奶牛分娩后早期血漿代謝物變化研究
2020-07-10 06:42:14張萌羅芳王敏武彥澤王俊奎和東遷陳麗堯陶金忠
生物技術通報 2020年6期
張萌 羅芳 王敏 武彥澤 王俊奎 和東遷 陳麗堯 陶金忠
(1. 寧夏大學農學院,銀川 750021;2. 內蒙古圣牧控股有限公司,巴彥淖爾 015000;3. 寧夏上陵牧業有限公司,青銅峽 751600)
圍產期是從產前3周到產后3周,這個階段奶牛機體代謝發生巨大的變化[1]。主要體現在隨著分娩的臨近,奶牛干物質的攝入不能滿足自身能量消耗,從而導致奶牛負能量平衡(Negative energy balance,NEB)[2]。NEB 損害奶牛的健康和影響生產,并導致免疫力下降[3]。當奶牛發生NEB時通常引起脂肪肝和酮病[4],而這兩種疾病均能造成肝功能的損傷[5-6]。肝臟是能量代謝的主要器官,奶牛體內的糖代謝主要來自于肝臟糖異生[7]。天冬氨酸(Aspartic acid,ASP)、組氨酸、蛋氨酸、脯氨酸和絲氨酸的濃度在患有酮癥的奶牛中下降[8]。這些氨基酸的濃度降低,因為它們在糖異生過程中被大量使用[9-10]。而在分娩后的幾天內,乳腺對葡萄糖、氨基酸和脂肪的需求量成倍增加,肝臟糖異生和脂肪動員的速度大大加快[11-12]。因此,奶牛分娩后血漿代謝物發生顯著變化,影響奶牛的健康。代謝組學是一種高通量、高靈敏度的現代分析技術。目前,代謝組學方法已成功地應用于奶牛產后血漿生物標志物的篩選[13]。
為此,本研究通過對分娩當天和產后第7天血漿代謝組學研究,探究分娩后早期奶牛血漿代謝物的變化情況,探討分娩后血液代謝組變化,旨為了解分娩后血漿代謝物變化對奶牛健康狀況提供參考。
1 材料與方法
