載脂蛋白C3轉基因小鼠應用于嚴重高三酰甘油血癥急性胰腺炎的探索
2020-06-21 03:36:16徐有青崔純瑩趙志剛
首都醫科大學學報 2020年3期
柳 鑫 徐有青 崔純瑩 趙志剛*
(1. 首都醫科大學附屬北京天壇醫院藥學部,北京 100070; 2. 首都醫科大學附屬北京天壇醫院消化內科,北京 100070;3.首都醫科大學藥學院藥劑學系,北京 100069)
急性胰腺炎是由于胰蛋白酶原過度激活,胰蛋白酶生成增加,引起胰腺自我消化的一種急性炎性反應[1]。流行病學調查研究[2]顯示,繼膽道疾病與乙醇中毒因素外,嚴重高三酰甘油(triglyceride,TG)血癥已成為急性胰腺炎的第三發病因素。嚴重高三酰甘油血癥急性胰腺炎的發病機制尚不明確,主要原因在于缺少合適的嚴重高三酰甘油血癥動物模型[3-5]。
通過高脂飲食誘導動物,血漿三酰甘油濃度只能增加2~3倍,采用腹腔注射TritonWR1339抑制脂蛋白脂酶(lipoprotein lipase,LPL)活性,可使血漿三酰甘油濃度增加5~10倍,但是急性效應較短暫,且對動物應激狀態影響很大。早期有使用LPL基因敲除小鼠探討嚴重高三酰甘油血癥-急性胰腺炎易感性的報道[4],但由于LPL基因敲除小鼠的新生期致死性,救治存活率極低,該模型應用于嚴重高三酰甘油血癥-急性胰腺炎的病理機制研究非常困難,經濟成本也很高。為了探索嚴重高三酰甘油血癥-急性胰腺炎的病理機制,需要使用易于繁殖的嚴重高三酰甘油血癥小鼠模型。
載脂蛋白C3(apolipoprotein C3,ApoC3)是由肝臟和小腸合成的一種載脂蛋白,ApoC3除了能夠抑制LPL水解三酰甘油,還能抑制肝臟攝取乳糜微粒、極低密度脂蛋白(very low density lipoprotein,VLDL)及其殘體脂蛋白,引起血漿三酰甘油濃度升高[1]。在三酰甘油代謝中,過表達ApoC3基因,能夠引起血漿三酰甘油濃度極度升高。ApoC3基因發生突變后,可導致血漿三酰甘油濃度降低,與動脈粥樣硬化心血管風險下降有直接相關性[6-7]。
為了驗證過表達人ApoC3基因的嚴重高三酰甘油血癥小鼠是否也發生急性胰腺炎,使用人ApoC3轉基因(ApoC3 transgenic,ApoC3tg)的小鼠模型,使用雨蛙素(caerulein)誘導急性胰腺炎后,與野生型小鼠比較,ApoC3tg小鼠表現出嚴重高三酰甘油血癥,胰腺病理損傷加重,使用抗氧化劑叔丁基-特丁基對苯二酚(tertiary-butylhydroquinone, tBHQ)預防性給藥,并不能使胰腺病理損傷得到改善。
1 材料與方法
1.1 試劑
Caerulein購自美國Sigma公司,tBHQ由北京大學藥學院余四旺老師贈送,生物化學檢測試劑盒(淀粉酶活性試劑盒、三酰甘油試劑盒、總膽固醇試劑盒與血糖試劑盒)均購自北京中生北控生物科技公司。
1.2 小鼠分組與基因型鑒定
ApoC3tg小鼠由美國Jackson實驗室提供,采用同窩對照野生型小鼠。所用小鼠均為C57BL/6J背景下8周齡雄性小鼠(LA2010-059),實驗分為兩組:野生型(wild type,WT)、ApoC3tg小鼠,每組6只,提取鼠尾DNA,采用PCR鑒定基因型。所采用的引物詳見表1。

