荸薺表面拮抗細菌的分離鑒定及其表面抑菌活性
2020-06-19 02:23:50農建彪農進煥段振華
食品工業(yè)科技 2020年10期
張 巧,農建彪,農進煥,段振華,*
(1.賀州學院食品與生物工程學院,廣西賀州 542899; 2.廣西果蔬保鮮和深加工研究人才小高地,廣西賀州 542899)
荸薺(Eleocharisdulcis(Burm.f.)Trin. ex Hensch.)又名馬蹄、地栗、紅慈姑等,是一種果蔬兼用型的經濟作物,主要分布于廣西等地區(qū)。荸薺果肉鮮嫩質白,汁多爽脆,且營養(yǎng)豐富,有“地下雪梨”、“江南人參”的美譽[1-2]。但荸薺表面容易滋生腐敗微生物,鮮果荸薺儲藏期較短,嚴重影響荸薺的銷售及深加工。荸薺的貯藏保鮮方法多種多樣,傳統(tǒng)方法有地窖貯藏、堆藏法、細沙貯藏、溶液貯藏等[3],這些貯藏方法效果顯著,操作簡便且成本低廉,但方式粗放,不適合遠銷和高端銷售,已不能滿足現代荸薺產業(yè)的發(fā)展需求。以鮮切荸薺為主的初級加工產品,主要采用涂膜保鮮、熱處理、護色劑、氮氣保鮮等現代貯藏保鮮技術[4-7],成本較高,操作復雜,且對荸薺的安全品質有一定的影響。目前,對于大面積種植荸薺的農戶而言,仍采用傳統(tǒng)的泥土堆藏法。因此,安全、高效、無害的果蔬保鮮技術是荸薺保鮮的迫切需求。
生物防治是指利用微生物之間的拮抗作用來防治病原菌的一種方法。拮抗菌在果蔬采后病害防治方面發(fā)揮重要的作用,如地衣芽孢桿菌對芒果炭疽病有較好的防治,漢森酵母和梅奇酵母能有效抑制蘋果、油桃上霉菌的生長[8-10]。荸薺的腐爛與微生物活動息息相關,在貯藏過程中,其表面微生物種類與數量是在不斷變化的。在貯藏前期,由于荸薺水分含量高,利于細菌的生長。張巧等[11]從腐爛荸薺中分離出2株腐敗細菌木糖氧化無色桿菌和產氣腸桿菌。顏梅新等[12-13]研究荸薺腐爛病的主要病原真菌是棘孢木霉、尖孢鐮孢霉、奇異根串珠霉等16種。前期實驗發(fā)現,帶泥荸薺在貯藏過程中的質量損失率及腐爛率明顯低于洗凈荸薺[14]。帶泥荸薺表面泥土中的微生物及其在荸薺貯藏過程中發(fā)揮的作用,目前還未見相關研究。針對這一問題,本研究從荸薺表面泥土中分離出一些拮抗細菌,并探究其對荸薺表面微生物的影響,為荸薺貯藏過程中新型生物貯藏保鮮技術的開發(fā)提供一定的理論依據。
1 材料與方法
1.1 材料與儀器
荸薺 廣西賀州農戶;營養(yǎng)瓊脂培養(yǎng)基(NA)、平板技數瓊脂培養(yǎng)基(PCA)、孟加拉紅培養(yǎng)基 廣州環(huán)凱生物技術有限公司;2×Taq PCR MasterMix、細菌基因組DNA提取試劑盒 天根生化科技(北京)有限公司;一次性針頭式過濾器(0.22 μm,13 mm,水系) 天津市津騰實驗設備有限公司;大腸桿菌、金黃色葡萄球菌 廣東省微生物菌種保藏中心。