1.1 材料
1.1.1 試驗儀器和設備 超高效液相:1290 UHPLC(美國Agilent公司);高分辨質譜1:QTOF 6550(美國Agilent公司);高分辨質譜2:Triple TOF 6 600(AB Sciex);天平:BSA124S-CW(Sartorius);研磨儀:JXFSTPRP-24(上海凈信科技有限公司);超聲儀:PS-60AL(深圳市雷德邦電子有限公司);純水儀:明澈D24 UV(Merck Millipore);色譜柱:ACQUITY UPLC BEH Amide 1.7 μm 2.1×100 mm(Waters)。
1.1.2 試驗材料和試劑 醋酸銨(CNW Technologies);乙腈(CNW Technologies);氨水(CNW Technologies);甲醇(CNW Technologies);L-2-氯苯丙氨酸(上海恒柏生物科技有限公司)。
1.2 方法
1.2.1 試驗動物分組和血液的采集保存 在寧夏某大型集約化奶牛場,選擇15頭2-3胎、預產期和體況相近(體況評分BCS 3.0-3.5)的健康荷斯坦奶牛,奶牛奶產量(分娩當天由于采集初乳無法統計)和胎次的均值±SD,見表1。分別在分娩當天(A組)和產后第7天(B組)晨飼前(5:00-6:00)尾靜脈采集血液10 mL,使用肝素鈉抗凝,并以3 000 r/min離心10 min進行分離,收集上層血漿于冷凍管中,標記后置于-80℃冰箱冷凍保存,以備后續分析。

表1 奶牛奶產量和胎次
1.2.2 檢測方法
1.2.2.1 代謝物提取 參照張萌等[14]的代謝物提取方法。
