脂聯素及其受體在哺乳動物中的研究進展與展望
2020-07-10 06:42:18張德榮馬曉霞李羽翡趙永清霍生東馬忠仁柏家林
生物技術通報 2020年6期
關鍵詞:研究
張德榮 馬曉霞 李羽翡 趙永清 霍生東 馬忠仁 柏家林
(1. 西北民族大學生物醫學研究中心 生物工程與技術國家民委重點實驗室,蘭州 730030;2. 甘肅省食品檢驗研究院,蘭州 730030;3. 西北民族大學生命科學與工程學院,蘭州 730030)
作為唯一與肥胖癥狀呈負相關的脂肪細胞因子——脂聯素(Adiponectin,APN)[1-2],自1995年美國懷特黑德生物醫學研究所學者Philipp E·Scherer發現至今的24年來,研究者們對脂聯素及其受體的相關研究不斷深入,脂聯素從單一的“能量儲存”功能,到參與機體葡糖代謝、維持能量平衡,具有抗氧化、抗消炎、抗動脈粥硬化和胰島素拮抗以及心血管疾病等方面的調節作用,是發揮多種生物功效的活性物質[3]。隨著人們對健康生活的不斷追求,如何提高動物機體的生長性能,如何有效降低體內脂肪沉積,如何預防和減少疾病的發生,如何加強臨床診斷及新藥物治療開發已逐漸成為研究熱點與關注的重點,使得脂聯素及其受體的相關研究顯得愈發重要。
1 脂聯素
脂聯素是源于人和動物脂肪組織的一類特異性表達的分泌蛋白[2-3]。在哺乳動物中,主要是由白色脂肪組織特異性分泌呈膠狀樣的內源性細胞激素因子,因此被稱為脂肪細胞補體相關蛋白(Adipocyte complement related protein of 30 kD,Acrp30)[4]、APN[5]、特異性脂肪基因(Adipose most abundant gene transcript l,apM1)[6]、 膠狀蛋白(Gelatin binding protein of 28 kD,GBP28)[7], 即 :Acrp30、APN、GBP28和apM1統稱脂聯素。1999年,Arita等[8]將apM1產物稱為脂聯素,并建立了人體血液中脂聯素濃度的檢測方法。隨后,學術界將該產物統稱為脂聯素。研究表明,在血漿中脂聯素的劑量單位是mg/mL[9]。人體內脂聯素的濃度約為3-30 μg/mL[10],鼠體內脂聯素的濃度約為 2-7 μg/mL,豬體內脂聯素的濃度約為3-4 μg /mL。哺乳動物機體內的脂聯素含量增加,能促進脂肪酸的氧化[11-12],通過對肝臟和肌肉等胰島素靶向器官的攝取,以此增加機體胰島素的增敏作用[13-14]。
1.1 脂聯素結構與類別
