裂腹魚指環蟲(Monogenea: Dactylogyridae)一新記錄種
2020-06-12 12:52:20喀迪爾丁艾爾肯郭愛民郝翠蘭張文潤容夢婕宋鵬慧
水生生物學報 2020年3期
喀迪爾丁·艾爾肯 郭愛民 郝翠蘭 張文潤 容夢婕 宋鵬慧 岳 城
(新疆農業大學動物醫學學院, 烏魯木齊 830052)
斑重唇魚(Diptychus maculatesSteindachner,1866)和新疆裸重唇魚(Gymnodiptychus dybowskiiKessler, 1874), 分別隸屬于鯉形目(Cypriniformes),鯉科(Cyprinidae), 裂腹魚亞科(Schizothoracinae)[1],是伊犁河水系中具有代表性的土著種和生態敏感種[2], 二者分別為自治區I級和II級水生野生重點保護動物[3]。
指環蟲是一種常見的魚類寄生蟲病原, 其形態多樣, 種類豐富, 隸屬于單殖吸蟲綱、指環蟲目、指環蟲科[4], 廣泛寄生于淡水魚類, 宿主種類達8科194屬, 而其中約95%的種類寄生于鯉科(Cyprinidae)魚類, 表現出極強的宿主特異性[5]。指環蟲屬為單殖吸蟲綱中種類最多的屬, 全世界已報道970余種, 我國記載有400余種[6], 其中裂腹魚寄生指環蟲全球已報道約16余種, 我國現記錄有強壯指環蟲(D. pollensZhang et Gou, 1981)、裂腹指環蟲(D.schizothoraxiMa et Zhao)、塔里木指環蟲(D. talimuensisLong et Ma)、塊莖指環蟲(D. lumpycirrusMa et Zhao)、拉薩指環蟲(D. lhasaensisYao et Wang,1997)、裸裂尻指環蟲(D. SchizopygopsisYao et Wang, 1993)、四川指環蟲(D. sichuanensisMa et Long)等7種指環蟲, 均寄生于裂腹魚屬的裂腹魚類,而斑重唇魚和新疆裸重唇魚所屬的兩屬裂腹魚并未有相關記載[4,7,8]。
我們在進行裂腹魚寄生蟲病原區系調查時, 在伊犁河流域鞏乃斯段的斑重唇魚和新疆裸重唇魚鰓絲上發現寄生有大量指環蟲。結合形態學和分子生物學方法對其進行形態鑒定和系統發育分析,并查閱了國內外文獻, 未發現相關記述, 依據采集地命名重唇魚指環蟲(Dactylogyrus diptychusn.sp.)。正模標本GNS20181101和副模標本GNS2018 1102-03分別保存于新疆農業大學動物醫學學院標本館和寄生蟲研究室。
1 材料與方法
1.1 寄生蟲樣品采集
斑重唇魚和新疆裸重唇魚[9]采自于伊犁河上游鞏乃斯河, 兩種裂腹魚各30余尾。對魚類標本編號,測量體高和尾叉長、稱重并記錄相關數據。依照《魚病調查手冊》[10]的方法對魚進行解剖, 分別將每條魚的鰓部剪下, 逐個在解剖鏡下進行鏡檢, 發現寄生蟲后將其吸取到盛有清水的小皿中, 沖洗干凈后, 將其吸取到1.5 mL的EP管中, 滴加75%和95%濃度的酒精保存, 分別用于標本制作和DNA提取。
1.2 寄生蟲的形態學鑒定
在解剖鏡下挑取形態完整的新鮮蟲體, 用4%聚乙烯乳酸酚[11]固定并封片, 在光學顯微鏡(Nikon E2000)下觀察其幾丁質結構, 并通過EZMET軟件測量其中央大鉤、聯結片、交接器等幾丁質結構, 最后繪制特征圖。
1.3 DNA提取、擴增及測序
取95%無水乙醇固定的蟲體樣本若干只, 再次沖洗后置于TE緩沖液(pH 8.0)中浸泡過夜后, 吸去TE緩沖液, 分別裝入1.5 mL的EP管中, 用Trans-Gen動物組織基因組DNA提取試劑盒提取蟲體DNA, 在-20℃冰箱中保存。