1.2.2.2 上機檢測 使用的色譜柱為購自Waters的UPLC BEH Amide 色譜柱(1.7 μm×2.1×100 mm)。進樣體積為2 μL。按照表2中的流動相參數在安捷倫1290超高效液相控制下進行分析。
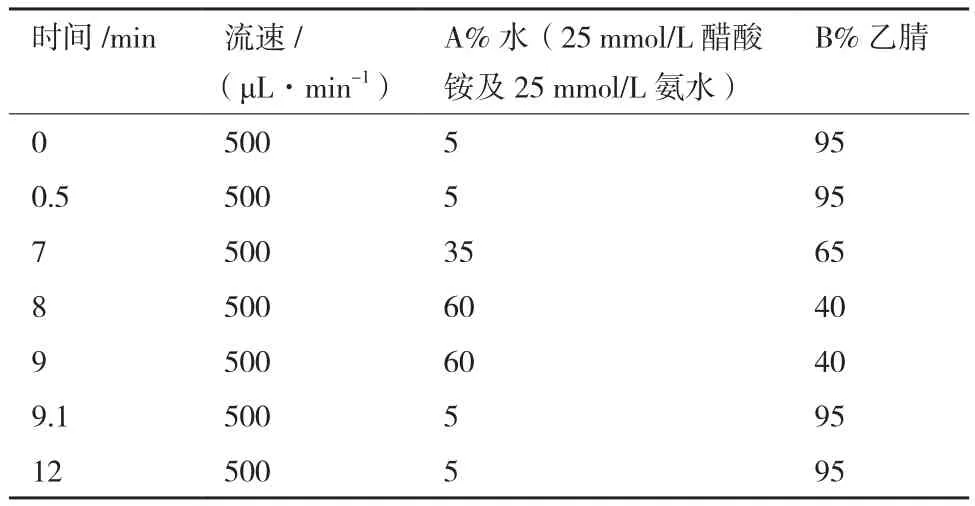
表2 液相色譜流動相條件
控制軟件(Analyst TF 1.7,AB Sciex)控制下AB 6600 Triple TOF & Agilent 6550 QTOF質譜儀基于IDA功能進行一級、二級質譜數據采集。每個數據循環采集中,篩選出強度最強且大于100的分子離子進行采集對應的二級質譜數據。轟擊能量:30 eV,每50 ms15張二級譜圖。ESI離子源參數設置如下:溫度:650℃,輔助氣壓:60 Psi,氣簾氣壓:35 Psi,霧化氣壓(GS1):60 Psi,噴霧電壓:5 000 V(正離子模式)或-4 000 V(負離子模式)。
1.2.3 數據處理 原始數據經ProteoWizard軟件處理,將數據轉換成mzXML格式。采用XCMS程序對轉化后數據進行峰對齊、提取峰面積和峰積分等處理工作。檢查數據的完整性和缺失值狀況,補充或刪除數據中的極值和數據中組內缺失值大于50%的離子峰。內標校正和歸一化處理數據,使得代謝物之間和各樣本之間均可以進行平行比較。
1.2.4 統計學分析 使用SIMCA軟件對處理后數據進行模式識別和Pareto-scaling預處理。對處理后數據進行多元統計分析,無監督主成分分析(Principal component analysis,PCA),正交偏最小二乘判別分析(Orthogonal partial least squares-discriminant analysis,OPLS-DA)。