多倍體銀鯽F系bmp15不同等位基因的分子特征、基因組結構和表達模式
2020-06-12 12:51:26江詩雨桂建芳
水生生物學報 2020年3期
江詩雨 汪 洋 周 莉 陳 帆 李 志 桂建芳
(1. 中國科學院水生生物研究所, 淡水生態與生物技術國家重點實驗室, 武漢 430072; 2. 中國科學院水生生物研究所種子創新研究院, 北京 100101; 3. 中國科學院大學, 北京 100049)
骨形態發生蛋白信號通路(Bone morphogenetic protein signaling, BMP signaling)在動物生殖系統的發育和功能維持的多個生命過程中發揮著重要的調控作用, 包括原始生殖細胞特化、增殖和遷移, 配子發生、濾泡形成等[1]。其中, BMP15主要由卵母細胞分泌, 首先在人和小鼠中被克隆[2], 后被證實為脊椎動物所特有[3], 在哺乳動物的濾泡發育和閉鎖、產卵、繁殖力維持以及授精等過程中起著關鍵作用[4—10]。
關于魚類bmp15基因研究得較少, 近年來僅在斑馬魚(Danio rerio)[11]、歐洲鱸魚(Dicentrarchus labrax)[12,13]、銀鯽(Carassius gibelio)[14]、稀有鯽(Gobiocypris rarus)[15]、黃尾(Seriola lalandi)[16]、黑鯛(Acanthopagrus schlegelii)[17]、大西洋鮭(Salmo salarL.)[18]和胡鲇(Clarias batrachus)[19]中報道了bmp15的克隆和表達分析。對這些魚類bmp15基因的表達分析表明,bmp15基因不僅在成體組織的分布, 而且在不同發育階段性腺和胚胎中的表達譜式以及在卵巢中的定位都存在差異, 表現出物種的特異性。譬如, 在斑馬魚、歐洲鱸魚、銀鯽和稀有鯽中, 除了在卵巢中高表達和精巢中中等豐度的表達外, 在肝、腎、腦、垂體等多個組織中也有低豐度的表達[11,12,14,15]; 而大西洋鮭bmp15-like僅在性腺中表達, 在其他組織中檢測不到[18]。不同于哺乳動物和其他魚類bmp15的報道[2,12,14,15,20—22], 斑馬魚bmp15不僅在卵子中表達, 還在顆粒細胞和鞘細胞中檢測到了mRNA和蛋白的存在[11]。
多倍化(Polyploidization)是指基因組中加入了一套或多套完整的染色體組。除了在脊椎動物進化歷程中均發生的兩輪全基因組復制外, 輻鰭魚類還發生了第3輪魚類特異的全基因組復制(Fish-specific genome duplication, Ts3R)[23—25]。在突破不穩定的瓶頸后, 多倍體會通過隨后的二倍化進化成一個新的二倍體。在“多倍化-二倍化”的進化歷程中,多倍體通過重復基因的新功能化或亞功能化、基因組的重排、基因的表達變異、等位基因的特異沉默、表觀遺傳的重塑等創造了大量的進化機會,增加了等位基因的多樣性, 改變了基因組的復雜性,產生了新的性狀并驅動改變生態位和擴大地理分布[26—31]。多倍體銀鯽通常被視為鯽(Carassius auratusL.)的一個亞種(C. auratus gibelioBloch)[32], 但隨著對其核型、生殖方式、性別決定方式等研究的深入, 已逐漸被視為一個獨立的物種[33—40]。銀鯽擁有156條染色體或162條染色體[41], 由于其具有100條染色體的近緣種金魚已被證明發生了四倍化事件[42], 因此銀鯽應為超三倍體或進化的六倍體[43]。通過對銀鯽2個歧化的Dmrt1基因的序列和染色體定位分析揭示了銀鯽進化歷程中發生了兩輪多倍化事件[44]。盡管已在銀鯽中鑒定了一個bmp15基因[14],但對銀鯽Dmrt1基因[44]、頭腎組織的轉錄組[45]以及干擾素系統基因的分析[46]均表明, 銀鯽是一個異源多倍體, 一個基因在其基因組中至少存在2個高度分化的等位基因。除了已鑒定的bmp15基因外, 銀鯽是否還存在其他bmp15等位基因, 它們在表達上是否發生了明顯歧化, 目前尚不清楚。因此, 本研究克隆鑒定了2個歧化的bmp15基因Cgbmp15a和Cgbmp15b的6個等位基因, 分析了它們的基因組結構和與其鄰近基因的共線性關系, 以及Cgbmp15a和Cgbmp15b在成體組織、不同發育階段卵子和孕酮激素DHP誘導卵母細胞成熟過程中的表達差異,為研究多倍體魚類重復基因的進化和bmp15在魚類生殖調控中的作用提供基礎數據。
