傷寒沙門菌非編碼RNA ArpH表達特征及功能初探
2020-06-18 01:10:00熊昌艷李雪嬌黃新祥
華中師范大學學報(自然科學版) 2020年3期
關鍵詞:水平
熊昌艷,李雪嬌,黃新祥
(江蘇大學醫學院,江蘇 鎮江 212013)
傷寒沙門菌(Salmonellaentericaserovar Typhi,S.Typhi)是一種重要的革蘭陰性人類病原菌,可通過多種毒力因子作用,引起輕度腹瀉甚至是嚴重的全身性感染.沙門氏菌毒力基因的集中區域被稱為沙門氏菌致病島(Salmonellapathogenicityislands,SPIs),可位于細菌染色體或質粒上.S.Typhi與宿主能產生交互作用,有些毒力基因能保護細菌免受宿主免疫系統的攻擊[1-2].
病原菌在感染過程中通過調控其毒力基因的表達,可以快速適應環境的變化.許多非編碼RNA(non-coding RNA,ncRNA)通過堿基配對相互作用形成與DNA或蛋白質的復合物,參與復雜的基因表達調節網絡.以前,許多ncRNA被認為是不參與蛋白質編碼的轉錄噪聲,然而近年研究已經證實,大多數ncRNA可通過調節細菌中許多基因的表達來發揮生理作用,包括ABC轉運系統、群體感應、氧化應激、耐酸性能和毒力等[3-4].
前期本實驗室對傷寒沙門菌野生株進行了全基因組序列分析,獲得了基因組結構精細圖.通過轉錄組序列分析發現有大量ncRNA序列位于一千余個基因的反義鏈上,有的拼接長度甚至超過了1 000 nt.ArpH就是新發現的ncRNA之一,前期研究確定其分子全長為3 508 nt,位于基因rpoH對側鏈,5′端位于yhhk起始密碼子上游411 nt,3′端位于rpoH起始密碼子上游238 nt,與rpoH完全重疊,為rpoH的反義RNA,因此命名為ArpH.研究發現ArpH高表達后能提高rpoHmRNA表達水平[5].RpoH是由rpoH基因編碼的σ32(σH),現已知腸道細菌的σ因子之間存在相互作用,如RpoE可促進RpoH和RpoS的表達[6-7].RpoH能激活小RNA的伴侶分子基因hfq的表達,而Hfq也可調節RpoH介導的環境應答反應.在沙門菌中也發現RpoH和RpoE可促進抗氧化作用[8].高滲應激可激活大腸桿菌和鼠傷寒沙門氏菌中依賴σE和σS的基因表達.σE是一種啟動一系列基因轉錄并對應激作出反應的細胞外因子.而σS是極端條件下細菌生存和發揮毒力所需的主要調節因子[7].另外,原核生物體內ncRNA的含量除了與其表達生成有關,與其降解也有重要關系.細菌體內存在多種RNase來降解RNA.本文通過研究σ因子和RNase對ArpH表達的影響,并結合全基因組芯片的結果,初步探討ArpH對細菌侵襲力和胞內生存力的影響.
1 材料與方法
1.1 材料
1.1.1 菌株和質粒 菌株:S.Typhi 野生株GIFU 10007 (WT),σE缺陷株 (ΔrpoE)[6],σS缺陷株 (ΔrpoS)[7],RNase III缺陷株(ΔRNase III)[9],RNase G缺陷株(ΔRNase G)[9],RNase E缺陷株(ΔRNase E)[9]由本實驗室保存,缺陷株使用自殺質粒法制備.arpH缺陷株 (ΔarpH),WT-pBAD (WT含pBAD/Myc-hisA),WT-pBAD-arpH(WT含pBAD-arpH) 由本實驗室制備[5].
質粒:pBAD/Myc-hisA (Ampr)購自Invitrogen公司,pBAD-arpH(pBAD/Myc-hisA連接了arpH序列) 由本實驗室制備[5].
1.1.2 主要試劑DNA聚合酶Taq、T4 DNA連接酶(Takara公司);質粒提取試劑盒、DNA凝膠回收試劑盒(AXYGEN公司);RNA提取試劑盒 (QIAGEN公司);PrimeScript逆轉錄試劑盒 (Takara公司);Cy3-dCTP、Cy5-dCTP(Amersham公司);傷寒沙門菌基因組芯片由本實驗室制備[10].