選取18S-ITS1-5.8S rDNA序列作為目的序列,擴增引物參照?imková等[12]。引物由上海生工生物工程服務有限公司合成, 反應體系為50 μL, 具體反應條件和引物序列見表1。
PCR產物經1%瓊脂糖電泳分析并切取目的條帶后, 用TransGen公司的膠回收試劑盒進行回收和純化, 回收后的目的片段連接到pMD18-T載體進行TA克隆, 再轉化到E.coliDH5α中, LB平板培養,PCR檢測成功后送上海生工生物工程服務有限公司測序。

表1 PCR引物及反應條件Tab. 1 PCR primers and reaction conditions
1.4 系統發育分析
測序結果與GenBank公布的已知序列進行比對并下載相關序列(表2), 用ClustalⅩ軟件進行序列校正后, 采用MEGA6.0軟件計算所得序列中各堿基含量及變異情況, 同時計算遺傳距離(P-distance)。系統發育分析采用最大似然法(Maximum Likelihood, ML)和貝葉斯法((Bayesian Inference, BI), 以Ancyrocephalus percae作為外類群, 建樹采用Modeltest程序進行AIC檢驗選擇最適進化模型, ML樹和BI樹最適模型均為GTR+G+I, 在MEGA6.0軟件中構建ML樹, 系統樹各分支的相對支持率采用自展檢驗分析(Bootstrap) 進行檢驗, 1000次重復取樣;利用MrBayes軟件構建BI樹, 設置代替模型為6(nst=6)、位點間變異模型為gamma(rates=gamma),運行6000000代蒙特卡羅模擬(Markov chain Monte Crlo; MCMC), 每100代取樣1次, 各分支的置信度以后驗概率(Posterior probability)來表示[13]。
2 結果
斑重唇魚共采集32尾, 均有指環蟲寄生, 自然感染率約為100%, 每尾魚感染1—20枚蟲體, 每片鰓感染0—6枚, 感染強度為12.62; 新疆裸重唇魚共采集34尾, 均有指環蟲寄生, 自然感染率為100%,每尾魚感染1—24枚蟲體, 每片鰓感染0—7枚, 感染強度為8.67。兩種宿主平均感染強度為10.58。
2.1 蟲體的形態學特征
基于8個封片標本, 測量單位為微米(μm)。蟲體呈扁平長條狀(圖1), 體長1171(943—1385), 體最大寬100(85—120); 眼點兩對; 咽近乎呈圓形, 大小51(36—62)×49(36—60)。后吸器長86(65—114), 寬138(108—162), 包含1對中央大鉤、7對邊緣小鉤、1個背聯結片和1個腹聯結片。邊緣小鉤發育良好,可明顯區分鉤尖、鉤柄和柄軸, 第1對和第7對較大,全長51(47—56); 其余5對等大, 全長46 (41—51)。7對邊緣小鉤的鉤尖長5.8(5—8), 鉤柄長11(8—17),柄軸長30(23—36)。中央大鉤全長55(51—60), 基部長53(51—56), 鉤尖長14(13—16), 內外突粗壯,內突長14(11—18), 外突長12(10—15)。背聯結片呈粗壯“一”字型, 大小14(10—16)×78(74—83), 腹聯結片短小, 中部呈峰狀隆起, 大小12(11—16)×33(26—50)。交接器由交接管和支持器組成, 交接管基部膨大, 管細長, 一分為二, 末端形成圓環, 管長36(33—44)。支持器由兩部分組成, 上半部分形成半環繞交接管的柄狀結構, 下半部分形成“Y”狀結構, 與交接管底部相連, 支持器長26(23—31)。卵呈橢圓形, 大小75(64—82)×58(54—61), 陰道口開于蟲體左側。