1 材料與方法
1.1 實驗材料
本研究所用的銀鯽F系選自于中國科學院水生生物研究所官橋實驗基地。繁殖季節隨機抽取3條性成熟的銀鯽F系, 取肝、脾、腎、腦、下丘腦、垂體和卵巢組織部分樣品; 并將卵巢置于盛有生理鹽水的培養皿中, 在顯微鏡下利用機械分離法獲得不同發育時期的卵細胞, 根據直徑大小進行分類, 分為初級卵母細胞(0.06—0.13 mm, Ⅰ期)、皮質泡期卵母細胞(0.21—0.23 mm, Ⅱ期)、卵黃形成期卵母細胞(0.30—0.33 mm,Ⅲ1期和0.45—0.48 mm, Ⅲ2期)、成熟卵母細胞(1.04—1.22 mm, Ⅳ期)。將各樣品放入無RNA酶的EP管中, 迅速經液氮速凍后置于-80℃備用。
1.2 總RNA提取和SMARTer cDNA模板合成
所用離心管、槍頭、溶液均無RNA酶, 玻璃器皿及金屬器械經240℃高溫烘烤4h后使用。按照Trizol和SV Total RNA Isolation System試劑盒(Promega)說明書進行各樣品總RNA的提取, 并分別用Nanodrop 2000C(Thermo Fisher Scientific, 美國)分光光度計和1.2%(w/v)瓊脂糖凝膠電泳檢測RNA的濃度和完整性。用SMARTer RACE 5′/3′ Kit(Clontech,美國)合成了銀鯽F系卵巢組織3′ cDNA和5′ cDNA,具體參考Xie等[47]描述的方法。合成的cDNA置于-20℃保存。
1.3 銀鯽F系Cgbmp15a和Cgbmp15b cDNA全長克隆
以斑馬魚bmp15的基因組序列為搜索序列, 在銀鯽F系基因組中搜尋到了2個同源性較高的序列ENSDARP00000054590-C1和ENSDARP 00000054590-C2, 分別命名為Cgbmp15a和Cgbmp15b。參照這2個序列, 利用Primer Premier 5和Oligo7.0軟件設計RACE內外引物(表1)。以卵巢3′ cDNA和5′ cDNA為模板, 使用RACE外引物Cgbmp15a-5′-Outer、Cgbmp15b-5′-Outer、Cgbmp15a-3′-Outer、Cgbmp15b-3′-Outer與UPM分別進行5′或3′第一次PCR, 電泳檢測首次PCR產物若無目的條帶, 將首次PCR產物稀釋100倍后為模板, 用相應的RACE內引物Cgbmp15a-5′-Inner、Cgbmp15b-5′-Inner、Cgbmp15a-3′-Inner、Cgbmp15b-3′-Inner和NUP進行巢式PCR, 2次PCR的反應體系和反應程序均參見文獻[48]。目的條帶經1.1%瓊脂糖凝膠電泳采用Gel Extraction Kit(Omega Bio-Tek, 美國)回收后, 產物連接pMD18-T載體(TaKaRa), 各選取20個陽性克隆由鉑尚生物技術(上海)有限公司進行雙向測序, 序列經拼接后, 得到Cgbmp15a和Cgbmp15b的cDNA全長序列。
1.4 銀鯽F系Cgbmp15a和Cgbmp15b基因序列分析與進化分析