1.1.3 主要儀器電穿孔系統Gene Pulsero Ⅱ(Bio-Rad公司);PCR儀2720 Thermal Cycler (ABI公司);實時熒光定量PCR儀CFX96TMReal-Time System (Bio-Rad公司);基因芯片雜交儀hydridiser HB-3d(Roller-Blot公司);基因芯片掃描分析系統Genepix personal 4100A (Axon Instruments公司);CO2細胞培養箱MODEL 3111 S/N 305713-7757(Thermo公司).
1.1.4 引物利用Oligo6軟件,根據arpH的核苷酸序列設計qRT-PCR特異性引物ArpH-qF和ArpH-qR.5S rRNA為內參.qRT-PCR引物見表1.

表1 本研究中所用引物序列Tab.1 The sequence of primers used in this study
1.2 方法
1.2.1 細菌培養及RNA提取 將單克隆細菌于LB液體37 ℃過夜培養,按1∶100轉接于相同條件LB液體培養基,需要時加入0.2%L-阿拉伯糖誘導30 min.采用TRIzol法提取細菌總RNA后,用DNase I(RNase free)去除混雜的少量DNA,酚仿-乙醇法純化.取適量RNA用1.2%的瓊脂糖凝膠電泳檢測RNA質量.
1.2.2 qRT-PCR分析 各取純化的總RNA 4 μg,用PrimeScript逆轉錄試劑盒 (Takara公司)分別反轉錄生成cDNA后,再進行qRT-PCR.內參基因為5S rRNA,用于qRT-PCR分析的引物序列如表1所示.每次實驗均按照說明書進行3次.
1.2.3 全基因組芯片檢測 將WT-pBAD、WT-pBAD-ArpH培養至對數期 (A600為0.4) 后加入2 g·L-1L-阿拉伯糖誘導30 min,然后在氧應激下(H2O25 mmol·L-1)的LB中37 ℃振蕩(250 r·min-1)培養4 h,總RNA用RNA提取試劑盒提取.取上述菌株的總RNA各20 μg進行逆轉錄,用隨機引物N9、傷寒沙門菌GNP引物和PrimeScript逆轉錄試劑盒,反轉錄生成cDNAs的同時摻入Cy3-dCTP或Cy5-dCTP,將cDNAs進行相互配對并標記,經提純后與S.Typhi基因組DNA芯片進行雜交,清洗后用雙通道熒光掃描,將熒光信號進行數據轉換并經標準化處理后行統計分析[11].
1.2.4 HeLa細胞侵襲實驗 用含10%小牛血清的RIPM-1640于37 ℃、5% CO2的培養箱中培養HeLa細胞,實驗前24 h按每孔2×105接種于24孔.將S.Typhi野生株(WT)、ArpH缺陷變異株(ΔarpH)、空質粒對照株(WT-pBAD)和ArpH高表達菌株(WT-pBAD-ArpH)培養至對數生長期,按20∶1的細菌與細胞比(multiplicity of infection,MOI)分別加入24孔板,培養90 min(37 ℃、5% CO2),以預熱的PBS洗細胞3次后,加入0.1% PBS-DOC作用10 min破膜,將破膜液中的細菌收集起來涂抹于LB平板,經37 ℃溫箱培養12 h后計算菌落數(t0);平行板每孔再加入終濃度為100 μg·mL-1的慶大霉素,再培養(37 ℃,5% CO2),讓細菌侵襲HeLa細胞,90 min后加入1% Triton X-100作用10 min破膜,收集破膜液中的細菌,涂LB平板,37 ℃過夜培養后計數菌落數(t90).以菌落數t90/t0的比值作為細菌的侵襲力指標.
1.2.5 巨噬細胞內增殖試驗 將THP-1單核細胞用含10%小牛血清的RIPM-1640培養基于24孔板培養(37 ℃,5% CO2),加100 ng·mL-1PMA(phorbol 12-myristate 13-acetate)培養24 h,誘導成巨噬細胞.將WT、ΔarpH及WT-pBAD、WT-pBAD-ArpH培養至對數生長期,WT-pBAD、WT-pBAD-ArpH加入0.2% L-阿拉伯糖誘導1 h,按細菌與細胞比20∶1,分別加入含巨噬細胞的24孔板,感染30 min后,每孔再加入慶大霉素(終濃度100 μg·mL-1)1 h以殺死胞外菌,吸取上清,用PBS將每孔吹洗3次.一部分孔加入1 mL 0.5%(V/V)Triton裂解細胞,反應10 min后將裂解物涂于LB平板,37 ℃過夜培養后的單克隆數用t0表示基礎細菌吞噬水平;另一部分孔細胞繼續培養12或24 h后同樣以1 mL 0.5%(V/V)Triton裂解細胞,過夜培養后計算克隆數代表胞內細菌增殖水平(以t12或t24表示).細菌在巨噬細胞內生存12 h及24 h的增殖力分別以t12/t0及t24/t0表示.