BSP-250型生化培養(yǎng)箱 上海博迅實業(yè)有限公司醫(yī)療設備廠;SQ510C型立式壓力蒸汽滅菌器 重慶雅瑪拓科技有限公司;722N型可分光光度計 上海儀電分析儀器有限公司;5424R型高速冷凍離心機 Eppendorf公司。
1.2 實驗方法
1.2.1 帶泥荸薺表面泥土中細菌的分離 取新鮮的帶泥荸薺,用一定量的無菌生理鹽水將泥土洗入三角瓶中,30 ℃、180 r/min搖勻2 h,靜置30 min,上層液體稀釋至合適的濃度。將15~20 mL無菌的NA瓊脂培養(yǎng)基倒入培養(yǎng)皿中,待其冷卻后凝固,吸取1 mL稀釋液至培養(yǎng)基表面,涂布均勻,37 ℃培養(yǎng)36 h,觀察細菌的生長情況。從上述培養(yǎng)皿中挑出菌落形態(tài)不一的單菌落50株,轉接至LB培養(yǎng)液中(酵母粉5 g/L,蛋白胨10 g/L,NaCl 10 g/L),37 ℃、200 r/min條件下培養(yǎng)24 h。取菌液劃線于NA瓊脂培養(yǎng)基中,獲得單菌落,如此重復3次,獲得純化菌株,并觀察菌落形態(tài),進行革蘭氏染色[15]。
1.2.2 拮抗細菌的篩選
1.2.2.1 發(fā)酵濾液的制備 將方法1.2.1分離的50株細菌分別接種至LB培養(yǎng)液中,37 ℃、200 r/min條件下培養(yǎng)24 h,獲得待試菌發(fā)酵液。將發(fā)酵液離心(4 ℃,10000 r/min,10 min),上清液在超凈工作臺中用0.22 μm的濾器過濾除菌,即為無菌的發(fā)酵濾液。
1.2.2.2 抑菌活性的測定 參照李亞輝等[16]的方法并稍作修改。以大腸桿菌作為革蘭氏陰性菌測試菌,金黃色葡萄球菌作為革蘭氏陽性菌測試菌,在3 mL已滅菌的LB液體培養(yǎng)基中,加入300 μL的上述無菌發(fā)酵濾液,對照組用等體積的無菌LB培養(yǎng)基代替,按照1%的接種量分別接種大腸桿菌菌液(OD600=1.5)和金黃色葡萄球菌菌液(OD600=1.0),搖勻后37 ℃、200 r/min條件下培養(yǎng)18 h,測定600 nm處的吸光度。細菌發(fā)酵濾液對大腸桿菌和金黃色葡萄球菌的抑制作用大小用抑菌率表示,按照如下公式進行計算。
抑菌率(%)=(A0-A1)/A0×100
式中:A0為對照組的吸光值;A1為實驗組的吸光值。
