甜櫻桃葡萄座腔菌的生物學特性研究
2020-06-19 02:20:50張嘉慧馬志輝張麗珍
食品工業科技 2020年10期
關鍵詞:生長
張 倩,李 陽,楊 埔,張嘉慧,馬志輝,張麗珍
(山西大學生命科學學院,山西省特色植物資源與研究利用重點實驗室,山西太原 030006)
櫻桃有較早的成熟期,素來有“春果第一枝”的美譽,食用價值高、經濟效益好[1-3]。但在櫻桃樹生長過程中經常會遭到一些病蟲害的影響,其中櫻桃流膠病就是櫻桃種植中常見的病害之一。櫻桃流膠病會危害枝干,造成主干、主枝甚至枝條流膠,導致樹勢衰弱,樹體抵抗力降低,果實產量和品質下降,嚴重時大枝枯死,甚至整株死亡[4]。意大利[5]、智利[6]等地均有發生。隨著櫻桃栽培面積的擴大,流膠病的發生也不斷呈上升趨勢,探明櫻桃流膠病的病因以及尋求流膠病的防治方法對促進櫻桃產業的發展具有十分重要的意義。
目前國內外對櫻桃流膠病的研究多側重于研究流膠的規律和其他相似核果類的致病菌。Bush[7]、Copes[8]、羅春香[9]和Saniewski[10]等多側重于研究流膠的爆發期及外部因素的影響。吳文能等[11]、Koh等[12]、Tang等[13]和吳小芹等[14]等發現Botryosphaeriadothidea是獼猴桃采后果實腐爛病、蘋果輪紋病及一些經濟林、用材林、綠化林木和較為珍稀樹種潰瘍的主要致病菌。但并未對造成甜櫻桃流膠的主要致病菌及侵染機理進行詳細的研究。Griffin等[15]是最早研究櫻桃流膠病的,但并未分離到引起櫻桃流膠的關鍵致病菌。張福興等[16]證實了甜櫻桃潰瘍型流膠病的致病菌屬為Botryosphaeria,但僅對此致病菌屬的致病性進行了簡單描述,并未對其生物學特性進行系統化研究。因此,本實驗在已成功分離純化并通過回接驗證一株能引起甜櫻桃流膠的病原菌SXYTLJ01的基礎上,對該菌株的生物學特性進行了更深入的探索。
本實驗以在山西榆次櫻桃園中甜櫻桃流膠病枝上分離得到的病原菌SXYTLJ01為對象,通過觀察其菌絲結構,不同脅迫條件下能否誘導產孢,其最適生長條件和致死溫度等方面,對該菌株的生物學特性進行了研究,以期為該菌種建檔提供有用信息,為明確病害的發病機理及防治提供試驗基礎與理論依據。
1 材料與方法
1.1 材料與儀器