1.2.6 統計學分析用平均數±標準差表示3個獨立實驗的平均值.利用SPSS 22.0軟件進行統計分析,組間差異采用方差分析,P<0.05表示差異有統計學意義.
2 結果
2.1 RpoE和RpoS在不同應激條件下對ArpH表達的影響
應用qRT-PCR分析野生株和rpoE、rpoS缺陷株在等滲非應激條件和環境酸、氧和高滲應激下ArpH的mRNA表達水平.結果顯示(圖1),ArpH表達在酸應激下明顯下降,在氧和高滲應激下略有增高.在rpoE缺陷株中,ArpH的表達水平在各種條件下均有明顯下降.在rpoS缺陷株中,ArpH表達水平在氧應激時下降較為明顯.
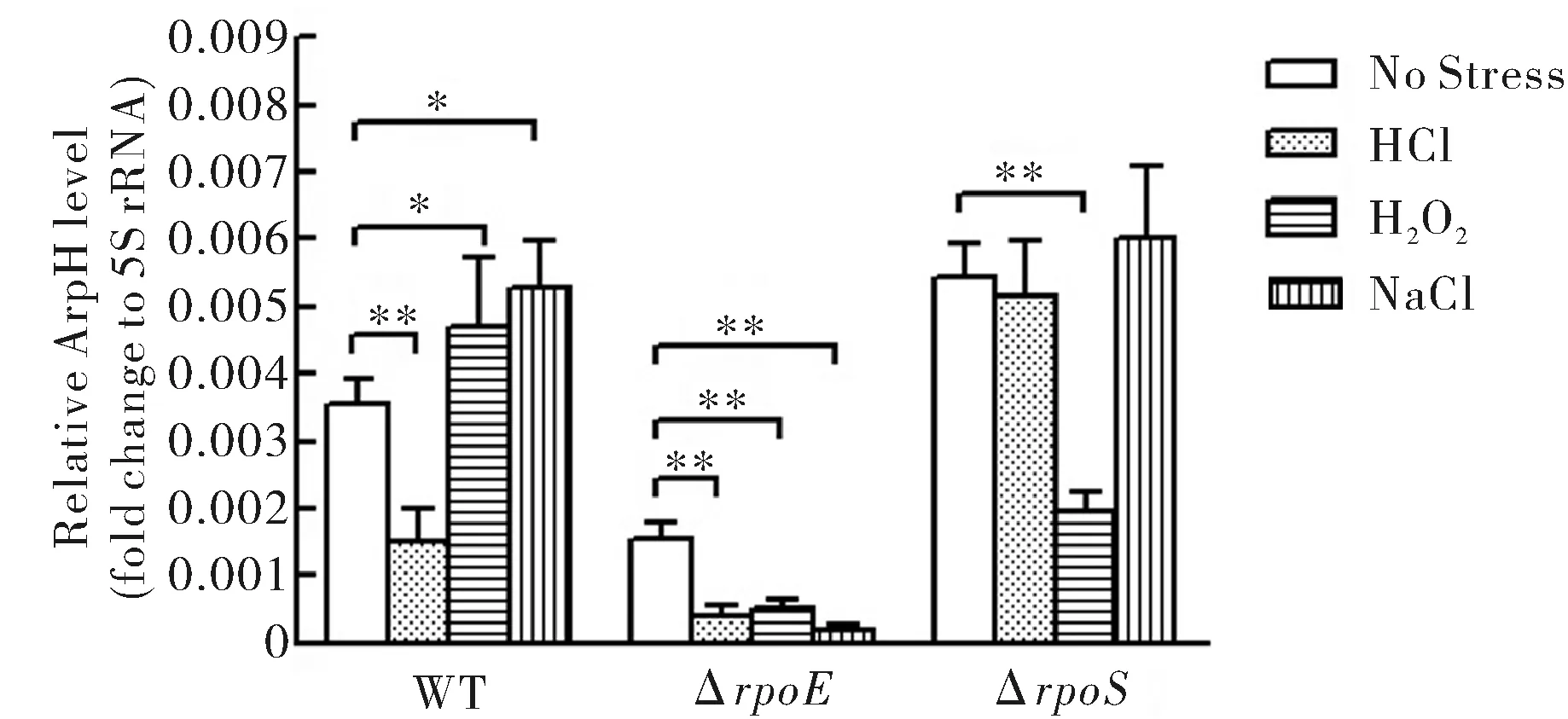
No Stress:無應激;HCl:酸應激;H2O2:氧應激;NaCl:高滲應激.*為P<0.05,**為P<0.01圖1 qRT-PCR檢測在WT、ΔrpoE和ΔrpoS中不同應激下ArpH的mRNA表達水平Fig.1 The mRNAexpression of ArpH in WT and mutants at different stresses
2.2 RNase III、RNase G和RNase E對ArpH降解的影響
S.Typhi野生株、RNase III、RNase G、RNase E缺陷變異株培養至A600=0.8后,經利福平處理0、5、10、20 min,RNase III缺陷株中ArpH水平明顯高于野生株,而RNase G和RNase E這兩種RNA酶缺陷株中ArpH水平無明顯差異(圖2).