1.2.3 將分離到的純菌株在NA培養(yǎng)基中劃線,培養(yǎng)皿置于37 ℃的恒溫培養(yǎng)箱中倒置培養(yǎng),每隔12 h觀察培養(yǎng)皿中菌落的生長速度、顏色、大小、形狀、透明度、邊緣結構等。
1.2.4 生理生化特征 參照《常見細菌系統(tǒng)鑒定手冊》[17]對具有抑菌活性的細菌進行生理生化特征的鑒定。
1.2.5 分子生物學鑒定 將分離到的純菌株通過測定16S rDNA基因序列進行鑒定,具體參照潘曉倩等[18]的方法并稍作修改。純菌株接種于LB培養(yǎng)液中并于37 ℃培養(yǎng)24 h,按照細菌試劑盒上的方法提取DNA,以提取的DNA作為模板,采用通用引物27F(5′-AGAGTTTGATCMTGGCTCAG-3′)和1492R(5′-TACGGYTACCTTGTTACGACTT-3′)進行PCR擴增。
PCR反應體系:2×Taq PCR MasterMix 25 μL、ddH2O 21 μL、27F 1 μL、1492R 1 μL、DNA模板 2 μL。
PCR擴增程序:94 ℃預變性5 min,94 ℃變性0.5 min,56 ℃退火0.5 min,72 ℃延伸1.5 min,進行33個循環(huán)后72 ℃延伸10 min。
1%瓊脂糖凝膠電泳后,得到1500 bp左右的擴增產物委托青島英派生物技術有限公司進行測序。將測得的序列結果與NCBI GenBank中序列進行比對,通過MEGA6.06軟件構建系統(tǒng)發(fā)育樹。
1.2.6 細菌發(fā)酵濾液對荸薺表面微生物的影響 按1.2.2.1的方法制備無菌發(fā)酵濾液。將新鮮荸薺貯藏在20 ℃、80% RH的環(huán)境中5 d,取荸薺皮10 g,切碎后加入90 mL的無菌生理鹽水,30 ℃、180 r/min搖勻2 h,靜置30 min,取上清液稀釋一定倍數,吸取1 mL稀釋液至無菌培養(yǎng)皿中。將滅菌后的PCA、孟加拉紅瓊脂培養(yǎng)基50 mL冷卻至45~55 ℃,分別加入無菌發(fā)酵濾液10 mL,對照培養(yǎng)皿加入10 mL的無菌LB培養(yǎng)液,混勻后倒入培養(yǎng)皿中,充分搖勻,待其凝固后按照表1條件進行培養(yǎng),觀察培養(yǎng)皿中的菌落數。并將培養(yǎng)皿中的單菌落一一挑出,分別進行革蘭氏染色,顯微鏡觀察,確定培養(yǎng)皿中的球菌數、桿菌數、G+菌數、G-菌數,比較對照組和實驗組PCA培養(yǎng)皿中細菌類群。
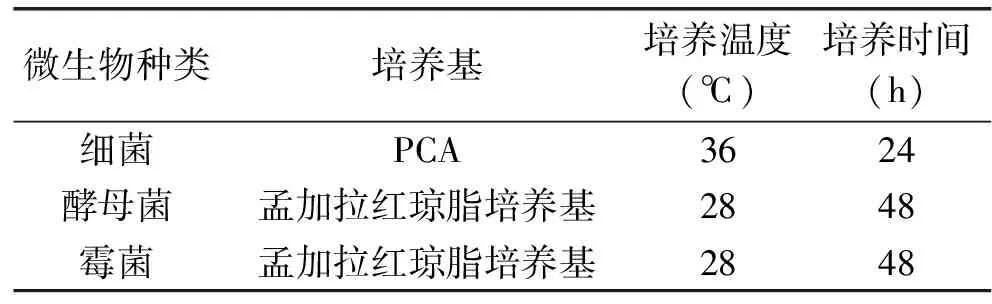
表1 微生物培養(yǎng)條件Table 1 Culture conditions of different microorganisms