GenBank上目前已登錄了人、大猩猩、獼猴、大鼠和小鼠、豬、牛、綿羊、山羊及狗等不同哺乳動物脂聯素基因的序列,并定位于特定染色體上(表1)。人、鼠、豬等哺乳動物脂聯素的同源性達80%以上[6]。Saito等[15]發現,人脂聯素基因位于染色體的3q27上,編碼蛋白含有244個氨基酸,分子量約為30 kD,基因全長約17 kb,人和哺乳動物的脂聯素基因均含有3個外顯子和2個內含子。Dasd[16]在小鼠第16號染色體的B3-B4位點發現脂聯素基因,全長約為20 kb,編碼蛋白有247個氨基酸。大鼠脂聯素基因位于11號染色體,編碼244個氨基酸。研究表明,豬、狗、獼猴、牛和羊的脂聯素基因編碼氨基酸數目幾乎相近[17-18]。
脂聯素由氨基端的信號肽序列(17AA)、非同源或非螺旋可變區域(22AA)、一段由22個氨基酸組成的膠狀樣重復序列區域(65AA)和羧基端類似于C1q因子的球狀區域(137AA)四部分構成[19]。研究表明,脂聯素是由3個相同單體組成的非對稱三聚體結構[20]。在分子間二硫鍵的作用下,通過特定區域被分泌到細胞外,約有65個類似膠原蛋白的氨基酸保守區域[21]。以低分子量(LMW,三聚體,-90 kD,占APN總量25%),中等分子量(MMW,六聚體,-180 kD,占APN總量25%-35%)和高分子量(HMW,12-18倍體多聚合物,360-540 kD,占APN總量40%-50%)這3種形式存在[22](圖1)。
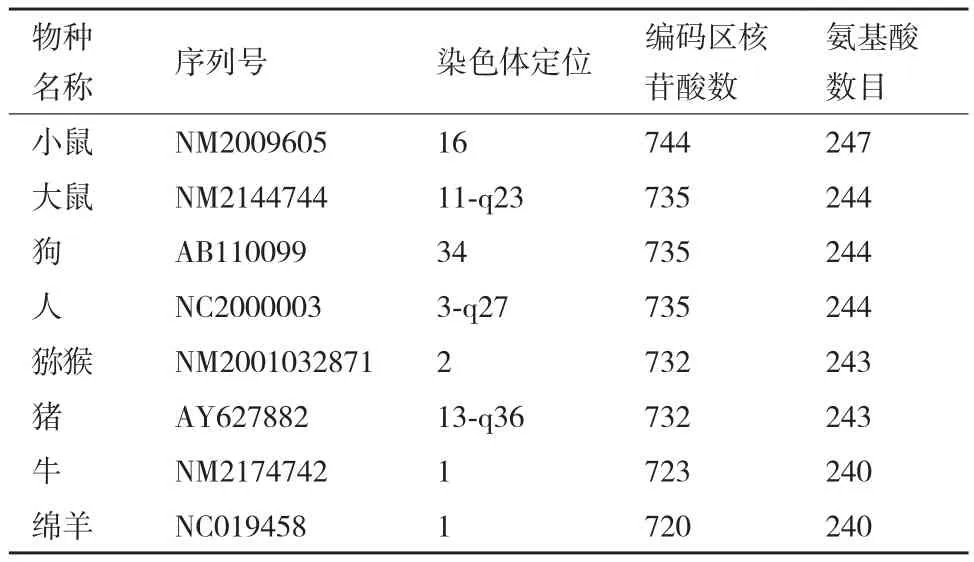
表1 部分哺乳動物脂聯素基因的染色體定位位點及氨基酸數目
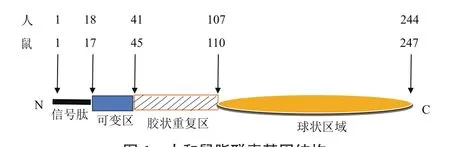
圖1 人和鼠脂聯素基因結構
1.2 脂聯素生物學特性
全長結構域脂聯素(Full of adiponectin,fAd)和球狀結構域脂聯素(Global adiopnectin,gAd)是脂聯素主要的兩種生物活性形式。在脂肪細胞中脂聯素是以單體形式存在,若要發揮其生物學功效,只能以多聚體或是多倍體形式的脂聯素被分泌到細胞外,才能發揮其重要作用。大量試驗已證實,多聚體形式的脂聯素生物學活性最高,具有抗動脈粥硬化、抗糖尿病、抗肥胖、抗消炎的作用,可以阻止胰島素拮抗和心血管疾病的診斷與治療[23]。
值得注意的是,在哺乳動物中,發揮脂聯素基因生物作用的首要場所不是肌肉而是肝臟。這是由于通常在脂肪細胞中,被分泌到細胞外的低分子量脂聯素在形成多聚體與其受體特異性結合后,才能發揮下游的生物學作用;而發揮下游最重要的生物學功能作用,關鍵取決于血液中脂聯素多聚體的分布;而脂聯素多聚體發揮生物學功能作用,又主要取決于由Cys-39介導二硫鍵的結構[9]。從已報道有關人的球狀脂聯素三倍體信號肽的結構可以得知[24],脂聯素結構對其信號傳導代謝,在發揮生物學功能等作用方面極為重要。
2 脂聯素受體
目前,已報道證實的脂聯素受體有3種:脂聯素受體1(AdipoR1)、脂聯素受體2(AdipoR2)和T-鈣黏蛋白(T-cadherin/CDH13)。一般情況下,脂聯素受體均指AdipoR1和AdipoR2。