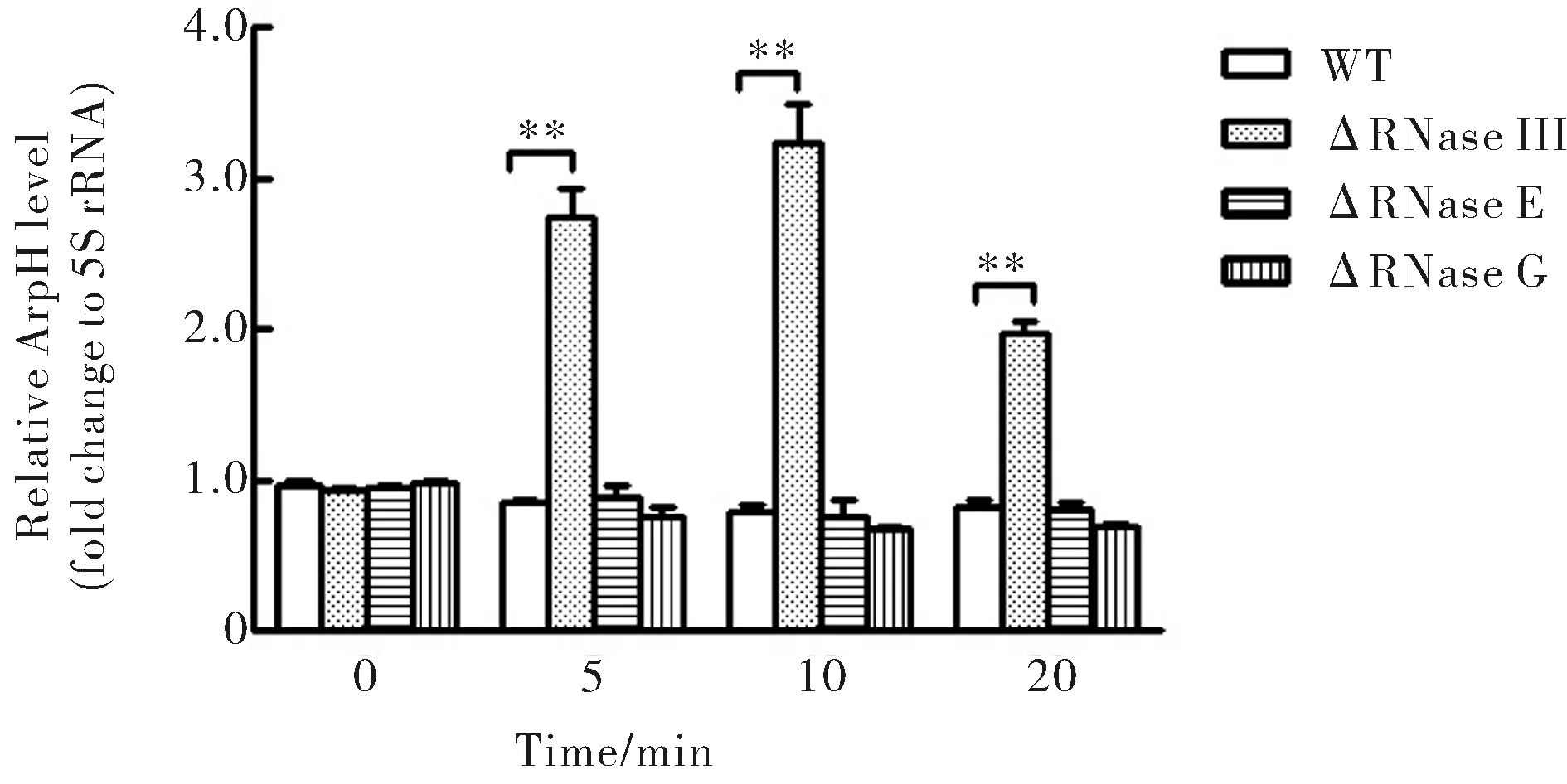
圖2 WT、RNase III、RNase G和RNase E缺陷株中不同時間的ArpH水平(**為P<0.01)Fig.2 The ArpH levels at different times in WT,RNase III,RNase G and RNase E deficient strains
2.3 ArpH高表達菌株氧應激下的基因表達譜分析
將空質粒對照株(WT-pBAD)和ArpH高表達菌株(WT-pBAD-arpH)培養至對數期(A600為0.4)后加入質量濃度為2 g·L-1的阿拉伯糖誘導30 min,然后在氧應激下的繼續培養4 h.提取RNA后,經熒光標記、逆轉錄、芯片雜交等步驟后進行全基因組芯片分析.數據分析結果顯示(表2),和空質粒對照株比較,ArpH高表達菌株中基因表達有顯著差異的基因共113個,包括89個上調基因和24個下調基因.上調的基因主要涉及鞭毛相關基因flhDC、fliACJHLSZ和鞭毛基體修飾蛋白基因flgBCDEFHIJKL,超氧化物歧化酶sodB等.