表2 用于建樹的指環蟲種類及ITS1序列的GenBank登錄號Tab. 2 The Dactylogyrus species used for building trees and the GenBank accession number of the 18S-ITS1-5.8S rDNA sequence
2.2 蟲體的分子鑒定及系統發育分析
18S-ITS1-5.8S序列長度為996 bp, 其中保守位點515個, 變異位點459個, 包括336個簡約信息位點。A、T、G、C堿基的平均含量為23.52%、26.62%、27.65%、22.21%, A+T的含量高于G+C的含量。Blast比對顯示與D. cryptomeres相似度為最高92%,與其他指環蟲相似度較低, 如與D. lamellatus、D.squamous、D. tuba、D. zandti相似度為79%, 與D.varius、D.benhoussai、D.dyki相似度為73%。
基于18S-ITS1-5.8S rRNA基因序列構建的BI系統發育樹顯示(圖2), 系統發育樹分為三個分支,D.prespensis、D. malleus等多數指環蟲聚為一支,D.diptychus與D. cryptomeres聚為一個單獨分支,D.anchoratus、D. formosus等四種指環蟲聚為一支,位于進化樹基部。ML樹(圖3)與BI樹在第一個分支上略有差異外, 其余兩個分支結果一致, 說明D.diptychus與D. cryptomeres屬內親緣關系最近。
3 討論
指環蟲因其具有較強的宿主特異性, 通過宿主進行初步鑒定也是較常見的方法之一。本研究根據其形態學特征確定了本種隸屬于指環蟲屬后, 根據宿主情況, 將本種與現已記載的裂腹魚指環蟲進行了形態學特征的比較。裂腹魚寄生指環蟲全球現已報道16余種, 大部分均寄生于裂腹魚屬的裂腹魚類, 而寄生于新疆裸重唇魚, 斑重唇魚和同屬宿主的指環蟲僅有兩種, 分別為Dactylogyrus driagini(Bychowsky, 1936)和Dactylogyrus simplex(Bychowsky, 1936)[8], 故先與上述兩種指環蟲進行了對比。
本種與D. driagini交接器結構較為相似, 兩者的交接器上端均伸入支持器上端, 下端均形成圓環狀。但本種支持器上端由兩個底部相連的柄狀結構形成“V”狀, 環繞交接器的上端。而后者只形成單一的柄狀結構, 交接器上端直接穿過該部分, 同時本種支持器分為上下兩部分, 下部分形成“Y”狀與上部分支持器相連, 而后者支持器是完整的一體結構。本種與D. driagini后吸器均由一對中央大鉤、7對邊緣小鉤和兩聯結片組成, 本種中央大鉤內外突不太明顯, 而后者內外突分葉很明顯; 本種邊緣小鉤可明顯區分鉤尖, 鉤柄和柄軸, 而后者區分不明顯; 同時, 本種邊緣小鉤長平均為46 μm, 同時第1對和第7對較大, 達51 μm, 而后者邊緣小鉤為19—26, 明顯小于前者。而將本種與D. simplex進行形態學特征的對比, 發現二者不僅是在交接器還是后吸器形態上都存在明顯差異。
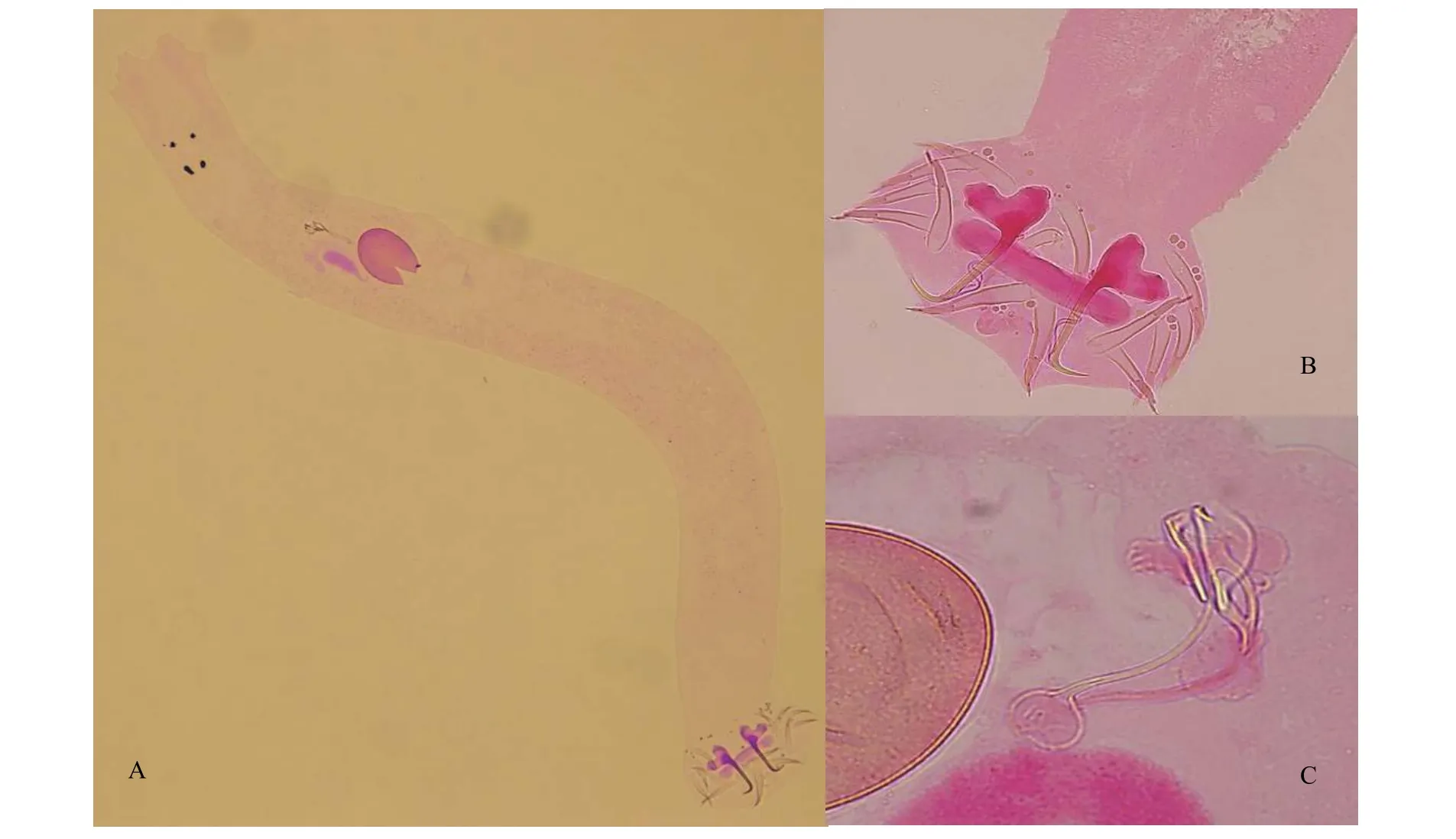
圖1 重唇魚指環蟲的形態特征Fig. 1 Morphological characteristics of Dactylogyrus diptychus n. sp.