2.1 AdipoR1和AdipoR2
2003年,Yamauchi等[25]通過構建人骨骼肌的cDNA文庫,從轉染到Ba/F3細胞中首次克隆得到脂聯素基因的兩種受體:AdipoR1和AdipoR2,其大小分別為2.0 kb和4.0 kb。人AdipoR1基因編碼375個氨基酸,AdipoR2基因編碼386個氨基酸,分別位于染色體1p36.13-q41和12p13.31兩個位點[26]。
研究表明,脂聯素受體(AdipoR)有7次跨膜結構的蛋白,與G蛋白偶聯受體家族結構(羧基末端在膜內、氨基末端在膜外)的拓撲結構相反,脂聯素受體的氨基端在細胞膜內,羧基端在細胞膜外,與脂聯素特異性結合后發揮作用[21]。研究還發現AdipoR2比AdipoR1多一個開放閱讀框,二者的羧基端結構相似度高,同源性達到66.7%[14]。人與小鼠AdipoR1的同源性為96.8%,與小鼠AdipoR2同源性為95.2%,說明脂聯素受體的保守程度較高[19]。
脂聯素受體的分子克隆和晶體結構鑒定為人們進一步掌握和了解脂聯素的生物活性及其相關功能的研究提供了一條新的途徑。2015年,Tanabe等[27]第一次報道了人AdipoR1和AdipoR2的晶體結構,證實兩種受體晶體結構基本相似,都是由氨基端的胞內區域、短的胞內螺旋區域、7個跨膜區域以及羧基端的胞外區域組成,而且7個跨膜螺旋結構以順時針方式從第1螺旋到第7螺旋被循環捆綁排列著。最重要的是已證實在兩個受體相似跨膜區內的相對巨大的空腔活動區,存在一個鋅離子結合位點,能促進腺苷酸活化蛋白激酶(AMP-activated protein kinase,AMPK)和過氧化物酶體增殖物激活受 體(Peroxisome-proliferator-activated receptor ɑ,PPARɑ)的活性,增加胰島素的敏感性作用,并成為脂聯素信號傳導通路的一個新靶點[28]。
2017年,Vasiliauskaité-Brooks等[29]報道,AdipoR1和AdipoR2具有酶活性的新功能,在受限制后表現出極低的神經酰胺活性。研究表明,通過脂聯素結合位點,AdipoR2的晶體狀結構能夠與一種游離脂肪酸分子結合,以此加強受體的神經酰胺酶活性[21-29]。實驗報道也證實,脂聯素受體能誘導神經酰胺酶的活性,能夠導致神經酰胺酶和游離脂肪酸的水解[30],為進一步研究脂聯素受體酶活性及其功能提供一些新見解,以此為解決脂聯素受體與底物間的專一性提供理論依據。
2.2 T型鈣黏蛋白/CDH13
脂聯素的第3種受體——T型鈣黏蛋白(T-cadherin/CDH13),位于細胞膜上與高分子量脂聯素的亞型特異性結合的一種截短型鈣黏蛋白。T型鈣黏蛋白沒有跨膜區域,通過糖基磷脂酰肌醇分子附著在細胞膜上。