病原菌SXYTLJ01菌株 從山西榆次櫻桃園的流膠枝條上分離純化得到;菌株保存在4 ℃的冰箱中,備用,在試驗前轉接馬鈴薯葡萄糖瓊脂培養基(PDA培養基配方[17]:馬鈴薯20%,葡萄糖2%,硫酸鎂0.2%,無水磷酸二氫鉀0.2%,蛋白胨0.3%,瓊脂2%),28 ℃暗培養,活化得到試驗用菌落。瓊脂 北京索萊寶科技有限公司;葡萄糖、可溶性淀粉、麥芽糖、乳糖、蔗糖 天津市風船化學試劑科技有限公司;蛋白胨、牛肉膏、硫酸銨、硝酸鈉、硝酸鉀 北京奧博星生物技術有限責任公司;硫酸鎂 天津市北辰方正試劑廠;無水磷酸二氫鉀 北京精求化工廠;乳酸石炭酚棉蘭染色液 北京索萊寶科技有限公司;Ezup柱式SK8259真菌基因組DNA抽提試劑盒、Taq Plus DNA聚合酶、10X PCR Buffer(含Mg2+)、dNTP(10 mmol/L)、滅菌去離子水、6×DNA Loading Dye、DNA Ladder Mix(100~10000 bp)、DNA Ladder Mix(100~3000 bp)、50×TAE、瓊脂糖H、SanPrep柱式PCR產物純化試劑盒、異丙醇、1×TE 生工生物工程(上海)股份有限公司;無水乙醇 天津歐博凱化工有限公司。
BCM-1000A超凈工作臺 蘇凈安泰;霉菌培養箱、光照培養箱 上海博訊實業有限公司醫療設備廠;OLYMPUSDP73正置熒光顯微鏡 東京都涉谷區幡之谷生產;ZD-88全溫氣浴振蕩器 常州國宇儀器制造有限公司;PHS-3C PH計 上海儀電科學儀器股份有限公司;Verity 96well PCR儀、3730XL測序儀 美國ABI;FR-980A凝膠成像儀 上海復日科技有限公司;HC-2518R 冷凍離心機 安徽中科中佳科學儀器有限公司;TD5A-WS 臺式高速離心機 湖南湘儀實驗儀器開發有限公司;DYY-6C電泳儀 北京六一儀器廠;DYCP-32B電泳槽 北京六一儀器廠;WH-3 微型旋渦混合儀 上海滬西分析儀器廠有限公司;HS-800D數顯恒溫水浴鍋 太倉市科教器材廠;F619703-05 BP 系列精密單道可調移液器 加拿大BBI;SMA4000 UV-Vis Spectrophotometer Merinton;BCD-256KT冰箱 青島海爾股份有限公司;7.8×6×10牛津杯 上海三麝實業有限公司。
1.2 實驗方法
1.2.1 甜櫻桃流膠病原菌的鑒定 用無菌解剖刀刮取0.1~0.2 g已在PDA培養基28 ℃暗培養活化的病原真菌SXYTLJ01的菌絲于1.5 mL離心管中,按SK8259(真菌)試劑盒操作提取基因組DNA,置于-20 ℃冰箱保存備用。應用真菌核糖體基因轉錄間隔區通用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)、ITS4(5′-TCCTCCGCTTATTGATATGC-3′)和EF-1α-NS1(5′-GTAGTCATATGCTTGTCTC-3′)、EF-1α-NS6(5′-GCATCACAGACCTGTTATTGCCTC-3′)[18],以分離得到的病原菌基因組DNA為模板進行PCR擴增;引物均由生工生物工程(上海)有限公司合成。