構建表達模型,并對模型中的數據進行置換檢驗(Permutation test),計算根據OPLSDA模型得到變量投影重要度(Variable importance in the projection,VIP)。對數據進行單變量統計分析包括:變異倍數(Fold change,FC)和Students t-test分析。
1.2.5 差異代謝物鑒定和生物學信息學分析 經過多元統計分析得到VIP大于1的值,用產后第7天均值比上分娩當天均值得到FC值,結合P值。以VIP>1初步篩選出各組間的差異物,單變量統計分析P<0.05和FC>1.3或FC<0.77進一步篩選顯著性差異代謝物。顯著性差異代謝物使用Origin 8.0進行ROC Curve分析得到AUC面積大于0.8的代謝物。將篩選出的顯著性差異代謝物在線分析平臺MetaboAnalyst進行聚類分析和代謝通路分析。
2 結果
2.1 代謝譜分析
由圖1可知,QC樣本TIC出峰保留時間和峰面積都重疊很好,表明在整個實驗過程中儀器穩定性很好。由圖2可知,QC樣本相關性越接近于1,說明整個方法穩定性越好數據質量越高。QC樣本相關性很高,表明數據質量很高。
2.2 多元統計學分析、
由圖3可知,對分娩當天(A組)和產后第7天(B組)的PCA分析,觀察所有樣本之間的總體分布趨勢,表明正、負離子模式數據下A組和B組有一定的分離趨勢。由圖4可知,建立A組和B組的OPLS-DA模型,得到的模型評價參數正離子模式R2Y=0.959和負離子模式R2Y=0.973。R2越接近1表明模型越穩定可靠,本試驗正負離子均R2Y≥0.95,表明模型穩定可靠。由圖5可知,顯示了基于正、負離子模式數據兩組OPLS-DA模型的置換檢驗圖,正、負離子模式數據置換檢驗截距,分別是Q2=-
