于發熱持續階段的機制有過不同的理論,目前最被學者認可的是血腦屏障(BBB)在發熱的持續中產生重要的作用[39]。研究[40]發現BBB 周圍的星形膠質細胞、內皮細胞、巨噬細胞等均含有TLR-4 受體,而且大腦內皮細胞表達有TNF-α、IL-1、IL-6 受體。這些均表明BBB 不是信號傳導的障礙而是信號的產生者,在發熱的持續階段具有重要的地位。
2.2.1 持續階段涉及的關鍵因素
2.2.1.1 LPS的識別—TLR-4的激活
大多數病原體有著高度相似的組成結構,被稱為“病原體相關分子模式”,而這些結構會通過TLRs 家族激活機體的自身免疫系統。其中,TLR-4 是LPS 的同源受體,常在表達于巨噬細胞、中性粒細胞及樹突狀細胞表面。LPS與LPS連接血清蛋白(LPB)結合后,在CD14分子作用下與由MD2 分子作用激活的TLR-4結合,過程中還涉及到MyD88、IRAK和TRAP6等因子,最終激活核轉錄因子-κB(NF-κB),生成多種炎癥因子(TNF-α、IL-1、IL-6等)、黏附因子以及一些炎癥相關的酶[一氧化氮合成酶(iNOS)、環氧酶-2(COX-2)、前列腺素合成酶(mPGES-1)等][41-43]。研究[44-45]采用基因敲除技術發現TLR-4基因敲除C3H/HeJ大鼠不能對LPS刺激產生發熱反應,證實了TLR-4 在發熱中的重要作用。
2.2.1.2 內致熱源的產生
內致熱源最初由Beeson[46]提出,主要是指由外致熱源,如LPS,誘導下由機體自身產生的,促使發熱的因子,多是炎癥因子。在發熱過程中,無論是LPS 與TLR-4 結合激活NF-κB,還是補體系統,都可導致炎癥因子(TNF-α、IL-1、IL-6 等)的產生。研究[47-49]發現,靜脈注射 LPS 后30-60 min,TNF-α,IL-1,IL-6 相繼在血液中出現。而且,TNF-α與IL-1可相互促進彼此的生成,兩者都可促進IL-6 生成,而IL-6 能抑制TNF-α 與 IL-1 生成。TNF-α、IL-1、IL-6 主要在發熱的維持階段起到作用,與發熱的起始階段無關。其中,IL-6對機體免疫功能有促進作用。
2.2.1.3 PGE2的生成與傳導