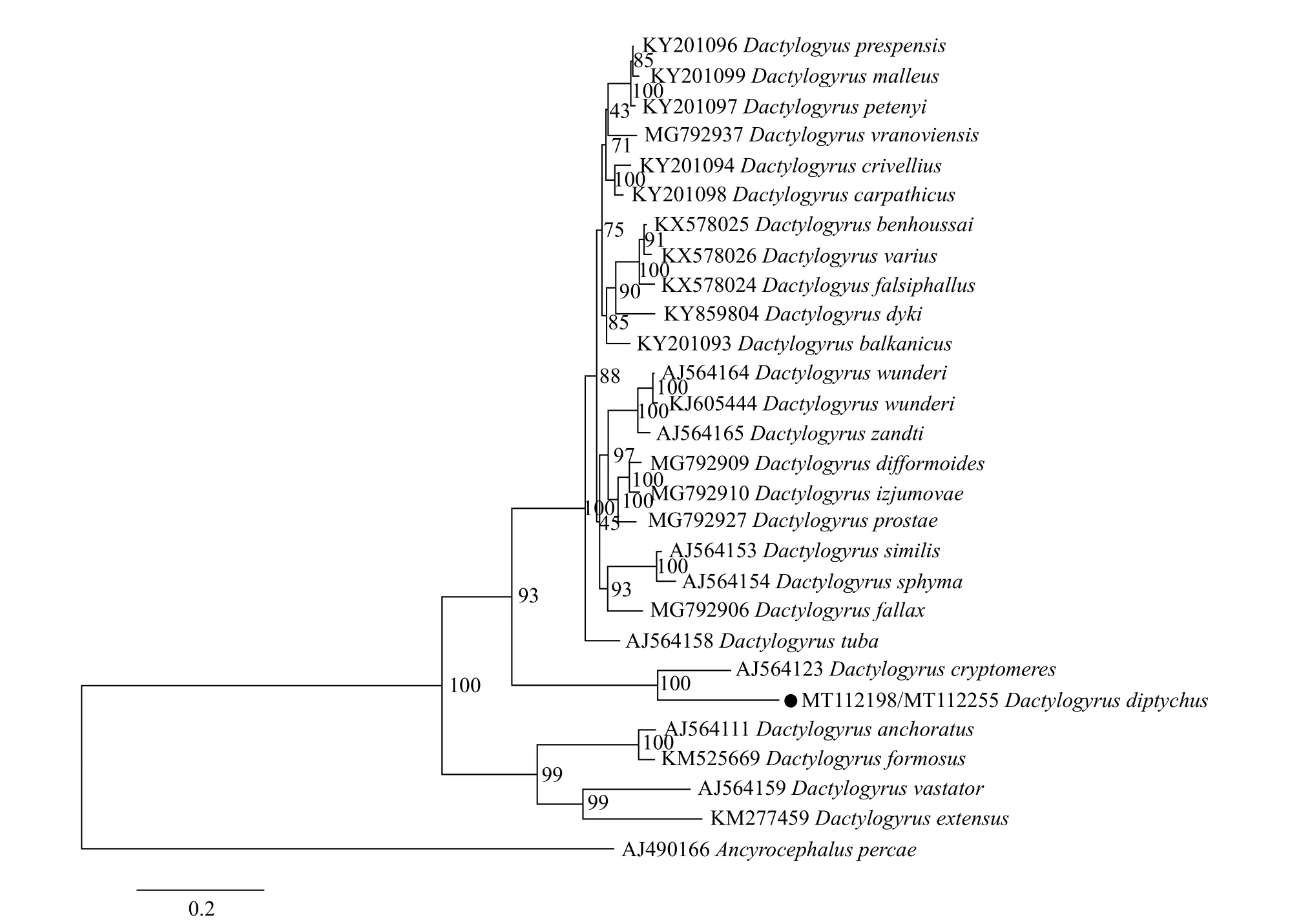
圖2 基于18S-ITS1-5.8S rDNA基因序列構建的貝葉斯法(BI)系統發育樹Fig. 2 Phylogenetic tree constructed via the Bayesian inference (BI) method based on 18S-ITS1-5.8S rDNA gene sequences

圖4 D. diptychus (A)、D. driagini (B)、D. longicopula (C)和D. simplex (D)幾丁質結構對比Fig. 4 Comparison of the chitinous structures of D. diptychus (A), D. driagini (B), D. longicopula (C) and D. simplex (D)
同時, 將本種與其他裂腹魚指環蟲進行形態學特征比較時發現, 與寄生于伊犁裂腹魚的Dactylogyrus longicopula(Bychowsky, 1936)形態類似, 但存在以下明顯的區別: 本種中央大鉤內外突非明顯分葉, 內外突大小基本相同, 而D. longicopula內外突明顯分葉, 且內突遠遠大于外突; 本種與D. longicopula均具兩聯結片, 前者腹聯結片粗短, 中部呈峰狀隆起, 而后者腹聯結片兩端細長, 中間呈三角狀隆起; 本種邊緣小鉤鉤尖與鉤柄連接處無刺狀突起, 而后者邊緣小鉤同部位均有刺狀突起, 同時度量值也存在較大差異; 本種支持器前端較扁平, 而后者形成兩個分化明顯的鉤, 同時前者支持器上下部分連接處呈“Y”狀, 后者為卷曲狀。
綜上所述, 重唇魚指環蟲(Dactylogyrus diptychusn. sp.)與上述三種裂腹魚指環蟲近似種在形態特征上均有明顯的差別, 結構比較見圖4。