0.340(正離子模式)和Q2=-0.329(負離子模式)。Q2intercept<0說明本試驗正、負離子模式數據建立的OPLS-DA模型均未發生過度擬合。
2.3 差異代謝物的篩選
以 VIP>1、P<0.05、FC>1.3或FC<0.77進一步篩選顯著性差異代謝物。分娩當天和產后第7天兩組共篩選出17種顯著性代謝物。正離子模式下篩選出9個顯著性差異代謝物,其中4個代謝物產后第7天顯著低于分娩當天組,5個代謝物產后第7天顯著高于分娩當天組(P<0.05)。負離子模式下篩選出8個顯著性差異代謝物,其中2個代謝物產后第7天顯著低于分娩當天組,6個代謝物產后第7天顯著高于分娩當天組(P<0.05)。正、負離子模式下鑒定出的差異代謝物見表3。
2.4 生物信息學分析和生物標記物篩選
利用ROC曲線對顯著性差異代謝物進行分析篩選,考察這些顯著性差異代謝物的分類識別能力。當ROC曲線下的面積(Area under curve,AUC)大于0.5識別能力較好,AUC值越接近于1,識別效果越好。如圖6,結果顯示A組和B組共有11個代謝物AUC面積在0.8以上。篩選出11個代謝物產后第7天比分娩當天全部上調:磷脂酰膽堿(16∶0/16∶0)、二醇膽酸、脯氨酸-絲氨酸、脯氨酸-甘氨酸、1-硬脂酰基-sn-甘油3-磷酸膽堿、乳清、L-天冬氨酸、N-乙酰基-L-天冬氨酸、異戊酰甘氨酸、L-肉堿和苯甲酸。