Hug[31]發現T型鈣黏蛋白不與脂聯素的球狀結構域或脂聯素的三聚體結合,但能作為脂聯素傳導代謝信號的協同受體,主要在內皮細胞和平滑肌中表達。研究發現,脂聯素分子羧基端的球狀結構域存在一個鈣離子的結合位點,可以通過鈣離子介導與T型鈣黏蛋白結合[32]。
T型鈣黏蛋白獨有的結構特性,使得對其研究主要集中在心血管方面。研究發現,在敲除小鼠心臟的脂聯素基因后,血管中T鈣黏蛋白的表達量下降,但用重組脂聯素的作用后,細胞表面的T型鈣黏蛋白表達量會恢復到正常水平[33]。而且T鈣黏蛋白具有減少小鼠心臟負荷的功能[34]。盡管T型鈣黏蛋白在介導脂聯素信號傳導通路的激活機制及其一些功能還不明確,而且在脂聯素受體生物學功能方面的作用也較薄弱。但對機體的健康和疾病控制等方面卻發揮著重要的作用。
3 脂聯素及其受體信號傳導
脂聯素只有與其受體特異性結合后,才能激活脂聯素受體,并和與之相匹配的蛋白銜接作用后,誘導細胞內的信號級聯激活,發揮其生物學作用。研究證實,脂聯素受體與磷酸絡氨酸銜接蛋白(APPL1)作用后,主要通過激活AMPK、PPARɑ和促分裂原活化蛋白激酶(MAPK)通路進行下游糖類和脂肪代謝調節[35]。研究表明,gAd與AdipoR1羧基末端結合后,AdipoR1與APPL1發生作用,促使肝激酶B1(LKB1)與AMPK活化,引起脂肪酸的氧化;而fAd在與AdipoR2羧基末端結合后,AdipoR2同樣在與APPL1的作用下,促使PPARɑ與酰基輔酶氧化酶(ACO)活化,引起脂肪酸的氧化。脂聯素受體通過與APPL1結合后,能誘導神經酰胺酶的活性,降低肝神經酰胺酶水平與促進神經酰胺酶的分解,提升胰島素的增敏作用[36]。因此,機體內脂類代謝通路中的磷酸酪氨酸銜接蛋白(Adaptor protein containing PH domain,PTB domain and leucine zipper motif 1,APPL1)作為信號分子,參與多種細胞信號通路介導、與多種膜受體、核因子和信號蛋白結合,具有調節細胞增殖、染色質重塑、細胞存活等多種生物學功效[37]。通常在骨骼肌中,AdipoR1與AMPK親和性高;在肝臟中,AdipoR2與PPARɑ親和性高(圖2)。研究表明,APPL1能直接增加LKB1在細胞溶質中的轉移和在肌細胞中AMPK的活性。另一方面,APPL1復合體受胰島素或脂聯素的刺激后,在肥胖癥和APPL1的缺陷補充方面有很強的抗胰島素性,參與脂聯素受體和轉運胰島素信號傳導作用,促使APPL1的誘導表達。
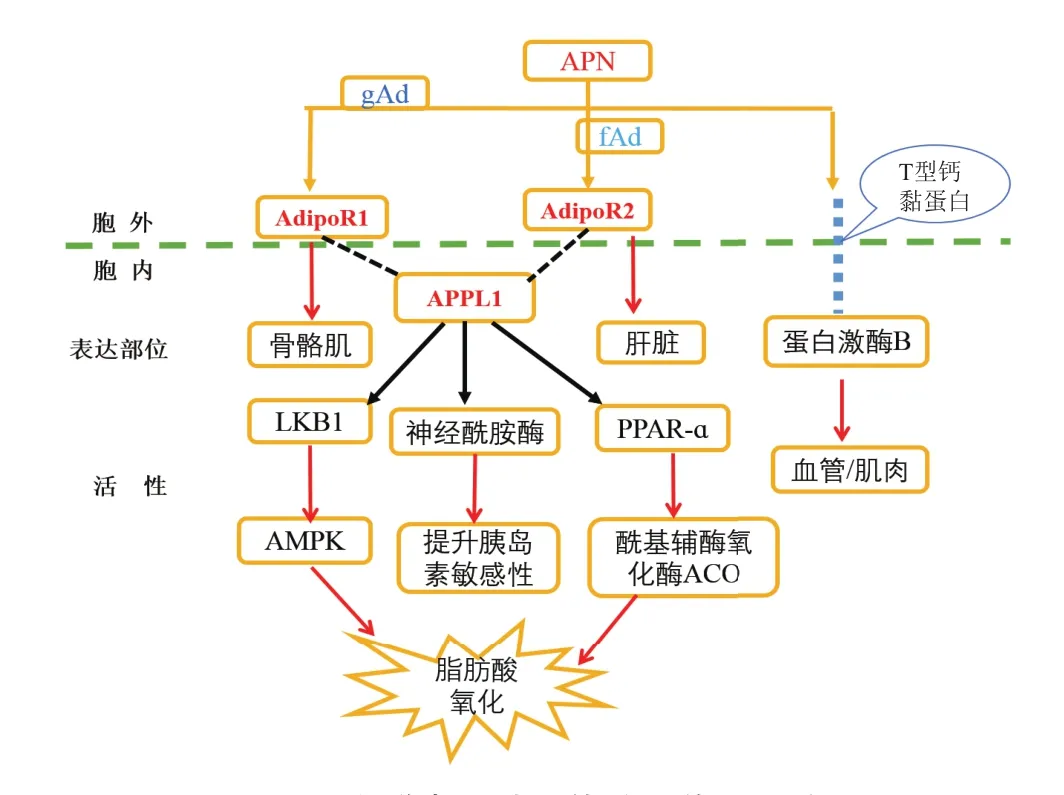
圖2 脂聯素及其受體信號傳導通路
4 脂聯素及其受體基因的表達調控
4.1 脂聯素基因(APN)的表達調控