下調的基因主要涉及致病島1效應蛋白prgHIK、sipACD,侵襲相關基因iagA、invAG、tviACDE、spaIKMN和營養代謝相關基因astB、msyB和glpD等.

表2 空質粒對照株和ArpH高表達菌株在氧應激下的部分差異表達基因Tab.2 Differential genes in control strain and ArpH overexpressing strain of S.Typhi induced by H2O2
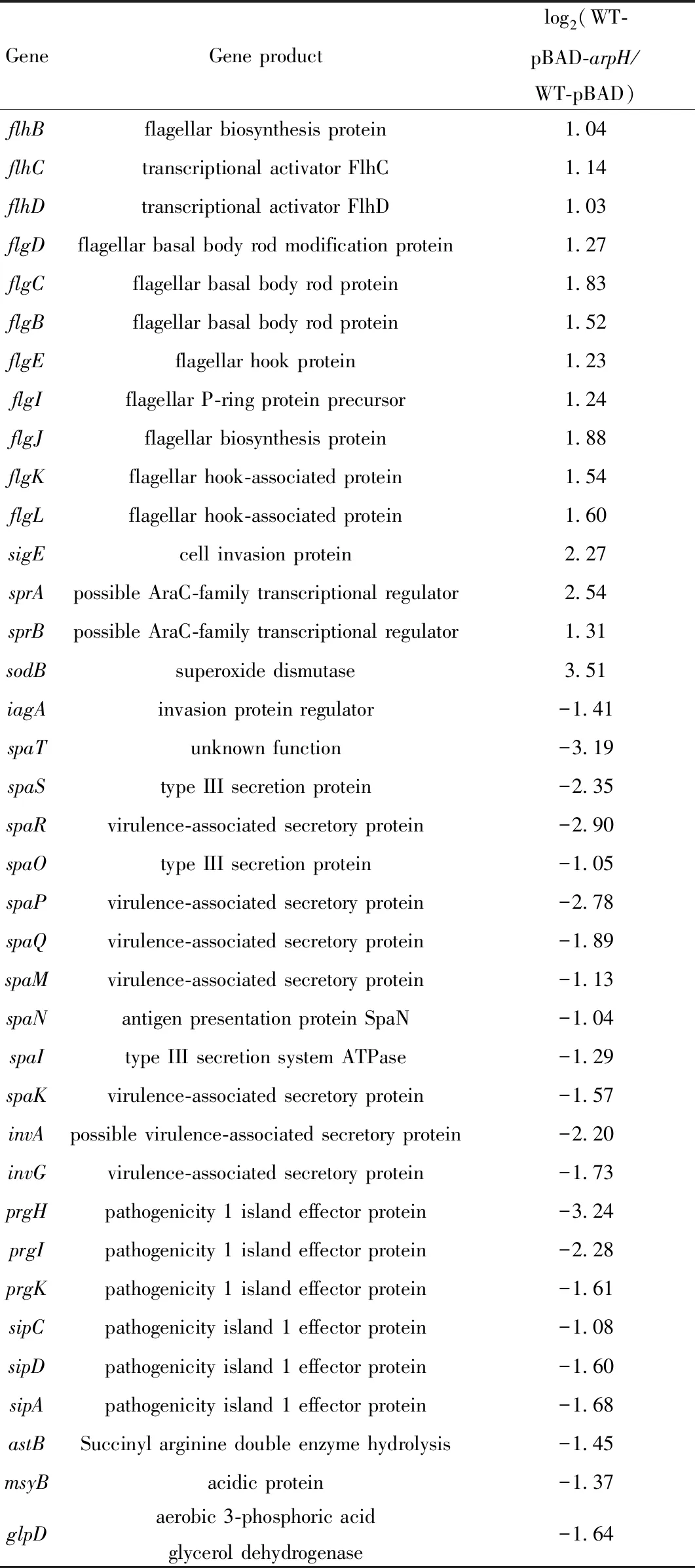
續表2
2.4 ArpH對細菌侵襲HeLa細胞活性的影響
為了探討ArpH對傷寒沙門菌侵襲上皮細胞的能力,應用S.Typhi野生株(WT)、ArpH缺陷變異株(ΔarpH)、空質粒對照株(WT-pBAD)和ArpH高表達菌株(WT-pBAD-arpH)對HeLa細胞進行侵襲實驗(圖3).當arpH基因缺失后,缺陷株的侵襲水平明顯強于野生株,約為野生株的1.5倍,其差異具有統計學意義(P<0.05).與空質粒對照株相比,ArpH高表達菌株的侵襲能力下降且差異具有統計學意義(P<0.05).