利用MetaboAnalyst分析平臺對ROC曲線篩選出的顯著性差異代謝物進行聚類分析和KEGG代謝通路分析。圖7顯示了正負離子模式A組和B組顯著性差異代謝物層次聚類結果。結果發現同組樣品很好聚類在同一簇中,聚在同一簇內的代謝物具有相似的表達模式,表明篩選的顯著性差異代謝物合理且準確。由圖8可知,A組和B組兩組11個顯著性差異代謝物共參與11種不同代謝通路。11個代謝通路分別是:丙氨酸,天冬氨酸和谷氨酸代謝、亞油酸代謝、α-亞麻酸代謝、精氨酸生物合成、組氨酸代謝、泛酸和CoA生物合成、β-丙氨酸代謝、甘油磷脂代謝、花生四烯酸代謝、嘧啶代謝和氨基酰基-tRNA生物合成。主要代謝通路以影響力大于0.2(Pathway impact>0.2)為篩選標準,Pathway impact>0.2代謝通路丙氨酸,天冬氨酸和谷氨酸代謝。甘油磷脂代謝和嘧啶代謝兩組影響力相對較小(Pathway impact>0.05)。
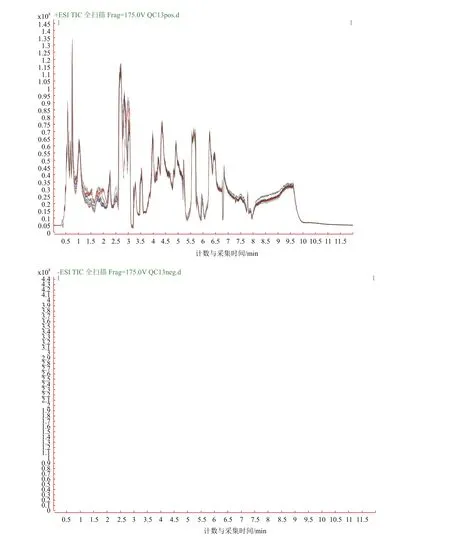
圖1 QC樣本TIC圖
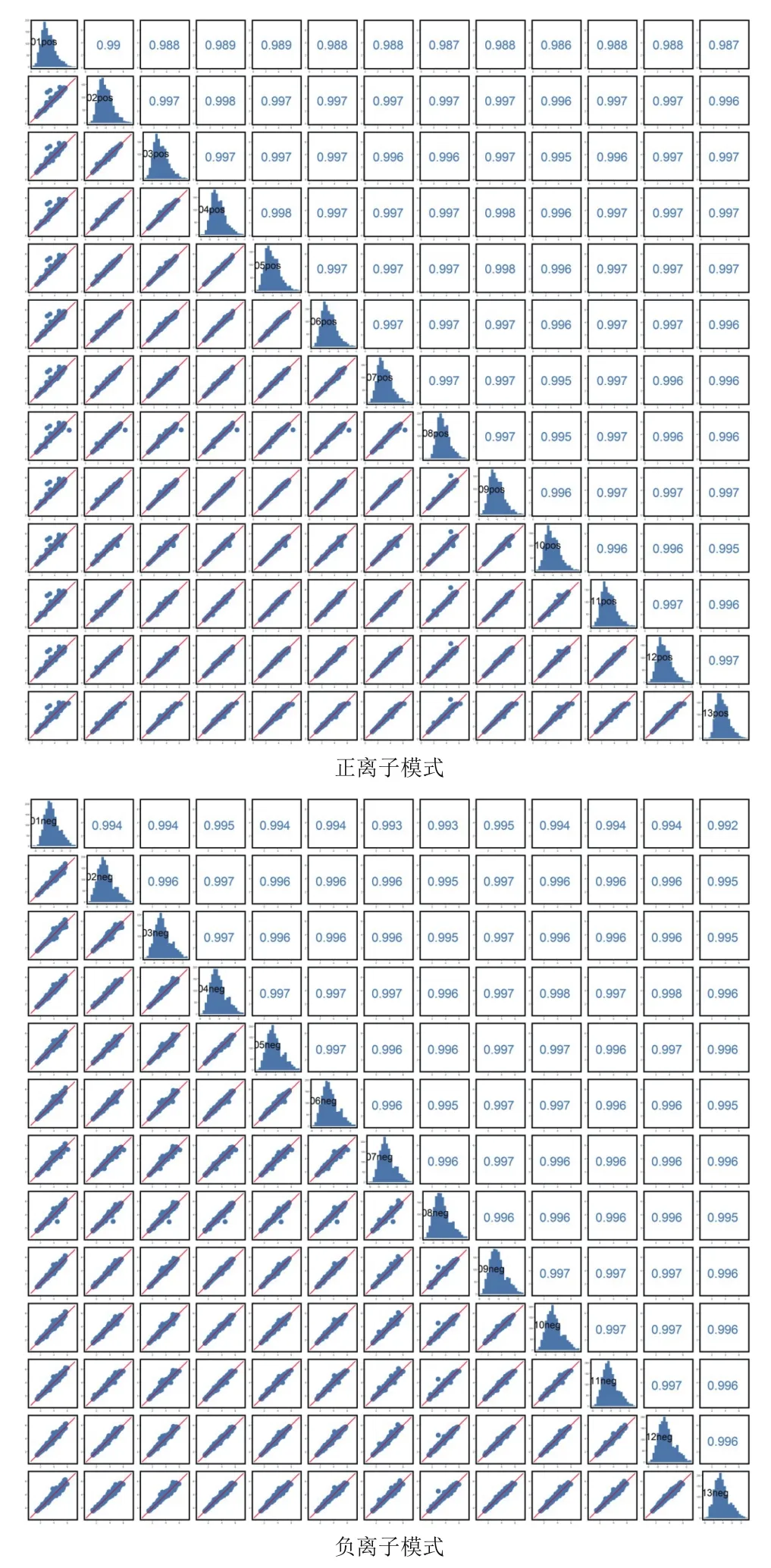
圖2 正離子和負離子QC樣本相關性分析
3 討論
3.1 丙氨酸,天冬氨酸和谷氨酸代謝
N-乙酰 -L-天冬氨酸(N-acetyl-aspartic acid,NAA)是存在于哺乳動物大腦中的游離氨基酸[15],在線粒體由游離ASP乙酰化以乙酰輔酶A(CoA)為輔助因子合成[16]。NAA被認為是多種神經系統疾病的生物標志物[17]。ASP是一種重要的非必需氨基酸,主要參與尿素循環、糖異生和蘋果酸-天門冬氨酸穿梭等多種生化過程[18]。這些生化過程主要發生在肝臟,當ASP含量降低時導致氨積累,且引起肝臟損傷[19-20]。ASP含量的增加表明通過糖異生等過程產生的能量增加[19]。