表1 基因型鑒定的引物序列
ApoC3tg: apolipoprotein C3 transgenic.
1.3 血脂測定
自小鼠眶靜脈采血,取上層血漿,利用生物化學檢測試劑盒測定野生型小鼠與ApoC3tg小鼠的血漿TG、總膽固醇(total cholesterol,TC)和血糖(glucose,Glu)。
1.4 急性胰腺炎誘導
每組小鼠自由飲水,禁食12 h,采用雨蛙素(caerulein)誘導胰腺炎,按50 μg/kg腹腔注射給藥,1次/h,連續注射7次,第一次腹腔注射24 h 后處死小鼠,心臟灌流,提取胰腺組織,用于病理學分析。
1.5 血漿淀粉酶活性檢測
肝素抗凝后眶靜脈采血,4 °C,12 000 r/min,離心60 min,吸取3 μL下層澄清液體,加入96孔板中,吸取100 μL淀粉酶工作液,37 °C恒溫孵育3 min,按多點法設置酶標儀參數,在605 nm波長處讀取1、2、3 min 末的A值,血漿淀粉酶活性(U/L)=[A(3 min)-A(1 min)]/2×3 245×稀釋倍數,當[A(3 min)-A(1 min)]/2>0.3時,用ddH2O稀釋樣品后測定。
1.6 胰腺組織染色與病理評估
胰腺組織在25%~100%(體積分數)乙醇中,分別浸泡10 min,脫水,采用二甲苯浸泡至組織透明,加入熔化好的石蠟,60 ℃下保溫30 min,石蠟切片厚度為6 μm,采用二甲苯脫蠟,無水乙醇洗脫二甲苯,分別使用蘇木精與伊紅染色1 min,二甲苯洗脫處理后,中性樹膠封片,采集圖像,進行胰腺組織病理學評分,炎性細胞數量取10個,400×放大視野的平均值,實質細胞壞死與空泡化水平以腺泡細胞壞死和空泡百分比來衡量(表2)。
1.7 抗氧化劑tBHQ對ApoC3tg小鼠淀粉酶活性與胰腺病理損傷的影響
為了觀察氧化應激在嚴重高三酰甘油血癥-急性胰腺炎中的作用,ApoC3tg小鼠灌胃給予不同劑量抗氧化藥物tBHQ,對照組給予空白溶劑聚乙二醇,7 d后采用上述方法誘導急性胰腺炎。

表2 胰腺組織病理評分標準

圖2 ApoC3tg小鼠血漿脂質濃度升高
1.8 統計學方法
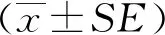
2 結果
2.1 ApoC3tg小鼠血脂升高
ApoC3tg小鼠基因鑒定如圖1所示,與野生小鼠相比,ApoC3tg小鼠體質量與血糖濃度差異無統計學意義(圖2A、B),血漿三酰甘油明顯升高(圖2C),總膽固醇含量伴隨三酰甘油升高而升高(圖2D)。

圖1 ApoC3基因型小鼠在60 bp處有明亮條帶
2.2 ApoC3tg小鼠血漿淀粉酶活性在9 h與12 h時明顯升高
ApoC3tg小鼠與野生型小鼠的血漿淀粉酶活性,隨時間發生顯著變化(F=28.576,P=0.001),在誘導胰腺炎后9 h與12 h時明顯升高,兩組之間的血漿淀粉酶活性差異沒有統計學意義(F=0.001,P=0.979),詳見圖3。

圖3 不同時間點下的血漿淀粉酶活性變化
2.3 ApoC3tg小鼠胰腺病理損傷加重
與野生型小鼠相比,ApoC3tg小鼠胰腺組織水腫加重,小葉間與內部間隙增寬,結構紊亂,小葉間有大量炎性細胞浸潤(圖4),分別對胰腺組織水腫與炎細胞浸潤指標進行病理學評分(表3),ApoC3tg小鼠胰腺組織的病理評分顯著高于野生小鼠,表明ApoC3tg小鼠的胰腺炎性反應加重,對雨蛙素誘導的急性胰腺炎更加敏感。