25 μL PCR反應體系:94 ℃預變性4 min,94 ℃ 45 s,55 ℃ 45 s,72 ℃ 1 min,共30個循環,72 ℃延伸10 min。擴增產物經1%瓊脂糖凝膠電泳,在全能成像系統上觀察并記錄結果。PCR產物送往生工生物工程(上海)有限公司測序。靶基因序列進行BLAST比對,MEGA6.0程序處理,NJ鄰接法[19]構建系統進化樹,分析病原菌的親緣關系,以確定其分類學地位。
1.2.2 菌絲結構的觀察 利用光學顯微鏡進行菌絲結構觀察:在回接有病原真菌SXYTLJ01的PDA上進行45°插片、28 ℃暗培養,培養3~7 d后用乳酸石炭酚棉藍進行染色,鏡檢,觀察該菌的菌絲形態結構[20]。
1.2.3 誘導產孢及孢子形態的觀察 采用碳氮源缺乏、菌落劃傷、近紫外線照射的方式誘導產孢[21]。每組處理至少重復3次。
碳氮源缺乏:將活化后長勢良好的菌株分別接種于不加碳源(葡萄糖)的馬鈴薯瓊脂培養基和不加氮源(蛋白胨)的馬鈴薯葡萄糖瓊脂培養基,28 ℃暗培養3~7 d,用乳酸石炭酚棉蘭染色,鏡檢,觀察產孢情況。
菌落劃傷:將活化后長勢良好的菌株接種于PDA培養基,28 ℃暗培養2~3 d,用無菌刀片沿培養基中部將菌落劃至見皿底,再培養1~3 d,在傷口邊緣取樣,染色液染色,鏡檢,觀察產孢情況。
近紫外線照射:將活化后長勢良好的菌株接種于PDA培養基,12 h近紫外線(18 W,直徑19 mm波長253.7 nm,距離培養基50 cm)照射后28 ℃暗培養2~3 d,鏡檢,觀察產孢情況。
1.2.4 最適生長條件 利用平板測定法[22]研究葡萄座腔菌的最適生長條件:將活化后長勢良好的菌株接種于PDA培養基中央,采用十字交叉法每天定時測定菌落直徑。分別將溫度、光照、pH、碳源、氮源這些因素作為單因素試驗條件,探究不同條件下菌株的生長狀況。每個處理至少重復3次,取其平均值。
溫度的影響:將菌株接種于PDA培養基中央,以15、20、25、28、30、35、40 ℃七個溫度作為試驗溫度梯度,暗培養,每24 h測量一次菌落直徑。
光照的影響:將菌株接種于PDA培養基中央,分別進行全光照、全黑暗、12 h光照/12 h黑暗交替三種培養處理,28 ℃培養,每24 h測量一次菌落直徑。
pH的影響:將菌株分別接種于pH為5.0、6.0、7.0、8.0、9.0、10.0的PDA培養基中央,28 ℃暗培養,每24 h測量一次菌落直徑。