根據已獲得的Cgbmp15a和Cgbmp15b全長cDNA序列, 應用DNAMAN預測開放閱讀框(ORF),推導其編碼的氨基酸序列, 并在Ensembl genome browser 92(http://asia.ensembl.org/index.html)和NCBI(https://www.ncbi.nlm.nih.gov/)上下載多個物種的Bmp15氨基酸序列, 應用BioEdit軟件進行Bmp15多重序列比對分析。不同物種Bmp15氨基酸的登錄號如下: 銀鯽Carassius gibelioBmp15(ADW20149.1)、鯽Carassius auratusBmp15a-1(MH297584)、鯽Carassius auratusBmp15a-2(MH297583)、鯽Carassius auratusBmp15b(MH 297582)、斑馬魚Danio rerioBmp15(AAI24107.1)、稀有鯽Gobiocypris rarusBmp15 (AHK22790.1)、河鲀Takifugu rubripesBmp15(ENSTRUP000000 17355)、羅非魚Oreochromis niloticusBmp15(XP_003457772.1)、青鳉Oryzias latipesBmp15(XP_004079992.1)、雀鱔Lepisosteus oculatusBmp15(XP_006632858.2)、爪蟾Xenopus tropicalisBmp15(XP_002936834.2)、雞Gallus gallusBMP15(ENSGALP00000007547)、小鼠Mus musculusBMP15(AAH55363.1)和人Homo sapiensBMP15(EAW 89914.1)。

表1 本文中所用引物Tab. 1 Primers used in this study
以Cgbmp15a和Cgbmp15bcDNA序列為基礎,在其ORF區域設計引物來進行銀鯽F系BAC文庫篩選。篩選到的BAC序列目的片段經測序驗證后, 利用同源法注釋對BAC上的基因進行預測, 進行bmp15的基因組結構和與其鄰近基因的同線性關系分析。用MEGA 6.06軟件, 采用最大似然法(ML法)對銀鯽bmp15、鯽bmp15和斑馬魚bmp15的基因組序列進行系統進化樹構建。
1.5 Cgbmp15a和 Cgbmp15b在銀鯽F系不同組織以及不同時期卵母細胞中的表達
選取樣品總RNA 0.1 μg, 利用GoldScript cDNA合成試劑盒(invitrongen, 上海)反轉錄得到cDNA,具體步驟參見文獻[49]。使用Primer Premier 5.0設計Cgbmp15a和Cgbmp15b基因RT-qPCR特異性引物(表1)。選取異育銀鯽β-actin作為內參基因, 用CFX96TMOptics Module(美國Bio-Rad公司)進行熒光定量PCR, 反應條件如下: 95℃預變性30s; 95℃變性5s, 60℃, 30s, 40cycles; 65℃, 5s; 95℃, 5s。進行3次生物學重復, 并每個樣品做3次技術性重復,以β-actin作為內參引物, 序列參見文獻[48], 用2-ΔΔCt方法計算基因的相對表達量, 采用平均值±標準誤(Mean±SEM)來表示。數據分別經過正態性檢驗和方差齊性檢驗后, 采用SPSS 21.0軟件進行方差分析,P<0.05表示存在顯著性差異, 柱形圖用Graph-Pad Prism 6進行繪制。
1.6 Cgbmp15a和 Cgbmp15b在DHP體外誘導銀鯽F系卵母細胞成熟過程中的表達分析
在繁殖季節取性成熟的銀鯽F系雌性個體, 用鑷子將卵母細胞從卵巢中剝離出, 挑取處于生發泡1期(Germinal vesicle 1 stage, GV1期)卵母細胞, 轉移至預熱(21—23℃)的卵母細胞培養液格氏平衡鹽溶液(Gey’s balanced salt solution, GBSS, 7.25 g NaCl, 0.41 g NaH2PO4, 0.38 g KCl, 0.95 g HEPES,1.0 g葡萄糖, pH8.5, 0.147 g CaCl2, 0.23 g MgSO4,0.1 g 1000 U青霉素, 0.1 g硫酸鏈霉素, 配制1 L)中培養。