圖4 ApoC3轉基因小鼠胰腺病理損傷顯著加重
2.4 抗氧化劑tBHQ沒有改變ApoC3tg小鼠的血漿淀粉酶活性與胰腺病理
隨著時間變化,血漿淀粉酶活性發生明顯改變(F=45.120,P=0.001),分組之間差異沒有統計學意義(F=2.050,P=0.185)(圖5)。
胰腺組織病理切片HE染色結果(圖6)顯示,與對照組小鼠相比,低、中或高劑量的tBHQ均未見胰腺病理損傷得到改善,對胰腺組織水腫與炎細胞浸潤指標進行病理評分(表4),tBHQ 50、 100、 200 mg/kg給藥組ApoC3tg小鼠的胰腺組織水腫沒有減輕,小葉間與內部間隙沒有縮小,小葉間炎性細胞浸潤沒有減輕。
表3 ApoC3tg小鼠胰腺組織病理學評分


GroupWTApoC3tgtPEdema2.8±0.845.4±1.52*2.7460.025Inflammatory infiltration2.6±0.554.6±1.14*4.1740.033
*P<0.05vsWT;WT: wild type;ApoC3tg: apolipoprotein C3 transgenic;n=6.

圖5 不同劑量抗氧化藥物tBHQ對ApoC3轉基因小鼠血漿淀粉酶活性的影響

圖6 不同劑量抗氧化藥物tBHQ沒有改善ApoC3tg小鼠的胰腺損傷

表4 tBHQ用藥后ApoC3tg小鼠胰腺組織病理評分
n=6;tBHQ: terbiary-butylhydroquinone;ApoC3tg: apolipoprotein C3 transgenic.
3 討論
本研究顯示,ApoC3轉基因小鼠血漿三酰甘油濃度明顯升高,使用雨蛙素誘導急性胰腺炎后,血漿淀粉酶活性在9 h與12 h均升高,胰腺病理損傷加重,具體表現為水腫加重,小葉間與內部間隙增寬,結構紊亂,小葉間有大量炎性細胞浸潤,給予抗氧化劑tBHQ后,未見淀粉酶活性改變與胰腺病理損傷的改善。
流行病學調查研究[8-9]顯示,我國高三酰甘油血癥患病率約為高膽固醇血癥的4倍,高三酰甘油血癥除了作為心血管疾病的獨立危險因素,其誘發急性胰腺炎的危險性越來越受到臨床關注。早期的動物實驗也顯示自發性嚴重高三酰甘油血癥動物LPL基因突變的水貂,可出現自發性急性胰腺炎,但其僅存在于挪威[10-11]。LPL基因突變的小鼠雖然可以通過救治進行高三酰甘油血癥急性胰腺炎研究,但國外報道[12]通過基因治療救治存活率很低。因此,以上研究采用的動物模型有很大的局限性。
高三酰甘油血癥急性胰腺炎有典型的急性發作或反復急性發作特點,部分患者出現反復腹痛癥狀,與非高三酰甘油血癥胰腺炎相比,高三酰甘油血癥急性胰腺炎患者早期容易出現血鈣降低、血鈉降低、低蛋白血癥以及結腸中斷征等主要臨床表征[13]。三酰甘油受進食狀態影響,在中等程度(177~885 mg/dL,2~10 mmol/L)的高三酰甘油血癥狀態下也會引起急性胰腺炎發作[14]。高三酰甘油血癥引起急性胰腺炎的發生和發展機制目前尚不明確,推測其可能有以下三種機制[15-16]。 ① 脂肪酸學說:三酰甘油的分解產物游離脂肪酸(free fatty acids,FFA)產生過多,在胰腺內不斷聚集,使胰腺腺泡細胞和較小血管受損,導致急性胰腺炎的發生與加重[17],而FFA誘導胰腺腺泡細胞受損的機制可能有以下幾種情況:FFA增多引起胰腺腺泡細胞內酸堿度下降,胰蛋白酶原激活,進而導致胰腺腺泡細胞自身消化[18]。② 胰腺微循環障礙:胰腺微循環障礙在急性胰腺炎的發生與病程發展中起重要作用,在急性胰腺炎的發病過程中,微循環障礙會影響胰腺的血流灌注,從而加重急性胰腺炎[19],高三酰甘油血癥患者血漿中,大顆粒的乳糜微粒會使胰腺內毛細血管發生栓塞,引起微循環障礙,從而導致胰腺缺血與壞死[20]。③ 氧化應激學說:在胰腺炎發病的初始階段,氧化應激狀態下產生氧自由基,一方面直接損傷DNA,引起蛋白質內巰基氧化直接損傷胰腺細胞[21],另一方面氧自由基的過氧化物與其他炎性介質協同作用,加重炎性反應,降低細胞膜穩定性,使胰腺腺泡細胞溶酶體酶釋放和消化酶活化[22]。