碳源的影響:將菌株接種于不含碳源和碳源分別為蔗糖、麥芽糖、淀粉、乳糖、葡萄糖的PDA培養基中央,28 ℃暗培養,每24 h測量一次菌落直徑。
氮源的影響:將菌株接種于不含氮源和氮源分別為牛肉膏、蛋白胨、硫酸銨、硝酸鉀、硝酸鈉的PDA培養基中央,28 ℃暗培養,每24 h測量一次菌落直徑。
1.2.5 致病菌致死溫度區間的確定 將活化后長滿平板的菌落通過牛津杯取成直徑約0.8 cm的菌餅,接取菌餅于5 mL PDB(馬鈴薯-葡萄糖肉湯培養基)培養基中,分別將試管放在35、40、45、50、55、60 ℃的恒溫水浴鍋中水浴20 min,對照組不水浴[23]。水浴完成后將上述試管置于28 ℃搖速為180 r/min振蕩箱中暗培養72 h,觀察菌絲生長情況。
將上述震蕩培養的菌液進行平板回接驗證。具體步驟:長出菌絲球的,分別用鑷子鉗取大小相同的菌絲塊于PDA培養基;菌餅未生長的,分別用鑷子從菌餅上鉗取同等大小的菌塊于PDA培養基。28 ℃暗培養,連續觀察3~7 d,看是否生長。如果菌株在PDB培養液中經某一溫度區間水浴后振蕩培養不生長,且回接PDA培養基也不生長,則將此溫度區間確定為菌株SXYTLJ01的致死溫度區間。
1.3 數據處理
每組處理重復三次,取平均值;用Origin 8.0制圖;用SPSS 22.0統計軟件對差異顯著性進行分析,P<0.05為顯著水平,P<0.01為極顯著水平;用MEGA6.0軟件以NJ鄰接法構建系統發育樹。
2 結果與分析
2.1 基因序列分析
將ITS和EF-1α引物進行PCR擴增,菌株SXYTLJ01獲得了長度為500 bp左右的清晰條帶,電泳結果見圖1。將純化的PCR產物測序,分別獲得了511 bp和444 bp的序列。
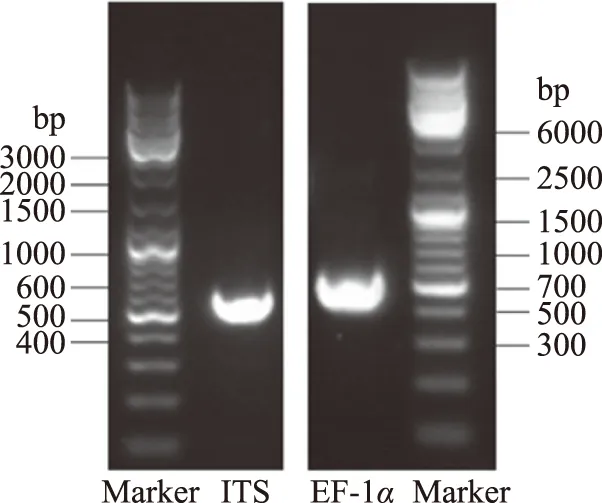
圖1 PCR擴增產物凝膠電泳結果Fig.1 Electrophoresis analysis of PCR amplified products
將測序所得的基因序列提交到NCBI的GenBank,應用BLAST進行同源序列比對。病原真菌的ITS序列和EF-1α序列與GenBank中Botryosphaeriadothidea分別有99.61%和99.77%的同源性。從GenBank數據庫中選取代表葡萄座腔菌屬不同種的ITS序列來構建系統發育樹,結果顯示:菌株SXYTLJ01與親緣關系較近的葡萄座腔菌(BotryosphaeriadothideaMK448265.1)聚于同一支(圖2)。結合形態學特征,最終將病原真菌SXYTLJ01鑒定為葡萄座腔菌(Botryosphaeria dothidea)。其ITS和EF-1α測序結果GenBank號分別為Accession No.SUB5881133H-ITS5_A01. ab1MN104148和Accession No.BankIt2240954 H-EF_A01.ab1BotryosphaeriaMN138458。