1.3 數據處理
所有數據均為三次平行實驗的平均值,采用Excel 2007對實驗數據進行處理,Origin 8.0繪制趨勢曲線圖,SPSS 17.0進行方差分析和顯著性分析。
2 結果與分析
2.1 拮抗細菌的篩選
病原微生物是引起果蔬腐爛的一個重要因素,利用拮抗菌進行生物防治具有環(huán)保、高效、經濟的優(yōu)勢,在果蔬保鮮方面具有較廣的應用前景[19]。從帶泥荸薺表面泥土中分離出50株細菌,其中8株細菌(編號分別為WB12、WB20、WB22、WB38、WB39、WB41、WB46、WB47)的無菌濾液對大腸桿菌和金黃色葡萄球菌具有抑制作用,抑菌率見圖1所示。由圖1可知,WB22、WB39對大腸桿菌的抑菌率分別為23.36%、36.32%,對金黃色葡萄球菌的抑菌率分別為28.14%、20.37%。因此,菌株WB22、WB39對2株指示菌的抑制作用較強,具有拮抗菌的抗菌潛力。
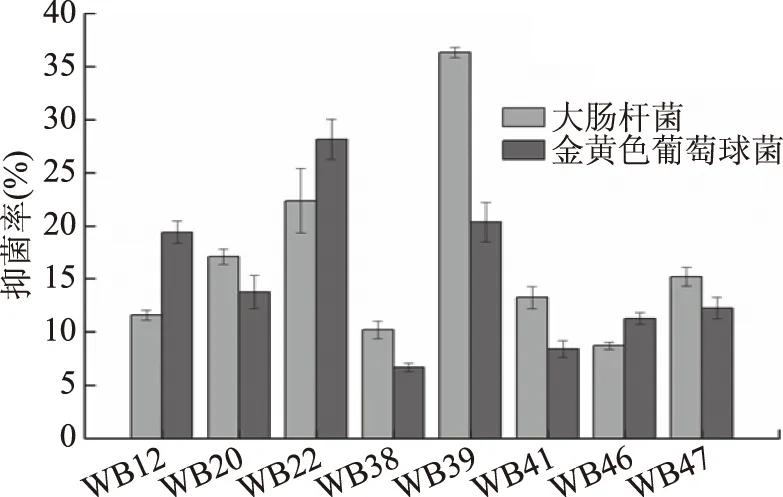
圖1 不同細菌對大腸桿菌和金黃色葡萄球菌的抑菌率Fig.1 Inhibitory rate of different bacteriaon Escherichia coli and Staphylococcus aureus
2.2 拮抗細菌的鑒定
2.2.1 菌落形態(tài)特征 通過觀察拮抗細菌WB22、WB39在固體培養(yǎng)基上的單菌落形態(tài),以及革蘭氏染色后的顯微鏡觀察,結果見表2。WB22為一株陽性芽孢桿菌,WB39為一株陰性無芽孢桿菌。

表2 菌落特征及菌體形態(tài)Table 2 The bacterial colony characteristics and morphology
2.2.2 生理生化特征 菌株WB22和WB39的生理生化特征結果由表3可知。WB22具有氧化酶、接觸酶活性,能水解淀粉、乳糖,V-P反應為陽性,不能產生H2S、吲哚。根據《常見細菌系統(tǒng)鑒定手冊》[17],結合菌落菌體形態(tài)及革蘭氏染色結果,WB22與芽孢桿菌的基本特性較為相似,初步判斷其為芽孢桿菌屬。而WB39具有氧化酶、接觸酶活性,但不能水解淀粉、乳糖等,不能進行V-P反應,能還原硝酸鹽,不產生H2S、吲哚,判定其為假單胞菌屬。

表3 生理生化特征Table 3 Characteristics of physiology and biochemistry