為了探討臨床上常見的嚴重高三酰甘油血癥與急性胰腺炎的關系,筆者采用ApoC3轉基因的嚴重高三酰甘油血癥小鼠,結果表明,與LPL基因敲除小鼠相似,嚴重高三酰甘油血癥-急性胰腺炎的ApoC3tg小鼠胰腺損傷明顯,表明嚴重高三酰甘油血癥下急性胰腺炎易感性增高。腹腔注射雨蛙素誘導急性胰腺炎后,ApoC3tg小鼠與野生小鼠的血漿淀粉酶活性均在不同時間點(9 h與12 h)明顯升高,但兩種小鼠的血漿淀粉酶活性差異無統計學意義。在臨床中也常見報道[23-24],和其他急性胰腺炎患者不同,嚴重高三酰甘油血癥-急性胰腺炎患者的血漿淀粉酶活性并不是一個靈敏的生物標志物,對于臨床診療意義不大。本結果顯示,在嚴重高三酰甘油血癥動物模型上,血漿淀粉酶同樣不是反映胰腺病理嚴重程度的靈敏的生物標識物。
早期有學者使用LPL基因敲除小鼠,對嚴重高三酰甘油血癥與急性胰腺炎的關系進行了初步探索,發現LPL基因敲除小鼠體內乳糜微粒與游離脂肪酸含量明顯升高,導致胰腺細胞受損,同時胰脂肪酶水解乳糜微粒,抑制鈣離子進入腺泡細胞內,促進嚴重高三酰甘油血癥下胰腺病理損傷[25]。LPL敲除小鼠的血漿丙二醛明顯高于野生型小鼠,而分離出的乳糜微粒氧化程度和年齡成正比,說明這一動物氧化應激易感性增高,并且因為氧化損傷導致自發性動脈硬化的發生[26]。這些數據提示,LPL基因敲除小鼠對胰腺炎易感性增高和發生自發性胰腺炎有可能與氧化應激損傷有關。
抗氧化劑tBHQ作為核轉錄因子Nrf2的激動劑,能通過上調血紅素加氧酶-1、超氧化物歧化酶等氧化應激因子,對多種炎性反應與氧化應激有改善作用[27-28]。因此,使用tBHQ對ApoC3tg小鼠進行實驗性預防給藥,未發現其對嚴重高三酰甘油血癥-急性胰腺炎有明顯改善作用,除了tBHQ,筆者也曾使用其他抗氧化劑(二叔丁基對甲酚與普羅布考等)對嚴重高三酰甘油血癥-急性胰腺炎進行干預,均未見胰腺病理損傷的改善(文章尚未發表)。由此推測,氧化應激可能并非ApoC3tg小鼠在嚴重高三酰甘油血癥下急性胰腺炎易感性增高的誘導機制。
本研究使用過表達人ApoC3基因的嚴重高三酰甘油血癥小鼠,誘導急性胰腺炎后,符合臨床嚴重高三酰甘油血癥-急性胰腺炎患者的特點,可以作為理想的研究嚴重高三酰甘油血癥動物模型探討急性胰腺炎發病機制。使用基因修飾的高三酰甘油血癥動物模型,探討嚴重高三酰甘油血癥狀態下急性胰腺炎發生與發展機制,對嚴重高三酰甘油血癥-急性胰腺炎的診斷與治療,以及藥物的有效性評估,具有一定的理論意義與實用價值。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
現代臨床醫學(2022年4期)2022-09-29 07:38:00
昆明醫科大學學報(2021年4期)2021-07-23 01:21:50
學苑創造·A版(2020年9期)2020-10-13 09:41:02
云南醫藥(2019年3期)2019-07-25 07:25:14
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
海南醫學(2016年8期)2016-06-08 05:43:00
醫學研究雜志(2015年9期)2015-07-01 17:28:15
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
鄭州大學學報(理學版)(2014年3期)2014-03-01 04:21:05