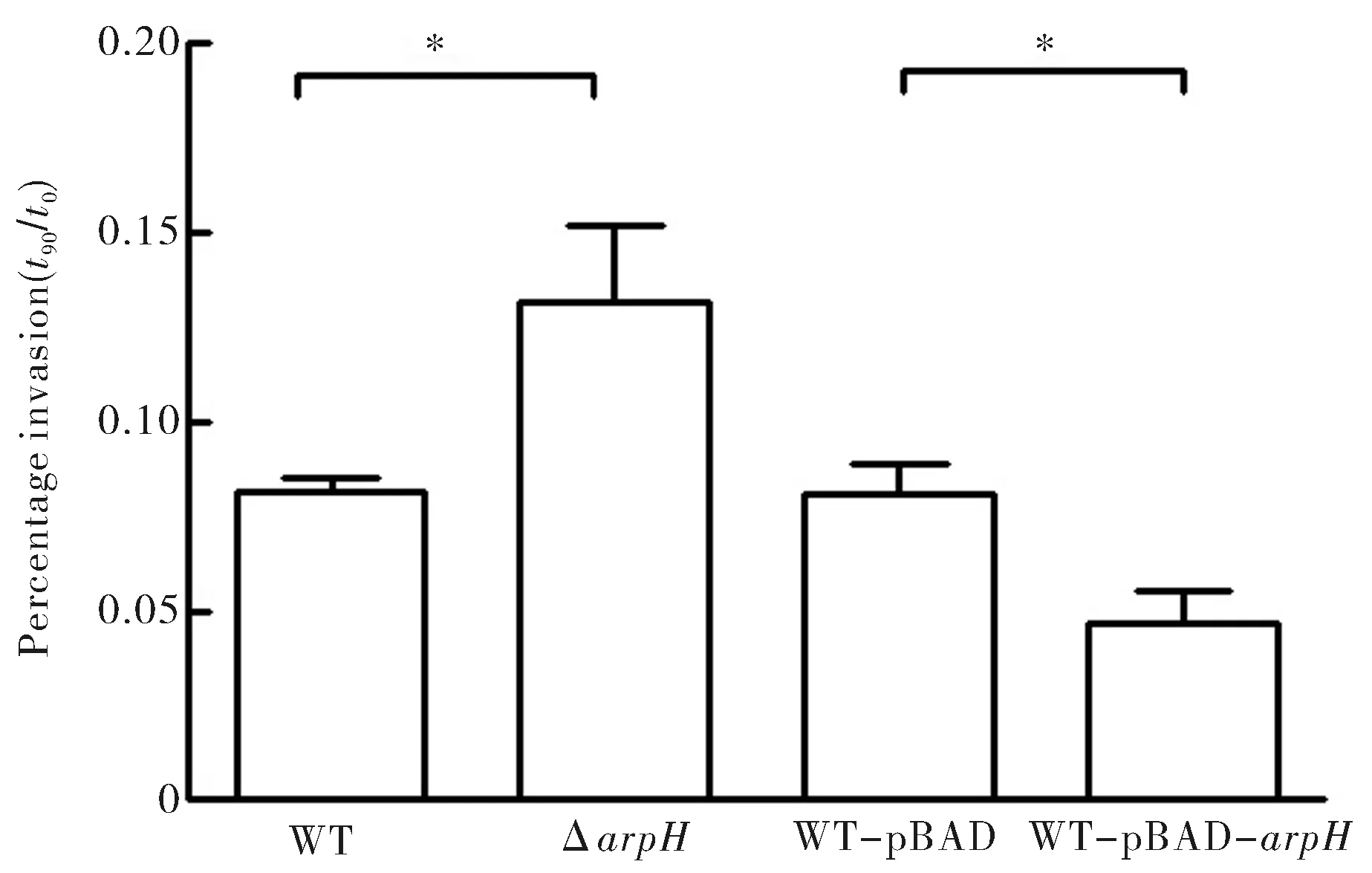
圖3 ArpH各菌株對HeLa上皮細胞侵襲力的影響(*為P<0.05)Fig.3 Invasion of HeLa cells by wild-type,ΔarpH,and WT-pBAD-arpH strains
2.5 ArpH對細菌在THP-1胞內生存力的影響
為了觀察ArpH是否會對細菌在THP-1細胞內生存產生影響,應用WT、ΔarpH、WT-pBAD和WT-pBAD-arpH菌株進行THP-1細胞內生存實驗.與時間零點的胞內細菌初始水平相比,野生株、空質粒對照株在感染12 h和24 h后的細菌水平均達到3倍以上,且此兩種菌株的胞內增殖趨勢沒有明顯差異.然而,ArpH缺陷變異株在感染12 h和24 h后,其胞內細菌水平則分別為初始水平的1.9 ± 0.3倍和1.2 ± 0.2倍,ArpH高表達菌株在感染12 h和24 h后,其胞內細菌水平則分別為初始水平的5.1 ± 0.5倍和3.7 ± 0.3倍(圖4).