脂聯素基因參與哺乳動物體內能量代謝調節,主要表現在脂肪組織中脂聯素表達量的高低,如在肥胖小鼠脂肪組織中脂聯素的表達量是顯著降低的[5]。研究發現,重組后的脂聯素能促進體內脂肪代謝,使得血漿中的甘油三酯含量降低,存貯在肝臟和肌肉組織中的甘油三酯含量減少,繼而使得脂肪酸氧化和清除相關蛋白的表達水平上調[38]。盡管機體中脂聯素主要在脂肪組織中表達,在皮下脂肪和內臟脂肪的表達量雖無明顯差異,但表達調控方式不同[9]。
研究表明,通過測定1-21 d新生仔豬脂肪組織中各種脂肪因子的發育性變化情況后,發現脂聯素表達量顯著上升[39]。豬脂聯素及其受體的mRNA,除了在皮下及內臟脂肪組織中大量表達外,在卵巢、子宮也有較高的表達,是影響豬繁殖性狀的相關基因[40-41]。張長慶等[42]用qPCR方法檢測荷斯坦公牛脂聯素基因后,發現在相同組織中脂聯素基因的表達量會因月齡變化而不同,以此反映荷斯坦牛生長發育的變化情況。沈留紅等[43]發現奶牛胎盤中脂聯素mRNA和蛋白表達水平含量與犢牛的出生體重呈正相關,可作為哺乳動物選育的候選基因。李雪梅等[44]發現藏山羊的脂聯素基因在不同組織中以皮下脂肪組織中表達量最高,在不同脂肪組織中以腎周脂肪的表達量最高,而且在不同脂肪組織中的表達差異不顯著,在藏山羊脂肪組織中發揮著重要作用,參與脂肪細胞分化與代謝活動。研究表明,肉牛育肥前期與后期脂聯素基因的mRNA水平在前期與肌內脂肪含量呈正相關,但在后期與肌內脂肪含量呈負相關[45]。De Rosa等[46]證實豬脂聯素的組織特異性表達與其脂肪的沉積有關。Wang等[47]在山羊骨骼肌衛星細胞中檢測到的脂聯素蛋白表達,能促使骨骼肌衛星細胞分化生成明顯的前體脂肪細胞,加快成熟脂肪細胞的生成。張輝[48]通過調節機體內脂肪細胞的分化和脂肪含量,發現脂聯素基因可以改善機體胴體性狀,其表達量與前體脂肪細胞分化及脂肪沉積呈正相關。
由此可知,脂聯素基因的表達水平與調控方式雖不同,但對于但在哺乳動物的繁殖性狀、脂肪細胞的分化、及脂肪酸合成代謝方面存在著密切的相關性,使得脂聯素基因已成為影響哺乳動物脂肪代謝的一個分子標記。
4.2 AdipoR的表達調控
AdipoR1主要在機體的骨骼肌中表達,與gAd親和力高,與fAd親和力較低,能激活AMPK;AdipoR2主要在機體的肝臟中表達,對fAd和gAd均具有中等親和力,能激活PPARɑ,能調節脂肪和葡萄糖代謝,具有抗氧化、抗消炎、抗肥胖癥等功效。而以多聚體結合為主的T鈣黏蛋白脂聯素受體,在蛋白激酶B(AKT)的作用下,主要在機體的血管和肌肉中表達。
Ding等[49]首次克隆出豬脂聯素及其受體基因,豬脂聯素mRNA在脂肪、肝臟、心臟和肌肉等組織中大量表達。Das等[16]發現豬AdipoR1和AdipoR2基因在脂肪組織中能特異性表達,并與人脂聯素受體的同源性較高。Kang[50]發現日本獼猴AdipoR1和AdipoR2的跨膜分布區域保守性高,在包括腦組織的大部分組織中,AdipoR1都有表達。