再向培養液中加入孕酮激素(17α, 20β-dihydroxy-4-pregnen-3-one, DHP, 終濃度為1 μg/mL),每毫升培養液培養10顆卵母細胞, 23℃培養誘導卵母細胞體外成熟[50]。每隔2h取樣1次, 每次取30顆卵, 利用Trizol和SV Total RNA Isolation System試劑盒說明書進行各樣品總RNA的提取、反轉錄cDNA以及qRT-PCR檢測Cgbmp15a和Cgbmp15b基因的表達。擴增條件和數據分析同上。
2 結果
2.1 銀鯽F系Cgbmp15a和Cgbmp15b的3個不同等 位基因的分子特征
根據銀鯽F系基因組中2個與斑馬魚bmp15基因的同源序列設計特異引物, 通過RACE-PCR各獲得了Cgbmp15a和Cgbmp15b的3個全長cDNA序列。其中Cgbmp15a的3個全長cDNA大小分別為2133、2147和2143 bp, 它們之間的平均一致性為(98.73±0.35)%; 它們的開放閱讀框(ORF)均為1128 bp,編碼375個氨基酸, 平均一致性為(99.29±0.13)%, 它們在第152、第157、第164和第286位氨基酸存在差異(圖1)。而Cgbmp15b的3個全長cDNA存在較大的差異, 大小分別為2030、1824和1648 bp, 它們之間的一致性為(84.88±4.40)%, 差異主要在3′端;它們的開放閱讀框均為1143 bp, 編碼380個氨基酸,平均一致性為(99.3±0.33)%, 在第167、第203、第268和第359位氨基酸存在差異(圖1)。Cgbmp15a和Cgbmp15b之間cDNA的一致性為(73.85±6.25)%,氨基酸的一致性為(87.36±0.17)%。
不同物種Bmp15蛋白序列比對結果顯示, 本研究克隆的CgBmp15a和CgBmp15b與已報道的銀鯽Bmp15[14]的一致性分別為(92.13±0.12)%和(94.20±0.12)%, 表明已報道的銀鯽Bmp15跟CgBmp15b同源性更高。同時我們在四倍體鯽基因組中搜索到3個bmp15等位基因, 分別命名為Cabmp15a-1、Cabmp15a-2和Cabmp15b; 在斑馬魚、稀有鯽、河鲀、羅非魚、青鳉和雀鱔等其他魚類基因組中,均只搜索到一個bmp15基因。CgBmp15a和CgB-mp15b與其他魚類Bmp15的平均一致性分別在35.50%—80.52%和36.55%—81.82%, 而與人、小鼠、雞和爪蟾Bmp15的一致性在28.27%—41.52%和29.30%—32.48%。

圖1 銀鯽Cgbmp15a和Cgbmp15b的3個等位基因的基因組結構Fig. 1 Schematic diagram for gene structure of gibel carp Cgbmp15as and Cgbmp15bs
2.2 銀鯽F系Cgbmp15a和Cgbmp15b的不同等位基因的基因組結構和相鄰基因共線性分析
我們將所克隆到的6個銀鯽bmp15等位基因序列在銀鯽F系的BAC文庫中進行篩選, 并將篩選到的BAC克隆的全序列與6個cDNA序列進行比對, 分別獲得6個含銀鯽bmp15 cDNA序列的BAC克隆。Cgbmp15a的3個BAC克隆的大小分別為122.75、145.62和106.83 kb,Cgbmp15b的3個BAC克隆的大小分別為90.06、151.96和35.05 kb。我們對6個BAC克隆上的基因進行了預測, 并分析了銀鯽6個bmp15等位基因的基因組結構。研究結果表明,Cgbmp15a和Cgbmp15b的3個不同等位基因具有相同的基因組結構, 均由兩個外顯子和一個內含子組成(圖1)。Cgbmp15a的3個等位基因的兩個外顯子長度均分別為315和810 bp, 內含子大小為398 bp;Cgbmp15b的3個等位基因的2個外顯子長度均分別為333和807 bp, 內含子為416 bp; 通過SMART(http://smart.embl-heidelberg.de/)軟件分析表明這6個Bmp15蛋白都含有一個保守的TGF-β結構域, 且N端都含有一個信號肽。其中CgBmp15a的信號肽處于N端1—32位, TGF-β結構域位于第274—375位氨基酸; 而CgBmp15b的信號肽處于N端1—28位,TGF-β結構域位于第279—380位氨基酸。