PGE2是公認的發熱介質之一[43]。PGE2與下丘腦部位的EP3受體結合是維持發熱的重要過程。其代謝過程主要由細胞膜磷脂在磷脂酶A2 作用下水解生成的 AA 經 COX 及 mPGES 代謝生成。在 LPS 誘導的發熱過程中,COX-2 和mPGES-1 不僅在大腦部位的巨噬細胞中大量表達,而在肝臟和肺部的巨噬細胞中也大量表達[50]。采用抑制前列腺素(PG)合成的藥物以及COX-2或mPGES-1缺陷小鼠均可抑制發熱產生[51]。同時,在發熱的持續階段,肝臟部位的PGE2代謝酶表達量降低,促進了PGE2向大腦的轉移[52]。
2.2.1.4 EP3受體
EP3受體在下丘腦視前區大量表達[53]。PGE2受體與EP3在腦部結合是發熱產生的必要條件之一[54]。Michael 等[55]研究表明PGE2與處于內側視前核的神經元表達的EP3受體結合是發熱產生的關鍵因素。
2.3 信號傳導過程
發熱持續階段的PGE2可分為兩部分,一部分由外周生成轉移至大腦,另一部分由大腦部位自身生成。兩部分的產熱機制基本相同,都是通過LPS 直接或間接作用而產生。過程如圖2 所示。血液循環中的LPS通過與外周巨噬細胞、中性粒細胞等表面的TLR-4 受體結合并激活NF-κB 產生各種炎癥因子TNF-α、IL-1、IL-6 等,生成的炎癥因子可通過 PLA2-AA-PGE2產生PGE2。同時,炎癥因子及LPS 可直接與大腦內皮細胞表面受體(TLR-4R、IL-1R、IL-6R)結合,產生PGE2,機制與外周相同。外周生成的PGE2通過BBB進入中樞,同BBB 處生成的PGE2共同與神經元的EP3受體結合,生成NE,繼而引起棕色脂肪代謝增加,血管收縮等反應,引起發熱。
3 小結
LPS致大鼠發熱模型現已廣泛應用于各種解熱實驗中,充分了解LPS 致大鼠發熱的特點與機制對實驗者有著重要的幫助。本文對LPS致大鼠發熱過程中的影響因素進行了總結,主要包括LPS的劑量、環境溫度以及注射的方法,把控好這些因素會對模型的建立以及實驗的可靠性提供一定的幫助。在模型建立的過程中,可根據實驗目的給予 LPS 劑量在 101.75~104μg·kg-1范圍內維持發熱的穩定性。可通過控制通風、空調等條件嚴格控制實驗環境的溫度在25℃左右,一定不能高于體溫。同時,在給藥及測量體溫的過程中,實驗操作者對大鼠的抓取要經過練習,最好是使大鼠習慣操作者的抓取動作,避免因抓取使大鼠產生掙扎導致體溫的變化。最后,條件較好的情況下可采用皮下植入體溫測定裝置,以計算機實時監控大鼠的體溫變化,這里需要注意,手術植入體溫測定后需要對大鼠進行至少一周的消炎處理,避免其發生炎癥反應。