2.2.3 16S rDNA鑒定 將菌株WB22和WB39的PCR擴增產物采用瓊脂糖凝膠電泳進行純度分析,電泳結果見圖2。由圖2可知,菌株WB22和WB39僅在1500 bp附近顯示較強的電泳條帶,兩株拮抗細菌的16S rDNA均被成功擴增。
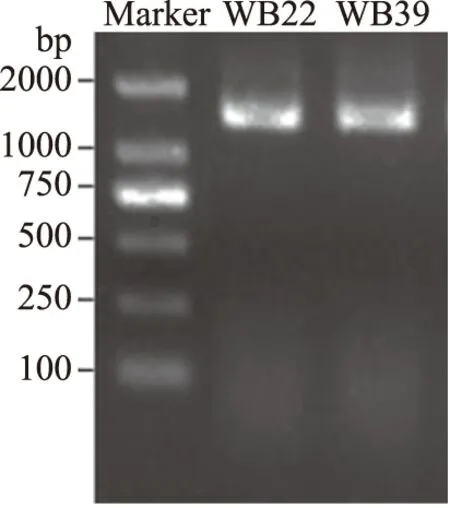
圖2 WB22和WB39的16S rDNA PCR擴增產物電泳圖Fig.2 Electrophoresis patterns of PCR amplificationproducts of WB22 and WB39 based on 16S rDNA
將菌株WB22和WB39的16S rDNA序列與相似度高的已知菌株序列進行比對,構建的系統(tǒng)發(fā)育樹見圖3所示。由圖3可知,菌株WB22與Bacillusamyloliquefaciens的同源性最高,WB39與Pseudomonasgeniculata的同源性最高,序列相似度均大于99%。因此,WB22被鑒定為解淀粉芽孢桿菌,WB39被鑒定為彎曲假單胞菌。據報道,解淀粉芽孢桿菌是一種具有廣譜抑菌活性的細菌,通過形成一些抑菌蛋白、脂肽類物質、聚酮化合物等抑菌物質,能有效地抑制細菌、真菌等腐敗微生物。因其具有抑菌譜廣、安全無污染、不產生抗藥性等優(yōu)點,在果蔬采后保鮮方面有著較好的應用前景[20]。蔣凱麗等[21]從櫻桃番茄果熟果中分離到一株解淀粉芽孢桿菌,對番茄采后常見病原菌抑制效果較好。假單胞菌能夠產生較多的活性物質,比如細菌素、抗生素、嗜鐵螯合物等,在生物防治上已有較多應用[22]。Kumar等[23]分離出一株能產生抗生素的假單胞菌,對多種植物病原菌起著抑制作用;張美琴等[24]從土壤中分離的一株具有開發(fā)潛力的假單胞菌,其對馬鈴薯晚疫疫霉菌的拮抗率高達89%。
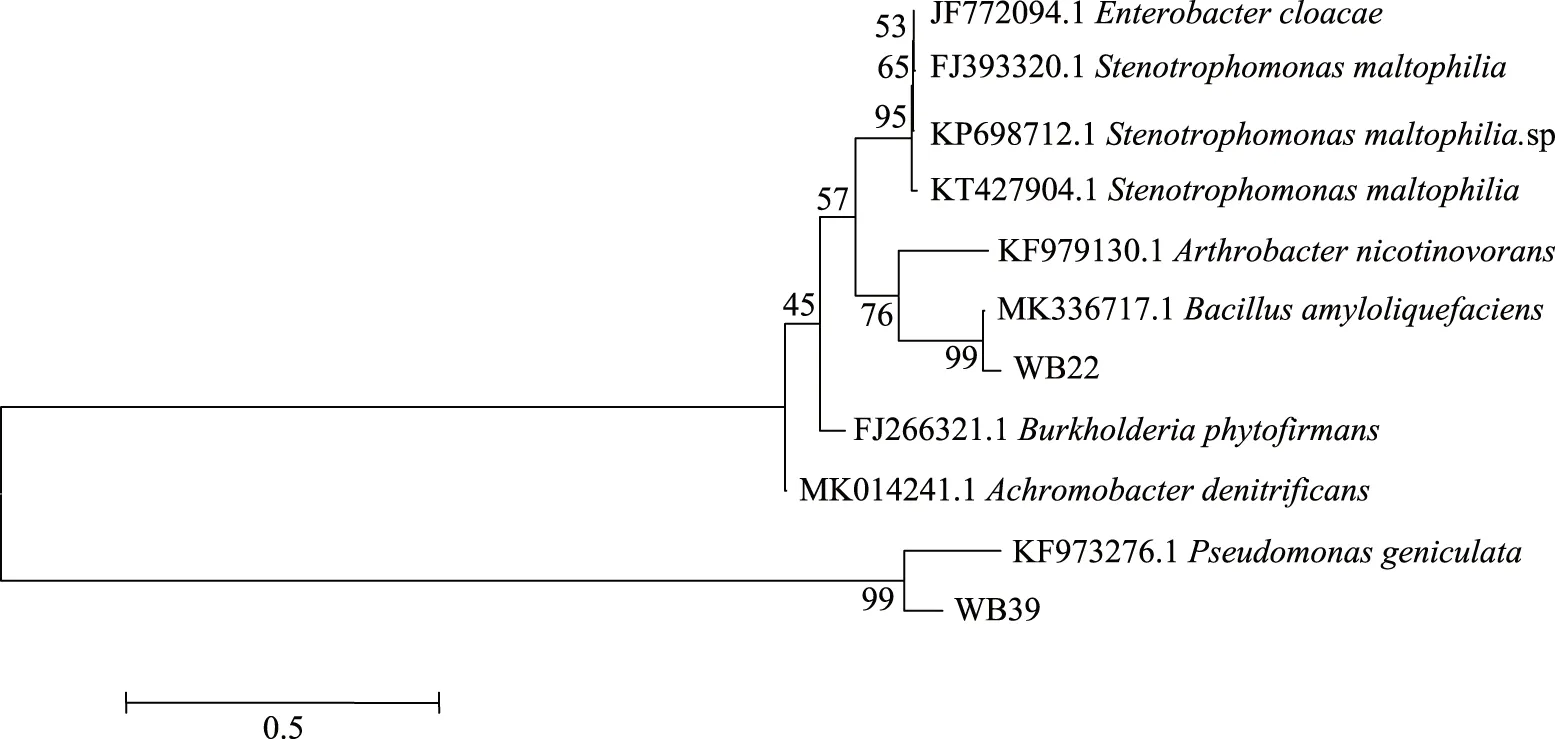
圖3 WB22和WB39的16S rDNA系統(tǒng)發(fā)育樹Fig.3 Phylogenic trees of WB22 and WB39 based on their 16S rDNA
2.3 拮抗細菌對荸薺表面微生物的影響
在培養(yǎng)基中分別添加一定量的B.amyloliquefaciens和P.geniculata發(fā)酵濾液,培養(yǎng)皿中生長的菌落數越少,說明抑菌效果越好。