接著, 我們比較了6個BAC克隆上與其他鄰近基因的同線性關系(圖2)。Cgbmp15a-1-BAC、Cgbmp15a-2-BAC和Cgbmp15a-3-BAC均含有NHS-like2(nhsl2)、ribosomal protein S4、X-linked(rps4x)、histone deacetylase8(hdac8)、liver-expressed antimicrobial peptide2(leap2)、bmp15、glutamic-oxaloacetic transaminase2b(got2b)、solute carrier family38、member7(slc38a7)基因, 且具有保守的共線性關系。由于Cgbmp15a-2-BAC比Cgbmp15a-1-BAC和Cgbmp15a-3-BAC分別長22.87和38.79 kb,因此還包含lysosomal protective protein-like(lppl)、cytochrome P450,family2、subfamily X、polypeptide10.2(cyp2x10.2)、3A1-like、pollen-specific leucine-rich repeat extensin-like protein2 (pslr relp2)基因。而Cgbmp15b的3個BAC克隆序列長度和包含的序列差異較大, 除了bmp15基因外, 還均含有got2b基因。與Cgbmp15b-1-BAC和Cgbmp15b-3-BAC相比,Cgbmp15b-2-BAC缺失了slc38a7基因。與Cgbmp15a的3個BAC序列相比,Cgbmp15b-2-BAC的bmp15上游還存在Tc1-like基因, 但缺失了nhsl2、leap2和slc38a7基因; 而Cgbmp15b-1-BAC的bmp15下游還包含3A1-like基因, 但缺失了lppl和cyp2x10.2基因。同時, 我們在Ensembl數據庫獲得了人、小鼠、雞、斑馬魚、青鳉和河鲀的bmp15基因所在區域的基因組序列, 與銀鯽bmp15基因所在區域進行同線性分析。結果表明, 斑馬魚bmp15基因及其8個鄰近基因與銀鯽Cgbmp15a和鄰近基因的同線性關系保守性最強, 僅在bmp15下游缺失了lppl和pslrrelp2基因。而其他脊椎動物bmp15基因下游未預測到與銀鯽Cgbmp15a的相同下游基因。與銀鯽Cgbmp15a-BAC、斑馬魚和青鳉相比, 人、小鼠、雞和河鲀bmp15基因上游還缺失了leap2基因。
為了揭示銀鯽6個bmp15等位基因的進化關系和起源, 我們以斑馬魚bmp15、3個鯽bmp15等位基因、6個銀鯽bmp15等位基因的基因組全序列, 采用最大似然法(ML法)構建了系統進化樹(圖3)。從圖中我們可以看出, 斑馬魚Drbmp15作為外群, 銀鯽Cgbmp15a的3個等位基因與鯽的Cabmp15a的2個等位基因聚為一簇, 銀鯽Cgbmp15b的3個等位基因與鯽的Cabmp15b聚為另一簇, 而不是同一物種bmp15的不同等位基因聚在一起。由此我們推測銀鯽和鯽擁有共同的原始祖先, 并且在圖3中標注出了在祖先家系中基因/基因組復制事件。