在了解發熱特點的基礎上,本文對LPS 致大鼠發熱的機制進行了初步總結。發熱可分為兩個階段,一是起始階段,二是持續階段。兩個階段有著不同的特點并有著不同的機制。在發熱的起始階段,LPS 通過肝臟Kc 中的補體系統快速生成PGE2,PGE2激活肝臟迷走神經傳入神經元、NST,最終將信號傳入POA,生成NE,引起發熱。在發熱的持續階段中,LPS 通過與TLR-4 受體結合激活NF-κB 產生各種炎癥因子TNF-α、IL-1、IL-6 等,這些炎癥因子以及 LPS 通過與大腦部位的細胞,包括內皮細胞、星形膠質細胞等部位的受體結合,激活細胞內COX-2 生成PGE2,PGE2通過與神經元EP3 受體結合,傳導信號至POA,引發NE 釋放,進而導致發熱。雖然研究將發熱分為兩個階段,并在時間上存在先后關系,但兩個階段并非相互獨立,兩者也是同時進行并相互影響的。
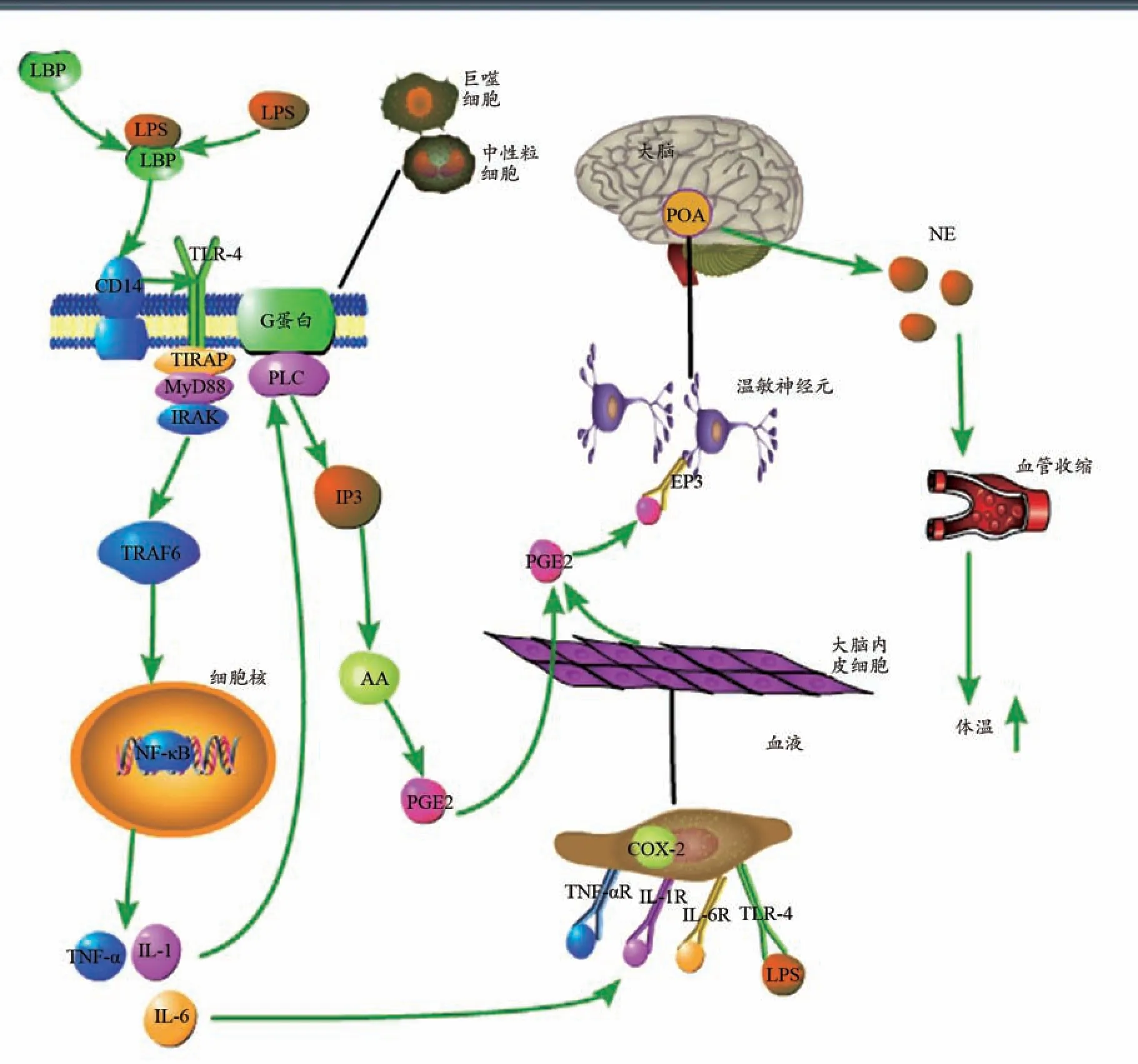
圖2 發熱持續階段信號傳導通路圖
目前對發熱的機制研究已較為明確,但仍然存在一些疑問,比如低溫環境中高劑量LPS 導致低體溫現象的機制是什么?替代激活的脂肪中巨噬細胞在LPS致發熱中有什么作用?這些問題仍需科研人員進一步研究解答。本文對LPS致大鼠發熱機制進行了一定的總結,希望對科研人員在建立LPS 致大鼠發熱模型中所遇到的問題有所幫助。
猜你喜歡
課堂內外·初中版(科學少年)(2023年10期)2023-12-10 00:43:06
全科護理(2022年10期)2022-12-26 21:19:15
中國合理用藥探索(2022年1期)2022-11-26 00:22:32
今日農業(2022年4期)2022-11-16 19:42:02
鄉村科技(2021年33期)2021-03-16 02:26:54
國際放射醫學核醫學雜志(2021年10期)2021-02-28 08:41:58
藥學與臨床研究(2015年4期)2015-06-05 11:35:54
衛生職業教育(2014年24期)2014-05-20 09:05:38
同位素(2014年2期)2014-04-16 04:57:20
中國合理用藥探索(2014年11期)2014-03-11 20:30:20