Lemor等[18]對綿羊5種不同脂肪組織中的脂聯素受體表達研究發現,AdipoR1和AdipoR2在內臟脂肪組織中的表達要高于皮下脂肪組織,繼而為小反芻動物脂聯素及其受體基因的生脂機理研究提供理論支持。薛科[51]發現,在山羊的瘤胃中AdipoR1和AdipoR2的表達量較高,表明這兩種受體在瘤胃內發揮著重要的作用,而且還能與酵母中一種結構類似于脂聯素的滲透蛋白相結合并發揮作用,因而推測AdipoR1和AdipoR2可能與其他受體蛋白結合后,將信號傳遞到胞內發揮相應作用。羅軍和趙旺生等[52]發現處于不同泌乳期的薩能奶山羊AdipoR1在肺中表達量最高;在干奶期和泌乳期時,AdipoR1在不同組織間的表達量差異顯著。乳腺上皮細胞用胰島素處理后,AdipoR1的表達量則會下調;用催乳素處理后,AdipoR1的表達量會上調,說明該基因對薩能奶山羊的乳腺上皮細胞有調控作用。Oliveira等[53]通過qPCR和免疫組織化學分析方法,檢測了脂聯素及其AdipoR1和AdipoR2在山羊卵巢卵泡中的表達,驗證出山羊重組脂聯素能增加其體外卵母細胞核的成熟。
綜上可知,在哺乳動物中,AdipoR1和AdipoR2幾乎同時分布于能夠被檢測到的所有組織中,在通過與胞外的脂聯素基因作用后,能激活位于表達部位關鍵信號分子的活性,從而調控機體的脂肪酸氧化代謝和維持機體的能量平衡。值得注意的是,AdipoR1和AdipoR2二者間的表達水平盡管不同,但在每種組織中的表達只有一種受體占據著主導地位。
5 脂聯素及其受體基因的多態性
目前,國內外主要以人、牛、羊、豬、禽類以及魚類為研究對象,對脂聯素及其受體基因展開有關機體生脂機理、脂肪代謝及繁殖性狀等方面的研究。以此探尋脂聯素及其受體在疾病預防和靶點藥物研制對其繁殖與遺傳性狀的影響。Kaklamani等[54]首先報道了人脂聯素及其AdipoR1基因的多態性(SNPs)與結腸癌關聯性研究,但結果存在爭議。后續報道發現,在人類的一些癌癥組織中發現了脂聯素受體的表達,如直腸癌、胃癌、肺癌、胰腺癌和前列腺癌等[55-56]。年士艷等[57]發現脂聯素及其受體基因的多態性與心血管疾病、動脈粥硬化、冠心病等密切相關并且成為疾病候選基因的遺傳標記,為治療如心血管等疾病的一類新型藥物靶點提供參考。
研究表明,秦川牛脂聯素基因第2外顯子的多態性被檢測后發現,有3種不同的基因型與秦川牛的屠宰重、背膘厚、胴體重、眼肌面積等產肉性狀高度相關[58],可以將脂聯素基因作為秦川牛產肉性狀的分子標記。楊磊等[59]對日本和牛和延邊黃牛雜交后代、西門塔爾牛和延邊黃牛雜交后代通過用遺傳學分析后發現,脂聯素基因在這兩大牛群體中具有豐富的多態性,脂聯素基因內含子1上和內含子2上均有2個核苷酸多態位點,為改善牛酮體性狀和生長性能奠定了基礎。