ASP的乙酰化可以促進其從線粒體中去除,有利于谷氨酸轉化為酮戊二酸,從而進入三羧酸循環進行能量生產[16]。有研究表明丙氨酸、ASP和谷氨酸代謝在淋巴細胞再生和免疫過程中起著重要作用[21]。在本研究中NAA和ASP在產后第7天相對分娩當天上調,可能是在丙氨酸,天冬氨酸和谷氨酸代謝中參與能量代謝,維持分娩后泌乳需要大量的能量。這可能是為了增加機體能量的產生,減輕對肝臟的影響,維持奶牛分娩后的機體健康。
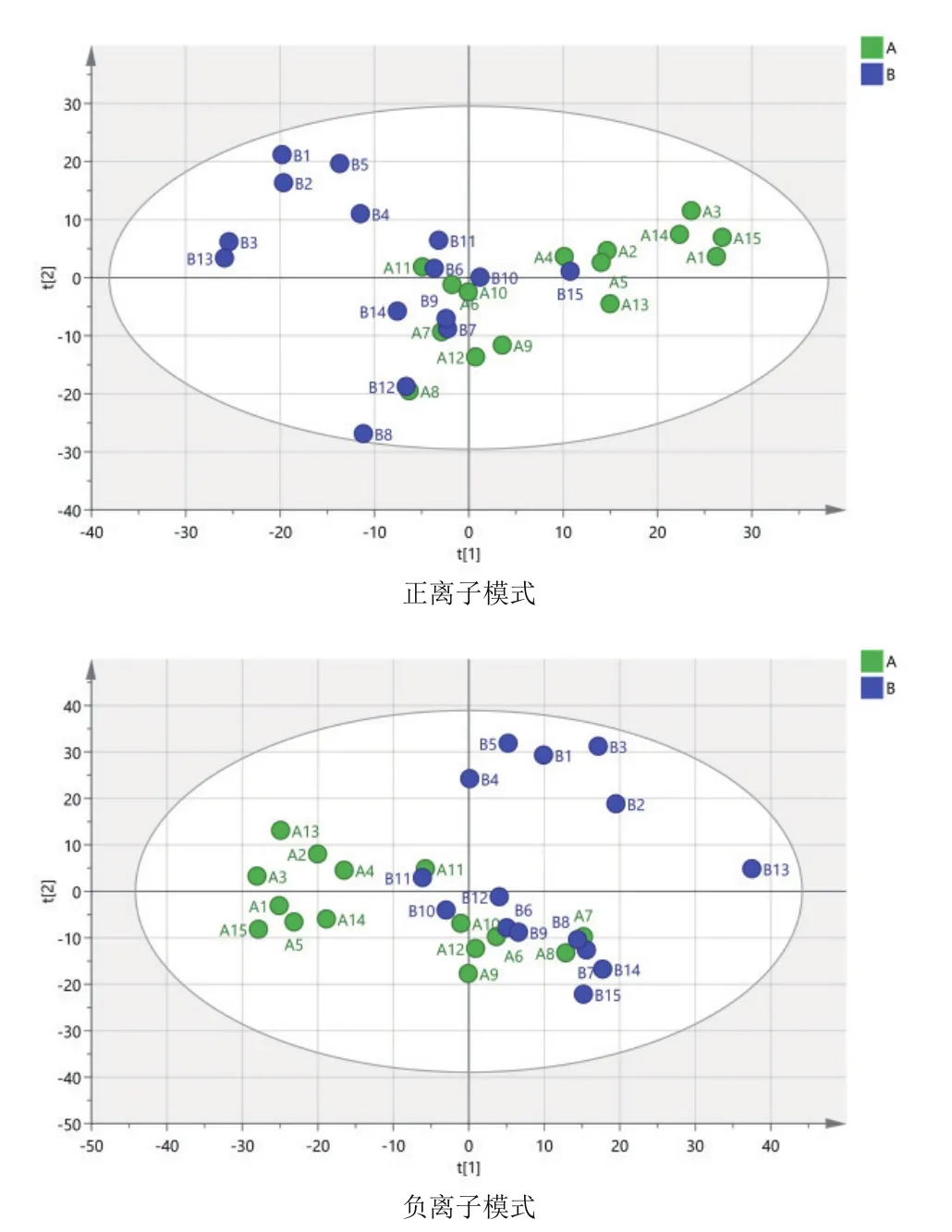
圖3 A組和B組兩兩比較正、負離子模式下PCA圖譜
3.2 甘油磷脂代謝
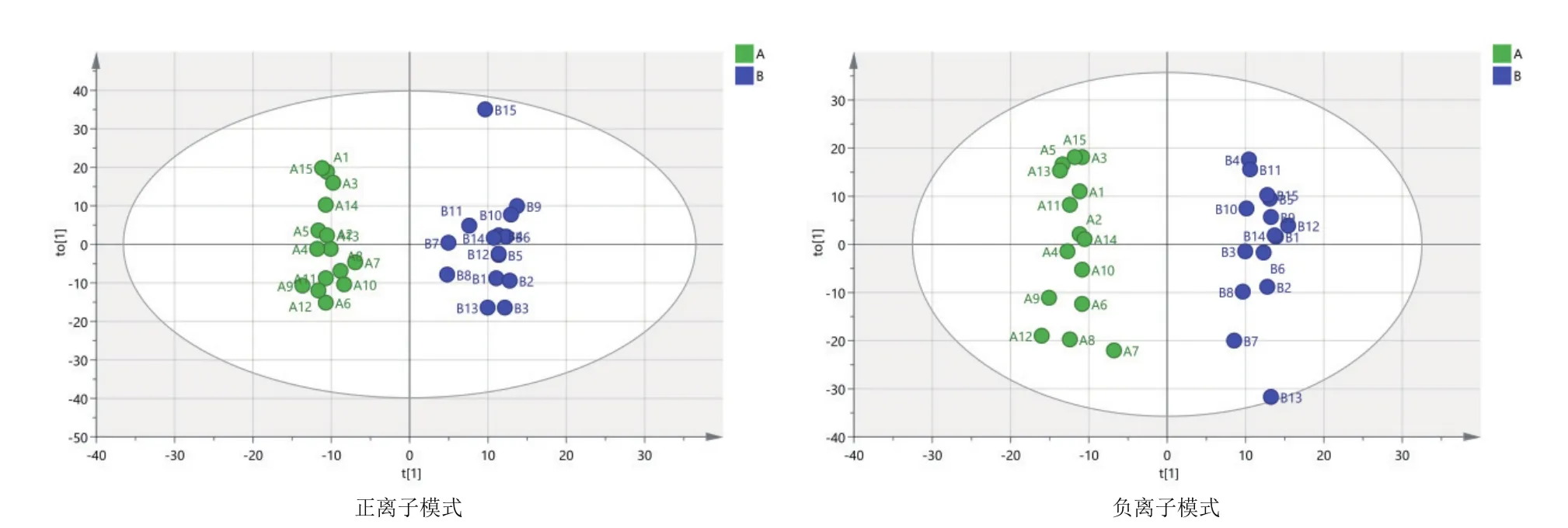
圖4 A組和B組兩兩比較正、負離子模式下OPLS-DA圖譜
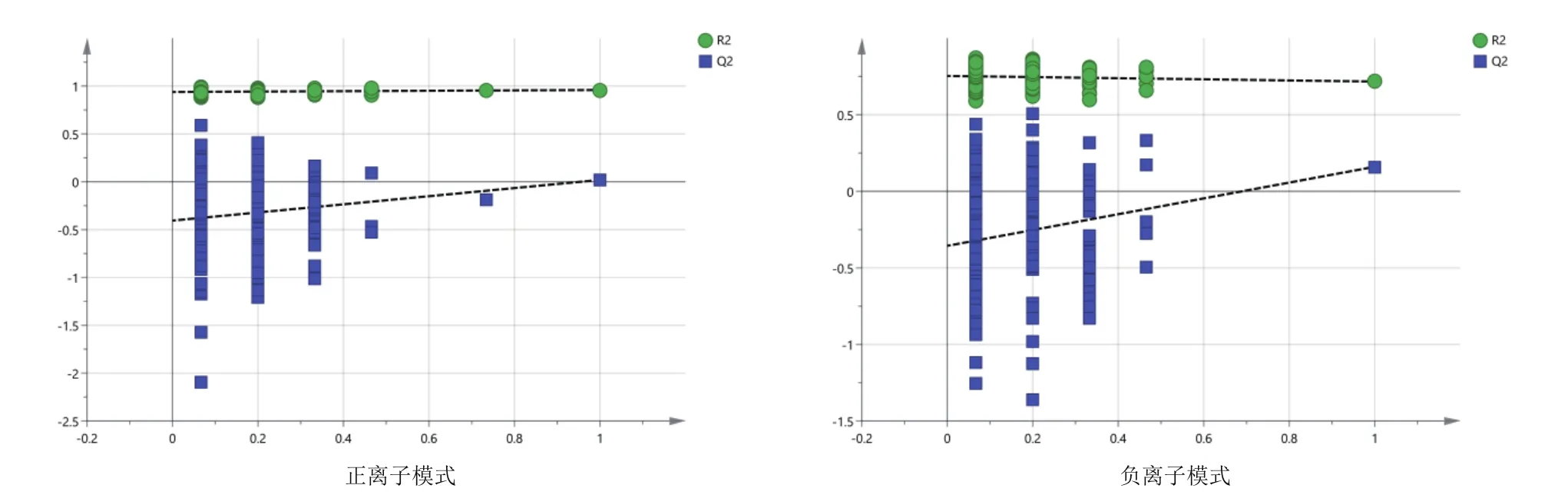
圖5 A組和B組正、負離子模式下OPLS-DA模型置換檢驗圖
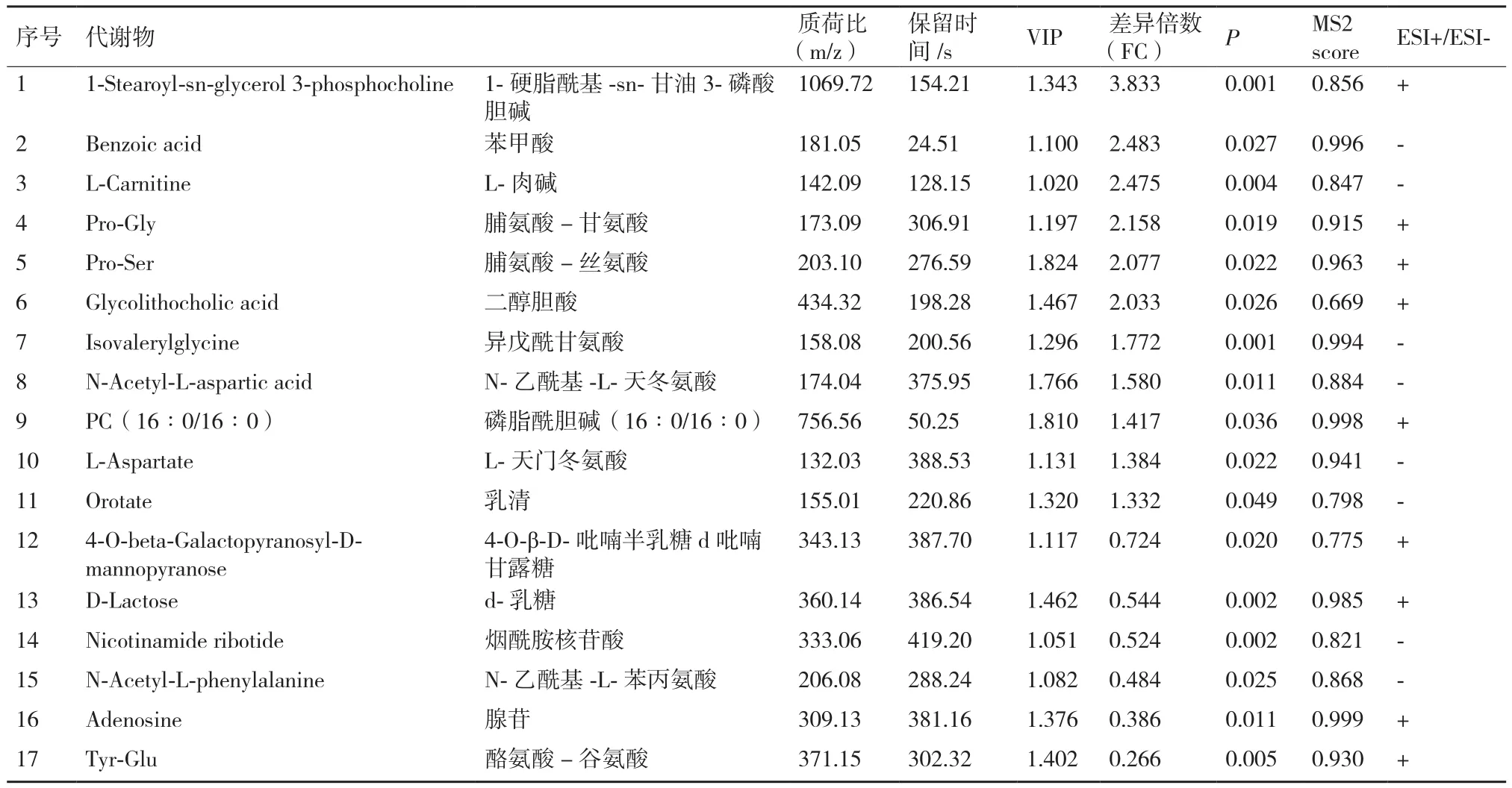
表3 A組和B組正、負離子模式下鑒定的差異代謝物
磷 脂 酰 膽 堿(Phosphatidyl cholines,PC) 是細胞膜的主要磷脂成分和生產脂質第二信使的底物[22],也是肝臟三酰甘油(Triacylglyceride,TAG)分泌所必需的甘油磷脂[23]。PC主要參與甘油磷脂代謝,甘油磷脂是比較復雜的代謝,代謝中主要有PC和磷脂酰乙醇胺(PE)兩種甘油磷脂[24]。而PE轉化為PC是肝臟特異性的PC合成途徑[25]。PC是促進肝臟中極低密度脂蛋白(Very low density lipoprotein,VLDL)轉運甘油三酯必需的物質[26]。