由圖4可知,對照組培養(yǎng)皿中的細菌數為4.88 lg(CFU/mL),分別添加B.amyloliquefaciens和P.geniculata發(fā)酵濾液的培養(yǎng)皿,細菌數分別為4.03和3.89 lg(CFU/mL),說明2株拮抗細菌的發(fā)酵濾液對荸薺表面細菌有抑制作用,且P.geniculata的抑制作用強于B.amyloliquefaciens;對照組培養(yǎng)皿中的酵母菌數為4.31 lg(CFU/mL),分別添加B.amyloliquefaciens和P.geniculata發(fā)酵濾液的培養(yǎng)皿,酵母菌數分別為3.95和4.11 lg(CFU/mL),荸薺表面的酵母菌受到一定的抑制作用,B.amyloliquefaciens的抑制作用強于P.geniculata;對照組培養(yǎng)皿中的霉菌數為3.78 lg(CFU/mL),分別添加B.amyloliquefaciens和P.geniculata發(fā)酵濾液的培養(yǎng)皿,霉菌數分別為3.26 lg和3.17 lg(CFU/mL),荸薺表面霉菌的生長被抑制,且P.geniculata的抑制作用強于B.amyloliquefaciens。經數據統(tǒng)計分析,B.amyloliquefaciens對荸薺表面細菌、酵母菌、霉菌的抑菌率分別為17.42%、9.05%和13.74%;P.geniculata對荸薺表面細菌、酵母菌、霉菌的抑菌率分別為20.29%、4.64%和16.14%。2株拮抗細菌發(fā)酵濾液對荸薺表面細菌、霉菌具有顯著的抑制作用(P<0.05),且對細菌的抑制作用最強,對酵母菌的抑制作用不明顯(P>0.05)。
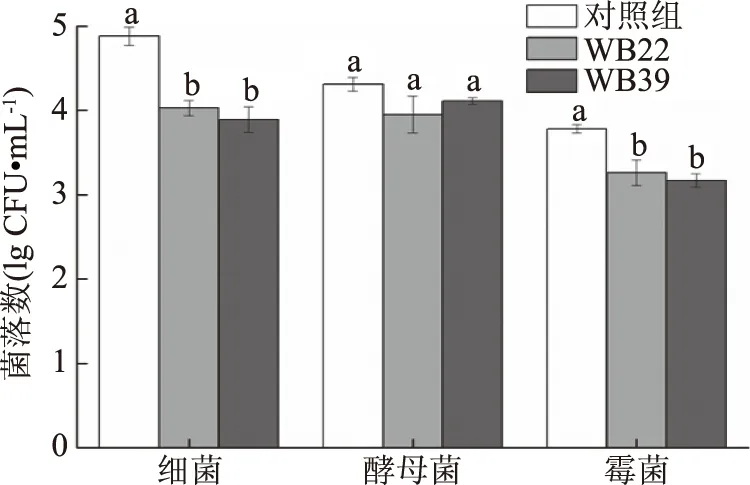
圖4 WB22和WB39對荸薺表面微生物數的影響Fig.4 Effects of WB22 and WB39 on the microbial counts ofEleocharis dulcis(Burm.f.)Trin. ex Hensch. surface 注:同類微生物柱形圖上的不同字母表示差異顯著(P<0.05)。
前期研究也表明,造成荸薺腐爛的主要病原微生物是霉菌和細菌[11-13]。因此,2株拮抗菌在荸薺采后的生物防治方面有較好的應用價值。
2株拮抗細菌對荸薺表面細菌不同類群(球菌、桿菌數,G+菌、G-菌)的影響結果見表4。由表4可知,對照組的球菌數和桿菌數分別為1.56和3.32 lg(CFU/mL),分別占細菌總數的31.97%、68.03%,G+菌和G-菌數分別為2.01和2.87 lg(CFU/mL),分別占比41.19%、58.81%。與對照組相比,添加B.amyloliquefaciens發(fā)酵濾液的培養(yǎng)皿中,球菌數和桿菌數分別為0.82和3.21 lg(CFU/mL),占比分別為20.35%和79.65%,G+菌和G-菌分別為1.32和2.71 lg(CFU/mL),占比分別為32.75%和67.25%。