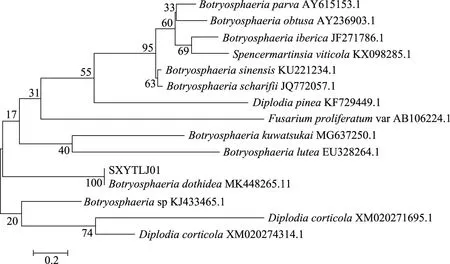
圖2 SXYTLJ01菌株基于rDNA-ITS序列的系統發育樹Fig.2 Phylogenetic tree based on the rDNA-ITS sequences of SXYTLJ01 pathogen
2.2 菌絲結構
菌絲經乳酸石炭酚棉蘭染色后的菌絲結構如圖3所示。
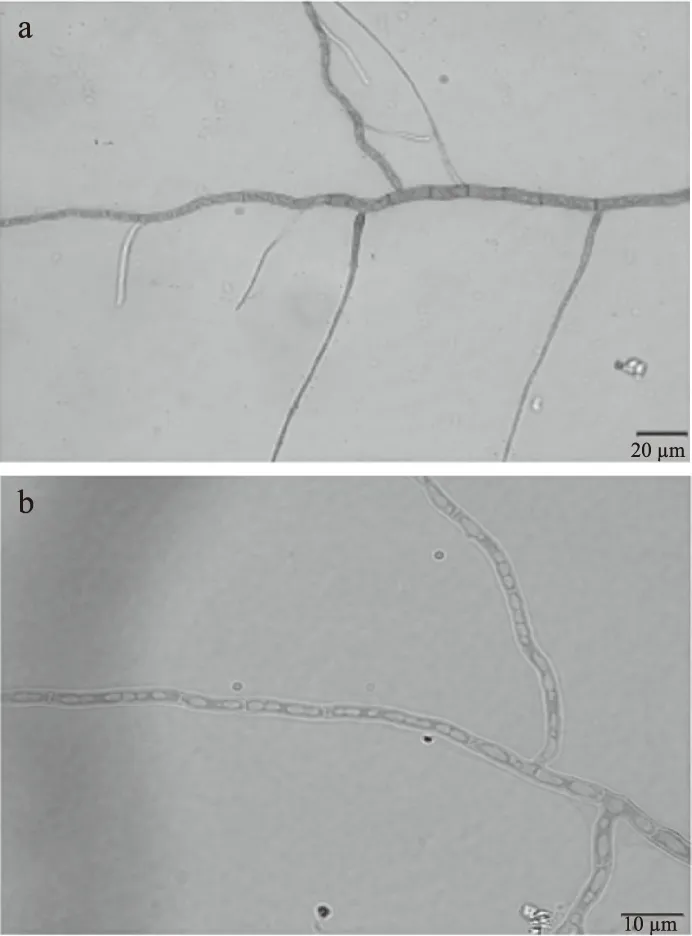
圖3 葡萄座腔菌菌絲結構圖Fig.3 Structure of hyphae of Botryosphaeria dothoidea注:圖3a為40倍光學顯微鏡下觀察所得;圖3b為100倍下光學顯微鏡下觀察所得。
40倍光學顯微鏡下,可以看到菌絲有明顯節狀結構,內含橫膈膜,同時菌絲有樹狀分支,有些菌絲聯結成網狀。100倍光學顯微鏡下,可以看出菌絲內部分隔明顯,將菌絲分隔成多個細胞,且菌絲呈分枝狀態。
2.3 不同脅迫條件下誘導產孢情況分析
由圖4可知:葡萄座腔菌孢子呈卵形或橢圓形,近紫外線照射、菌落劃傷、營養缺乏均能誘導其產孢。近紫外照射下誘導產孢,如圖4a,孢子在菌絲頂端分叉處大量聚集,大小不一。菌落劃傷誘導產孢,如圖4b,孢子呈卵形或橢圓形,有聚集現象。碳氮源缺乏誘導產孢,如圖4c和4d,孢子聚集度小,便于單獨觀察及抑制實驗的進行。
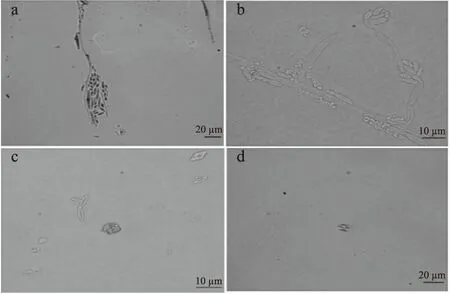
圖4 不同脅迫條件下誘導產孢Fig.4 Induction of sporulation under different stress conditions注:a為紫外誘導產孢圖;b為菌落劃傷誘導產孢圖;c為碳源缺乏誘導產孢圖;d為氮源缺乏誘導產孢圖。
2.4 致病菌最適生長條件
2.4.1 致病菌生長與溫度的關系 培養3 d后,菌落直徑與溫度的關系如圖5所示。
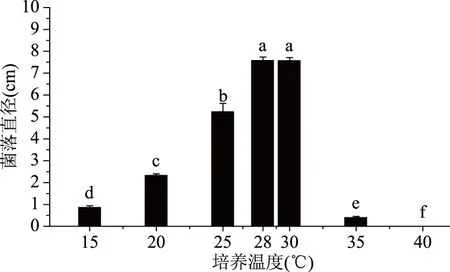
圖5 不同培養溫度對菌落生長的影響Fig.5 Effect of different culture temperatures on colony growth注:圖中不同字母代表菌落直徑差異的顯著性(P<0.05),圖6~圖9同。