圖2 銀鯽Cgbmp15a和Cgbmp15b基因與其鄰近基因保守的共線性關系(數據來自Ensemble數據庫)Fig. 2 Schematic diagram for bmp15 and context genes of gibel carp Cgbmp15a and Cgbmp15b BACs and the bmp15 homologs of other species from Ensemble database
2.3 銀鯽F系Cgbmp15a和Cgbmp15b在卵巢中高豐度表達
由于Cgbmp15a和Cgbmp15b的3個不同等位基因之間的一致性非常高, 因此我們針對Cgbmp15a和Cgbmp15b之間的序列差異, 設計2對特異引物,采用RT-qPCR方法檢測Cgbmp15a和Cgbmp15b在銀鯽F系成體組織中的分布。結果表明, 銀鯽F系Cgbmp15a和Cgbmp15b均在卵巢中高表達, 并且Cgbmp15a的表達水平顯著高于Cgbmp15b(P<0.05)。Cgbmp15b在垂體中可檢測到微量轉錄本的存在,而在肝、脾、腎、腦和下丘腦中檢測不到轉錄本的存在。與Cgbmp15b不同的是, 除了在垂體中外,Cgbmp15a還可以在肝、腦和下丘腦中檢測到低豐度轉錄本的存在(圖4)。
2.4 銀鯽F系Cgbmp15a和Cgbmp15b隨著DHP體外誘導卵母細胞成熟上調表達水平
由于Cgbmp15a和Cgbmp15b在銀鯽卵巢中高表達, 隨后我們分析了它們在不同發育階段卵母細胞中的表達水平差異。結果表明, 隨著卵母細胞的發育和成熟,Cgbmp15a和Cgbmp15b表達水平均呈先升高再降低的趨勢; 在皮質泡期卵母細胞中表達量達到最高, 隨后降低; 且在初級卵母細胞到卵黃形成期卵母細胞中,Cgbmp15a的表達量始終顯著高于同時期Cgbmp15b的表達量(P<0.05); 在成熟卵母細胞中二者無顯著性差異(圖5A)。
接著, 我們采用DHP體外誘導銀鯽F系GV1期卵母細胞成熟, 并分析了Cgbmp15a和Cgbmp15b在誘導成熟過程中的動態變化。結果表明,Cgbmp15a在誘導6h后開始上調表達, 在誘導8h后達到最高表達水平, 隨后下降。而Cgbmp15b在誘導8h后表達水平有微弱上調, 隨后下降。在誘導后6—12h,Cgbmp15a表達量均顯著高于同時期Cgbmp15b的表達量(P<0.05, 圖5B)。
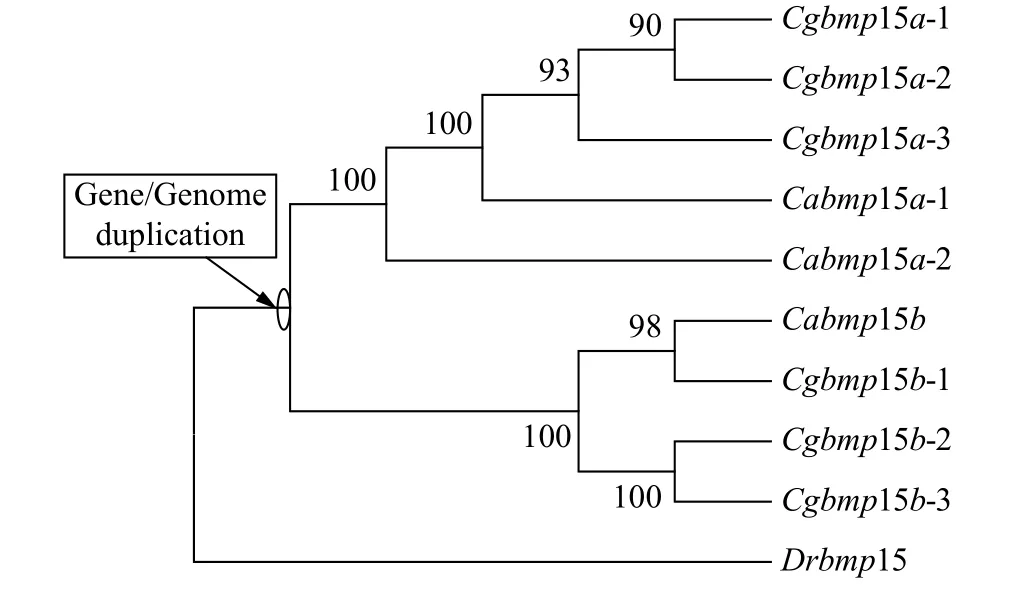
圖3 ML法構建的銀鯽、鯽和斑馬魚bmp15基因的系統進化樹Fig. 3 The phylogenetic tree of gibel carp, crucian carp, and zebrafish bmp15 based on ML method

圖4 Cgbmp15a和Cgbmp15b mRNA在銀鯽不同成體組織中的表達分析Fig. 4 Relative expression of Cgbmp15a and Cgbmp15b in different adult tissues

圖5 不同發育階段卵母細胞(A)以及DHP體外誘導GV1期卵母細胞成熟過程(B)中Cgbmp15a和Cgbmp15b的表達分析Fig. 5 Expression of Cgbmp15a and Cgbmp15b in oocytes during oogenesis (A) and GV1 oocytes induced by DHP in vitro (B)
3 討論
在本研究中, 我們首先從銀鯽F系中克隆了2個歧化的bmp15基因Cgbmp15a和Cgbmp15b, 它們各自具有一致性較高的3個不同等位基因。在魚類的進化歷程中, 除大部分魚類共同經歷的Ts3R外[23—25],多倍化是一個重要且頻繁發生的現象。多倍化根據染色體組的來源差異分為同源多倍化和異源多倍化, 其中, 異源多倍體通過種內或種間雜交產生。對鯉全基因組序列的解析表明在820萬年前發生了一次全基因組復制, 形成了異源四倍體[51]。