另外,細菌在感染24 h后的胞內水平均較12 h有所下降,可能是由于細菌的感染導致部分細胞死亡并裂解,其胞內的細菌被釋放到胞外被慶大霉素殺死.
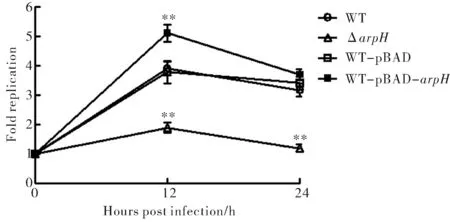
圖4 傷寒沙門菌在巨噬細胞內的生存力比較(**為P<0.01)Fig.4 The intracellular survival ability comparison of S.Typhi in macrophages
3 討論
ncRNA的功能是目前ncRNA研究領域的熱點.ncRNA可以在DNA水平、mRNA水平和蛋白質水平上發揮作用[12].現已發現ncRNA具有廣泛的生物學功能,包括作為信號分子、分子誘餌作用、引導功能和參與組成染色質支架等4個方面[13].某些ncRNA的生成具有組織和時間特異性,它們的表達是細胞針對特定刺激(如細胞應激和溫度等)引起反應的產物[14],這些ncRNA可作為信號分子引起細胞功能的變化.在此,本文初步探討了前期發現的ncRNA ArpH的部分表達特征和功能.