劉重旭等[60]研究發現貴州白山羊和古藺馬羊位于脂聯素基因3'端非編碼區的單堿基突變為多態位點,對其繁殖性狀相關性分析表明,貴州白山羊和古藺馬羊中GG型和AG型的產羔數差異顯著。因此,脂聯素基因可作為控制貴州白山羊和古藺馬羊多胎性的一個候選基因或是緊密連鎖的分子遺傳標記物。并且通過對貴州黔北麻羊和黑山羊的脂聯素基因進行多態位點檢測分析后,首次揭示在內含子1和外顯子3中發現5個單堿基突變,發現在這兩種山羊的遺傳多態性較為豐富[61]。
安清明等[62]發現綿羊羔羊脂聯素基因的多態性與其生長及屠宰等不同性狀具有相關性。曹海洋[63]檢測到小尾寒羊脂聯素基因10454位點A/G突變,顯著影響第四胎產羔數,10677位點突變對第二胎產羔數影響較為顯著,可知脂聯素基因與繁殖性狀間存在相關性。Houde等[64]研究發現,長白母豬脂聯素受體基因中存在11個多態位點(其中AdipoR1有6個多態位點,AdipoR2基因有5個多態位點)。這有助于增加平均窩產仔數、減少木乃伊豬、縮短斷奶與發情間隔等特異性相關多態性的選擇,提高長白豬的生產效率。周寧[65]發現豬脂聯素基因有3個多態位點,AdipoR1基因有1個多態位點,AdipoR2基因有1個未見報道的多態位點,均與母豬的繁殖力有相關性。
因此,脂聯素及其受體的多態性與哺乳動物的繁殖性狀、生產性狀、胴體性狀、肉品質及心血管疾病等方面具有相關性,將為動物選育、繁殖性能及生理營養調控等方面提供理論支持與科學依據,將對哺乳動物機體的健康、疾病預防和臨床診斷等有重要指導意義。
6 總結與展望
隨著生物醫藥研究領域技術的飛速發展,促進了脂聯素及其受體研究的快速發展。自1995年脂聯素被發現至今的24年的研究結果表明,脂聯素不單是機體內能量貯存和利用的場所,還是直接影響機體內脂類代謝與合成,促進脂肪酸氧化和增加抗消炎作用的重要因子。在抗胰島素、抗動脈粥硬化、抗心血管疾病和2型糖尿病等方面發揮著重要作用,并在增加胰島素敏感性,抗細胞凋亡、抗消炎、抗纖維化、抗脂類毒性及促進神經酰胺減少,刺激著鞘氨醇磷酸酯的生成,促使纖維原細胞活性增強[66-67],在能量代謝中起著關鍵的調節作用。
在哺乳動物中,通過對機體內脂肪代謝進行調控,具有高潛力的脂聯素及其受體基因在脂肪細胞分化、脂質代謝與沉積以及在畜禽經濟性狀方面發揮著重要的作用。動物生產中,若機體內脂肪沉積過多,則會降低飼料利用率,增加養殖成本。若機體內脂肪沉積太少,則會對動物肉品品質產生影響。脂聯素作為機體內脂肪代謝的主效基因,在動物生產和分子育種中,已成為代謝綜合癥重要的靶向分子標記。因此,對哺乳動物脂聯素及其受體基因開展功能性細胞因子的相關研究,為加快脂聯素及其受體基因的研究及運用,提供必要的理論依據和數據支持,并在動物疾病預防及靶向藥物臨床診斷等方面具有重要的指導意義。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