由圖5可以看出,溫度是影響該致病菌生長的重要因素,在15~35 ℃下該菌均能生長,但菌落直徑存在顯著性差異;15~28 ℃之間,菌落直徑隨著溫度的升高顯著增加,致病菌的最佳生長溫度為28 ℃;28~35 ℃之間,菌絲直徑會隨著溫度的升高而持續下降;高于40 ℃時,致病菌不再生長。綜上,該致病菌的最適生長溫度為28 ℃。
2.4.2 致病菌生長與光照的關系 培養3 d后,菌落直徑與光照類型之間的關系如圖6所示。
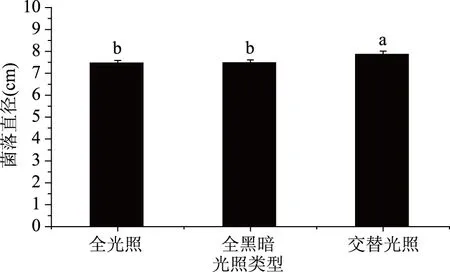
圖6 不同光照類型對菌落生長的影響Fig.6 Effect of different light types on colony growth
由圖6可知,無論何種光照類型,菌絲均能生長,但在不同光照條件下菌落直徑存在差異,全光照與全黑暗的菌落直徑差異不顯著,與交替光照的菌落直徑存在顯著性差異。由實驗結果可以看出,在培養到第3 d時,12 h光照/12 h黑暗交替培養比全光照、全黑暗的菌落直徑大。綜上,光照類型對該菌生長影響明顯,該致病菌的最適生長光照條件為12 h光照/12 h黑暗交替光照。
2.4.3 致病菌生長與pH的關系 培養3 d后,菌落直徑與pH的關系如圖7所示。
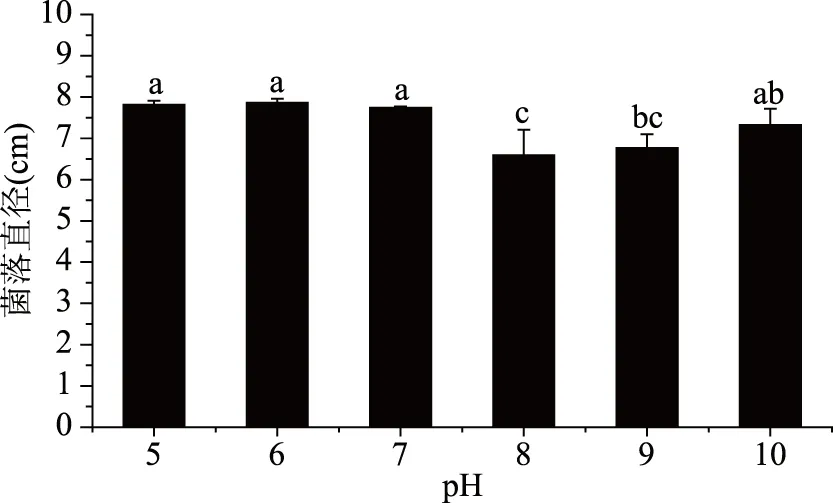
圖7 不同pH對菌落生長的影響Fig.7 Effect of different pH values on colony growth
由圖7可以看出,pH在5.0~10.0范圍內該菌均能生長,但在不同pH下菌落直徑存在差異,pH在5.0~7.0范圍內菌落直徑差異不顯著,在8.0~10.0范圍內菌落直徑存在顯著性差異;菌絲生長更適合偏酸性環境,pH為6.0時菌落直徑最大,說明在pH6.0下菌絲生長速率相對最快;當pH高于6.0時,菌絲還會生長,且隨著pH的升高,菌落直徑差異顯著,但都低于同培養天數下pH6.0時的菌落直徑。綜上,pH對該致病菌生長影響明顯,偏酸性的環境更有利于該菌的生長,以pH6.0最佳。
2.4.4 致病菌生長與碳源的關系 培養3 d后,菌落直徑與不同碳源的關系如圖8所示。
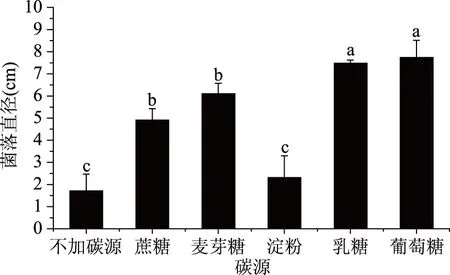
圖8 不同碳源對菌落生長的影響Fig.8 Effect of different carbon sources on colony growth