細菌屬原核生物,其基因表達均是由σ因子介導轉錄起始[15].許多ncRNA表達受應激反應σ因子的正向調節,如RpoE和RpoS、雙組分系統PhoP/Q[16].ncRNA通過作用于自己的調節子來進行反饋調控[17].腸道細菌在高滲應激或穩態期條件下,可以誘導產生σE、σH和σS.σH是大腸桿菌中首先被發現的σ因子.rpoH有4個啟動子,其中一個啟動子rpoH3P可被σE激活[18].本研究模擬體內酸、氧和高滲應激,檢測了σ因子對ArpH表達的影響,結果顯示在rpoE缺陷株中,ArpH的表達在各種應激條件下均有明顯下降.ArpH作為rpoH對側鏈的反義RNA[5],提示ArpH的轉錄可能通過RpoE激活RpoH而介導.hfq編碼的蛋白是一種能與RNA結合的調節性蛋白,而σH能激活hfq的表達.反之,Hfq也可調節σH介導的胞質應答反應.同時,Hfq對于rpoS的翻譯也起決定作用.在沙門菌中也發現,通過增加σS的水平,σH和σE可促進抗氧化作用[19].σS主要與穩態期和應激條件下基因的表達相關.前期研究顯示ArpH在對數晚期至穩態期的表達量最高[5],表明ArpH的表達有可能受到σS的調控.本研究在rpoS缺陷株中發現ArpH的表達水平在氧應激時下降較為明顯,這可能是由于σS表達的降低引起σH表達水平的下降,從而導致σH抗氧化作用的減弱,位于rpoH對側鏈的反義RNA ArpH也隨之下降.這些調控機制的深入研究可能為控制傷寒沙門菌感染提供有效的靶向基因.
RNase III是一種作用于雙鏈 RNA 的核糖核酸內切酶,對 ncRNA 和靶 mRNA 配對的雙鏈區進行切割,可以同時降解 ncRNA 和其配對的靶 mRNA.而RNase G和RNase E一樣,主要降解單鏈,并且多水解5′末端含有單磷酸基團的 RNA 和富含 AU 堿基的區域[20-21].ncRNA能保護靶基因mRNA來對抗RNase E的降解[22].本研究分析了RNase III,RNase G和RNase E對ArpH的降解特性.結果顯示這些RNA酶缺陷株經利福平處理后,RNase III缺陷株中ArpH水平明顯高于野生株,而RNase G和RNase E這兩種RNA酶缺陷株中ArpH水平無明顯差異,以上表明RNase III可能是ArpH的主要降解酶.ArpH主要以ArpH/RpoH雙鏈形式被降解,提示ArpH可能參與調控RpoH mRNA的穩定性來影響靶向mRNA的表達.
基因芯片技術可以對一些ncRNA的主要靶基因進行鑒定[23].前期研究發現ArpH的高表達能促進S.Typhi在氧應激條件下的生長[5].因此,本研究在氧應激下對ArpH高表達株應用全基因組芯片技術進行了基因表達譜分析,結果顯示超氧化物歧化酶sodB基因表達上調明顯.而sodB對S.Typhi在巨噬細胞中的生存力具有重要作用.這進一步驗證了ArpH直接或間接調節氧應激環境下的某些基因表達.進一步實驗觀察了ArpH對S.Typhi在THP-1細胞內生存力的影響,結果發現ArpH可以提高S.Typhi在THP-1的胞內生存力,可能是影響S.Typhi細胞內生存的正向調節因子,提示ArpH可促進S.Typhi抵抗氧化損傷[24-25].另外,下調基因中包括致病島1效應蛋白prgHIK、sipACD,侵襲相關基因如iagA、invAG、tviACDE、spaIKMN等.這提示ArpH可能是影響傷寒沙門菌侵襲的負向調節因子.為此,本研究進行了HeLa細胞侵襲實驗.當arpH基因缺失后,菌株的侵襲水平高于野生株,約為野生株的1.5倍.與空質粒對照株相比,ArpH高表達株的侵襲能力下降,這表明ArpH可以減弱S.Typhi對上皮細胞的侵襲能力,可能是影響傷寒沙門菌侵襲的負向調節因子,這與芯片分析結果一致.ArpH高表達后能提高rpoHmRNA的表達水平[5].ArpH作為rpoH對側鏈的反義RNA,ArpH對rpoH表達的影響可能通過順式作用在轉錄后水平得以實現.σH可介導沙門菌SPI-1表達的負向調控,從而影響沙門菌對細胞的侵襲作用.有研究表明,σH分別在轉錄前和轉錄水平上負向調節HilD和HilA這兩種沙門菌SPI-1特異性的轉錄調節子[26].因此,ArpH可能通過rpoH來間接調控鞭毛、侵襲等基因的表達,其具體機制尚待進一步研究.基因芯片還篩選到諸多涉及營養代謝等功能的基因發生了變化,ArpH對這些基因的調控機制尚待研究.
本文探討了ArpH的轉錄和降解特性,進而分析了ArpH對傷寒沙門菌侵襲力的影響和對胞內生存力的作用,研究結果將增強對傷寒沙門菌在感染環境下的基因表達調節網絡和致病機制的認識,也為控制細菌感染提供了新的研究策略.
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