B.amyloliquefaciens發(fā)酵濾液使得培養(yǎng)皿中的球菌和G+數下降較多,桿菌和G-下降較少,說明其主要抑制的細菌是球菌和G+菌。添加P.geniculata發(fā)酵濾液的培養(yǎng)皿中,球菌數和桿菌數分別為1.31和2.58 lg(CFU/mL),占比分別為33.68%和66.32%,G+菌和G-菌分別為1.43和2.46 lg(CFU/mL),占比分別為36.76%、63.24%。因此,添加P.geniculata發(fā)酵濾液的培養(yǎng)皿中,球菌、桿菌、G+菌、G-菌數明顯下降,但占比變化不大,說明P.geniculata發(fā)酵濾液抑菌范圍較廣,對桿菌和球菌、G+菌和的抑制作用均較強。拮抗菌對不同種類的細菌表現出不一樣的抑菌效果,這在其他研究也有發(fā)現,郝彥利[25]從海水中分離一株枯草芽孢桿菌,其對G+菌的抑菌效果同樣強于G-菌。陳幫丹等[26]研究一株糞腸堿桿菌的發(fā)酵液對部分臨床G-菌有抑制作用,但是不能抑制臨床G+菌。

表4 WB22和WB39發(fā)酵濾液抑制細菌的類別分析Table 4 Species of bacteria inhibited by the fermented filtrates of WB22 and WB39
3 結論
從帶泥荸薺表面泥土中分離篩選得到2株拮抗細菌WB22、WB39,經菌體形態(tài)、生理生化特征及16S rDNA基因序列分析,確定WB22為解淀粉芽孢桿菌(B.amyloliquefaciens),WB39為彎曲假單胞菌(P.geniculata)。目前對荸薺的生物防治研究還較少,本研究分離到的WB22、WB39對大腸桿菌和金黃色葡萄球菌具有較強的抑制作用,作為拮抗菌在荸薺的生物保鮮方面具有一定的開發(fā)前景。
2株細菌發(fā)酵濾液對荸薺表面微生物具有一定的抑制作用,其中B.amyloliquefaciens發(fā)酵濾液對荸薺表面細菌、酵母菌、霉菌的抑菌率分別為17.42%、9.05%和13.74%,P.geniculata對荸薺表面細菌、酵母菌、霉菌的抑菌率分別為20.29%、4.64%和16.14%,說明B.amyloliquefaciens與P.geniculata在生長過程中,形成了某些抑菌物質,從而抑制著荸薺表面微生物的生長。且B.amyloliquefaciens主要抑制的細菌是球菌和G+菌,而P.geniculata發(fā)酵濾液的抑菌范圍較廣,對桿菌和球菌、G+菌和G-菌均具有較好的抑制作用。本研究為2株拮抗菌的實際應用及抑菌機理的探究提供了理論參考。