由圖8可以看出,在不加碳源以及以蔗糖、麥芽糖、淀粉、乳糖、葡萄糖為碳源時,該菌均能生長,但是各實驗組菌落直徑存在顯著差異。不加碳源的培養基菌落直徑增長最慢;以淀粉為碳源時,菌落直徑與不加碳源組差異不顯著、與其他組有顯著性差異;以蔗糖和麥芽糖為碳源時,兩組菌落直徑差異不顯著,菌落直徑相對較大,但與葡糖糖和乳糖為碳源時的菌落直徑有顯著差異,且比這兩組的菌落直徑要小;以葡萄糖和乳糖為碳源時,兩組菌落直徑差異不顯著,但葡萄糖的菌落直徑明顯要比乳糖大。綜上,碳源對該致病菌生長有重要影響,在以葡萄糖為碳源時該菌長勢最好,而以蔗糖、乳糖、麥芽糖這些雙糖作為碳源時生長稍差,以淀粉為碳源時生長最差。
2.4.5 菌絲生長與氮源的關系 相同的培養時間,菌落直徑與不同氮源的關系如圖9所示。
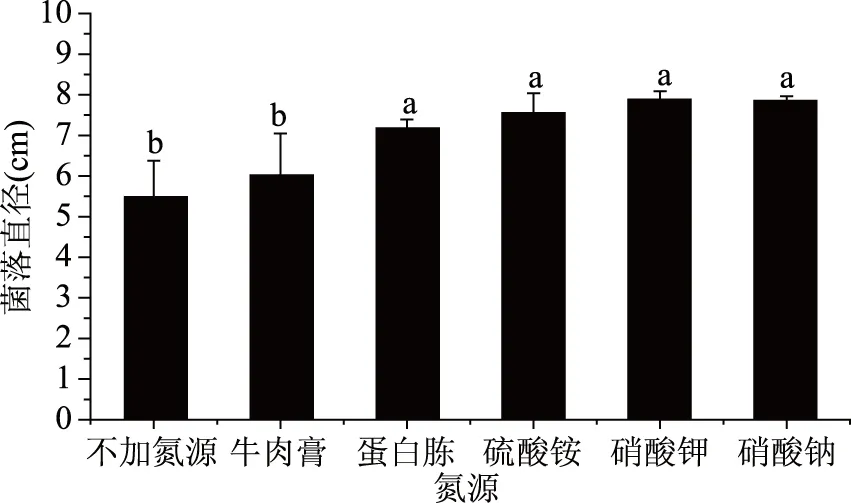
圖9 不同氮源對菌落生長的影響Fig.9 Effect of different nitrogen sources on colony growth
由圖9可以看出,不加氮源實驗組以及以牛肉膏、蛋白胨、硫酸銨、硝酸鈉、硝酸鉀為氮源的實驗組菌落均能生長,但是不加氮源組的菌落直徑明顯落后于加氮源組的,說明氮源為該菌生長的重要條件。從圖中可以看出,蛋白胨組、硫酸銨組、硝酸鉀組和硝酸鈉的菌絲直徑差異不顯著,但硫酸銨的菌落直徑>蛋白胨菌落菌落直徑,而以硫酸銨為氮源的菌落直徑明顯低于以硝酸鈉、硝酸鉀為氮源的實驗組。綜上,氮源為該致病菌生長的重要影響因素,且以硝酸鈉、硝酸鉀為氮源時,該菌生長最快,因此硝酸鈉、硝酸鉀類硝酸鹽類為該致病菌生長的最佳氮源。
2.5 致病菌致死溫度區間的確定
分別經35、40、45、50、55、60 ℃水浴20 min并振蕩培養72 h后,菌絲生長情況如圖10,回接后菌絲生長情況如圖11。

圖10 不同溫度對菌落生長的影響Fig.10 Effect of different temperature on colony growth
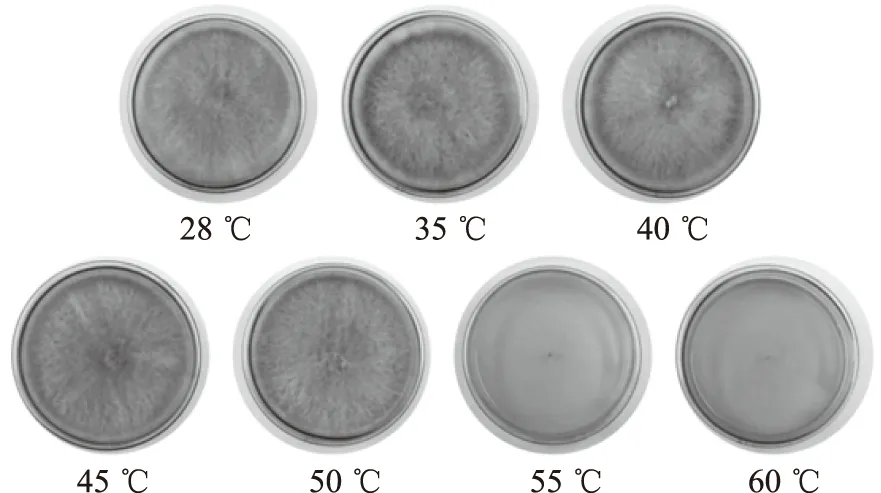
圖11 PDA回接實驗結果Fig.11 PDA verification experiment results
由圖10可知,分別經35、40、45、50、55、60 ℃水浴20 min后,28 ℃對照組、35、40、45、50 ℃實驗組均有菌絲生長且均形成了菌絲球,55、60 ℃的PDB培養液清澈,無菌絲生長。
取各試管中的培養液進行PDA回接實驗,結果如圖11。經28、35、40、45、50 ℃水浴20 min后回接在PDA培養基上,PDA培養基均有菌落生長,而經55、60 ℃水浴20 min后回接在PDA培養基上,PDA培養基無菌落生長。由以上實驗可知:該致病菌的致死溫度區間為50~55 ℃。