傳統形態學分類不管是在早期還是現在, 都為單殖吸蟲的分類鑒定做出了巨大貢獻[14,15]。然而在實際鑒定過程中受一些不可控的變量因素的影響, 傳統的形態學分類顯現出了其不足之處[16,17],單純的依賴傳統的形態學進行分類顯得日益困難,尤其是對同屬的相似種而言[18]。單殖吸蟲特異性分子標記目前以rDNA和ITS為主, 主要包括18S、5.8S、28S、ITS1、ITS2, 其中18S rDNA、ITS1、5.8S rDNA組成一個轉錄單元, 形成一個前體RNA。其中18S rDNA中的不同區域在進化上比較保守, 而ITS1屬于非編碼區序列, 進化速度快, 具有可變性并且變異的速度在不同的物種間也存在著較大的差異, 同時, 該基因中保守區占18S rRNA基因總長度的一半以上, 因此18S-ITS1-5.8S rDNA常用于單殖吸蟲, 特別是指環蟲的分類及系統發育研究[19,20]。本研究基于形態學結果, 選擇擴增了該蟲種的18S-ITS1-5.8S rDNA序列, 擴增后其結果在GenBank中進行比對, 與寄生于屬魚的隱藏指環蟲的相似性達92%, 且遺傳距離較其他指環蟲相比最小。構建的進化樹中兩者聚為一支, 置信度達100/99(BI/ML)。但基于形態學特征, 本種與隱藏指環蟲在后吸器和交接器形態上存在明顯差別, 同時隱藏指環蟲宿主為亞科的魚類, 與本種的兩個宿主非同科。其次, 裂腹魚類為高原冷水魚類, 分布于山區高原地帶, 而屬魚為溫水性魚類, 大多分布于平原地區, 兩種宿主魚類在生境上存在較大差別。因此結合形態學和宿主特點可證明, 兩種指環蟲在系統發育分析中雖顯示出了較高的親緣關系和序列相似度, 但并非同一種。
通過上述形態學分類和基于18S-ITS1-5.8S rDNA基因序列的分子生物學研究, 確定寄生于新疆裸重唇魚和斑重唇魚鰓上的待鑒定指環蟲為未記錄種, 按照宿主特點命名為重唇魚指環蟲(Dactylogyrus diptychusn. sp.)。研究結果豐富了我國單殖吸蟲名錄, 同時為伊犁河新疆裸重唇魚和斑重唇魚的資源保護提供了相關的病原基礎性資料。