對銀鯽2個歧化的Dmrt1基因的序列分析表明, 銀鯽在額外兩輪多倍化歷程中, 經過大約1849萬年前的一次早期多倍化事件, 形成了銀鯽、鯽和鯉的四倍體共同祖先; 而第二輪多倍化則發生在大約51萬年前, 在鯽這一分支上形成了六倍體銀鯽[44]。本研究克隆鑒定的6個銀鯽F系bmp15不同等位基因的序列比對表明, 銀鯽是一個異源六倍體。接著, 我們搜索了四倍體鯽以及人、小鼠、雞、爪蟾、雀鱔、斑馬魚、稀有鯽、青鳉、羅非魚和河鲀的基因組中的bmp15基因, 結果表明, 在鯽基因組中有3個bmp15等位基因, 而在其他物種的基因組中, 均只搜索到一個bmp15基因。系統進化樹和bmp15相鄰基因的同線性分析均表明(圖2和圖3), 在銀鯽的進化歷程中發生了兩輪多倍化, 其中早期的異源多倍化整合了來自2個原始祖先的染色體組, 導致銀鯽和鯽的四倍體共同原始祖先基因組中包含bmp15a和bmp15b; 隨后第二輪同源多倍化最終導致銀鯽存在6個bmp15等位基因。
由于染色體不平衡和基因組的不穩定, 新形成的異源多倍體會發生復雜的、非孟德爾遺傳方式的基因組變化, 包括染色體重排[52,53]、不同來源的2個部分同源基因(Homeologs)的缺失和沉默[54—59]、偏性或顯性表達[59—61]等。相對于Cgbmp15a的3個BAC克隆上的鄰近基因的分布,Cgbmp15b的3個BAC克隆基因均缺失了leap2基因, 并且Cgbmp15b-2-BAC還缺失了nhsl2和slc38a7基因,Cgbmp15b-1-BAC還缺失了lppl和cyp2x10.2基因(圖2)。同時, 盡管在成體組織分布、不同發育階段卵子及孕酮激素DHP誘導卵母細胞成熟過程中,Cgbmp15a和Cgbmp15b具有相似的表達譜式, 但Cgbmp15a的表達水平均顯著高于Cgbmp15b的表達水平(圖4和圖5)。這表明銀鯽在形成異源多倍體后, 其基因組在隨后的二倍化過程中發生了復雜的變化, 包括不同來源的2個部分同源基因的缺失或偏性表達。
與其他物種bmp15在成體組織中的分布相同,銀鯽Cgbmp15a和Cgbmp15b均在卵巢中檢測到了高豐度的轉錄本, 同時Cgbmp15a也在肝臟、腦、下丘腦和垂體等組織中檢測到低豐度的mRNA存在。在不同魚類中,bmp15基因在卵子發生的不同階段表達水平的變化是不同的。譬如,bmp15在斑馬魚不同發育階段的卵母細胞中均有表達, 且表達水平無顯著性差異[11]。在歐洲海鱸中,bmp15在初級卵母細胞中高表達, 從卵黃生成時期開始顯著降低[12]。在黑鯛中,bmp15在早期卵母細胞中高表達,隨著卵母細胞的生長, 表達水平下降, 在卵黃發生的卵母細胞中基本檢測不到轉錄本的存在[17];bmp15在銀鯽Ⅰ期卵母細胞(0.025—0.062 mm)中表達水平最高, 在Ⅱ-Ⅳ期卵母細胞中維持著相對低豐度、穩定的轉錄本含量[14]。研究結果與已報道的銀鯽bmp15不同,Cgbmp15a和Cgbmp15b在卵母細胞發育過程中高表達于皮質泡期卵母細胞中(0.21—0.23 mm, 圖5A)。通過比較卵母細胞分期的直徑大小, 我們推測導致這種不同結果的原因可能是由于取材分析的銀鯽卵巢處于不同發育階段,或者銀鯽克隆系的不同。與人絨毛膜促性腺激素(hCG)體外誘導銀鯽卵母細胞成熟過程中, 銀鯽bmp15上調表達結果相同[14], 當我們采用DHP體外誘導銀鯽F系GV1期卵母細胞成熟時,Cgbmp15a在誘導6h后開始上調表達水平, 在誘導8h后達到最高表達水平, 隨后下降。而Cgbmp15b變化較小(圖5B)。前期的研究表明, 銀鯽GV1卵母細胞在DHP體外誘導4.5h后, 生發泡開始破裂; 誘導8h后, 超過90%的卵母細胞的生發泡均破裂[50]。結合Cgbmp15a和Cgbmp15b在成體組織及不同發育階段卵子中的表達差異(圖4和圖5A), 我們推測Cgbmp15a在銀鯽的卵母細胞發育和成熟中起著主要作用。Cgbmp15a和Cgbmp15b在銀鯽生殖中的功能歧化有待進一步的研究。