3 討論與結論
本實驗以山西榆次櫻桃園流膠樹體上分離純化得到的一株葡萄座腔菌為試驗菌株,對其生物學特性進行了試驗研究。預實驗階段將葡萄座腔菌分別接種于查氏培養基和PDA培養基,結果發現PDA培養基更適合此菌生長;試驗階段通過對菌絲結構的觀察,發現該致病菌菌絲有明顯節狀結構,內含橫膈膜,同時菌絲有樹狀分支,有些菌絲聯結成網狀。通過菌落劃傷、近紫外線照射及營養缺乏進行誘導產孢試驗發現,三種脅迫方式均能誘導其產生孢子,這一觀察與何靖柳等[21]在觀察紅陽獼猴桃葡萄座腔菌時的結果相反,卻與韓青梅等[24]在研究Botryosphaeriadothidea侵染蘋果果實的研究結果一致。
該葡萄座腔菌的孢子為卵形或橢圓形,大小不一,有聚集現象,與Phillips等[25]對葡萄座腔菌孢子形態的描述一致。
研究最適生長條件的實驗結果表明,溫度、光照、pH、碳源、氮源均為葡萄座腔菌生長的重要影響條件。葡萄座腔菌最適溫度為28 ℃,有力說明了5~9月份為發病較為嚴重階段的原因[16]。葡萄座腔菌最適光照條件為12 h光照/12 h黑暗交替。盧偉[26]對桂花葉斑病病原葡萄座腔菌(B.dothidea)進行生物學特性的研究試驗也認為病原菌生長的最適光照條件為交替光照,但羅光明等[27]認為光照對病原菌的生長無明顯影響。葡萄座腔菌最適生長pH為6.0,說明致病菌適生存于偏酸性環境。最適碳源為葡萄糖,最適氮源為硝酸鈉、硝酸鉀類硝酸鹽。該致病菌水浴處理20 min的致死溫度區間為50~55 ℃,這與黃靜等[28]對侵染草莓的落葉松葡萄座腔菌的致死溫度70 ℃(20 min)不一致;這說明不同植株上分離的致病菌雖然同屬葡萄座腔菌,但生物學特性有所差異。這些結果表明即使是同類植物,分離得到的致病菌特性也不相同,因此對葡萄座腔菌應該專株專治。
我國多樣的氣候及各種地理環境造就了豐富的葡萄座腔菌科真菌。本實驗所用的葡萄座腔菌來自山西榆次櫻桃園流膠樹體,葡萄座腔菌是引起多種樹體流膠的病原菌之一,通過本實驗對該致病菌生物學特性的研究結論,可以為其防治工作提供一定的理論基礎:調節櫻桃樹的栽種溫度,栽種土壤中性偏堿,避免硝酸類肥料,在櫻桃樹生長過程中要保護樹體,避免讓樹體出現傷口,減少紫外線照射,盡可能破壞致病菌生長的最佳條件,抑制葡萄座腔菌的生長,從而降低甜櫻桃流膠病的發病率,增加甜櫻桃產量。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