血液中PC水平的升高會導致奶牛肝臟中VLDL的分泌增加[27]。從妊娠到哺乳期血漿PC減少與產后脂肪肝的發展有關[28]。血漿中PC從泌乳早期到泌乳后期增加了10倍[29]。奶牛血漿中PC的濃度從分娩前2周開始下降,分娩后開始上升到第4周達到高濃度[30]。有研究表明PC可以作為產后奶牛脂肪肝血漿脂質評價生物標志物[31]。本研究從分娩當天到產后第7天血漿PC濃度升高,與前人研究結果一致。說明分娩后PC濃度增加,使得奶牛肝臟中VLDL的分泌增加,增強了TAG從肝臟中轉運出來的速度,降低了產后奶牛患脂肪肝的幾率,降低了奶牛分娩后負能量平衡的發生機率。
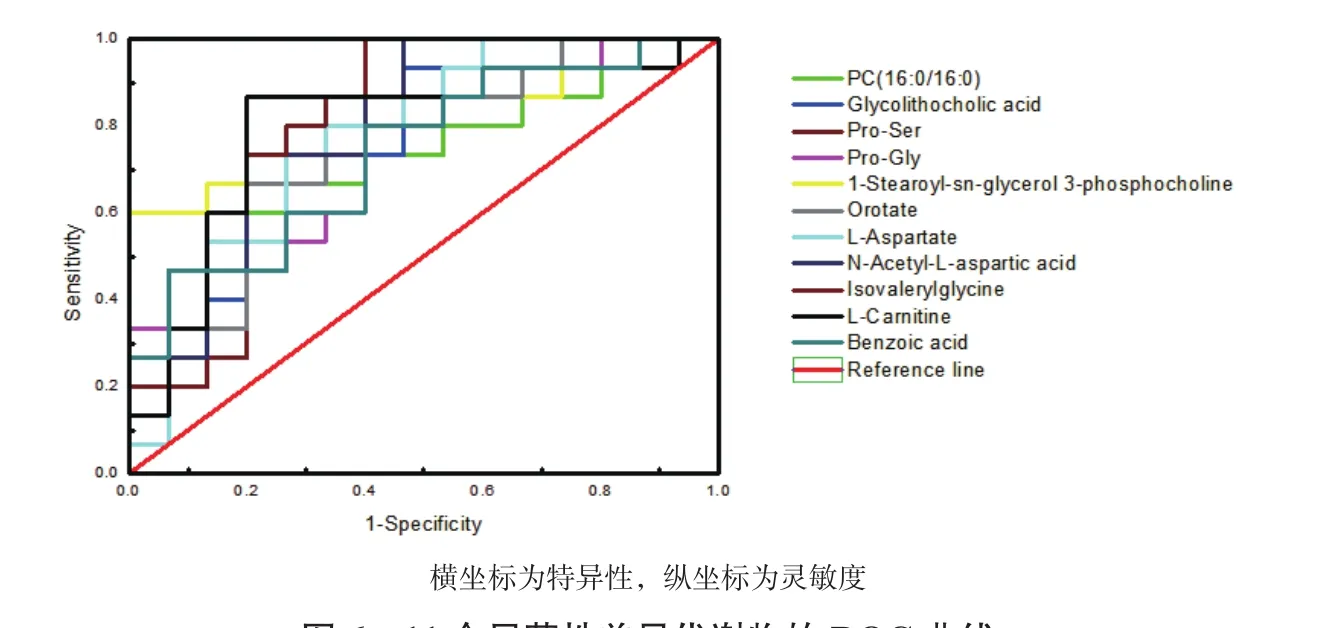
圖6 11個顯著性差異代謝物的ROC曲線
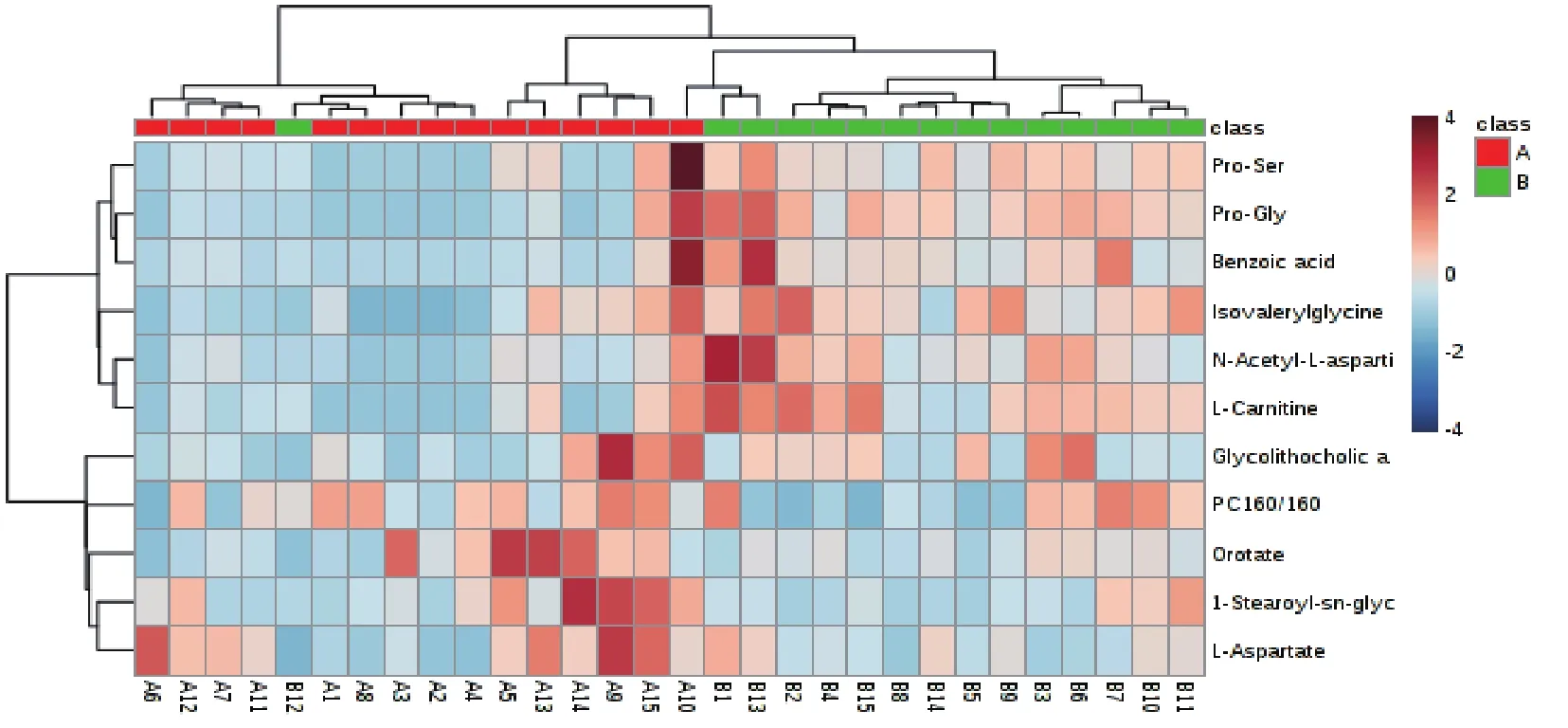
圖7 A組和B組顯著性差異代謝物聚類熱圖
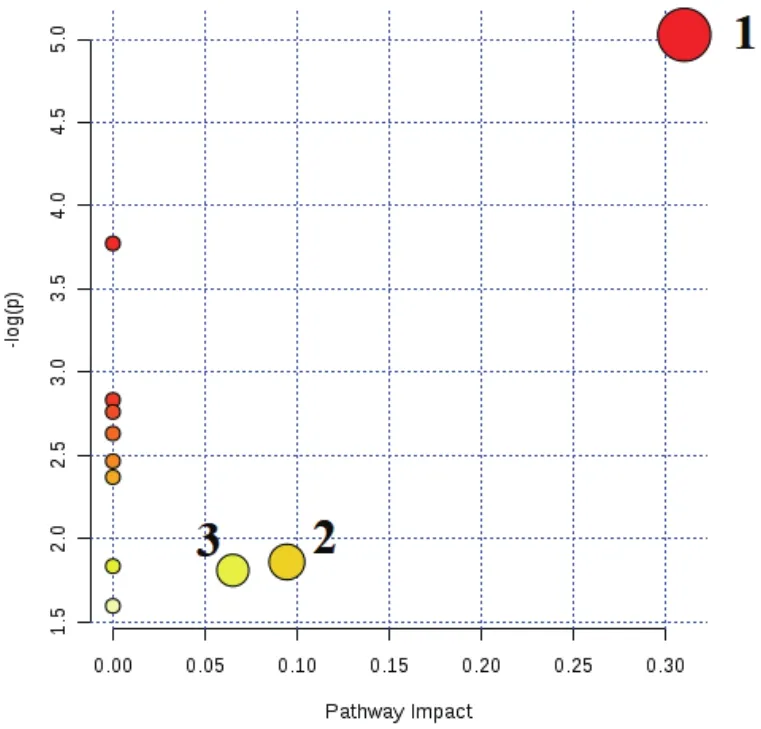
圖8 A組和B組差異代謝物的代謝通路分析
3.3 嘧啶代謝
乳清酸(Orotic acid,OA)是核酸堿基的前體,也是嘧啶合成的中間體。OA的多少與奶牛的來源、飲食和泌乳期有關[32]。比較泌乳奶牛和非泌乳奶牛乳腺組織表達時發現,OA是奶牛泌乳關鍵代謝產物,泌乳組奶牛OA顯著高于非泌乳組奶牛[33]。OA的再循環通常可以在肝臟中發生,肝細胞將其吸收并轉化為尿苷,用于嘧啶循環途徑[34]。嘧啶核苷酸合成的最后一步將OA轉化為有活性的尿酶[35]。當腎臟受到嚴重損害時,尿酶活性較低[36]。本研究產后第7天OA相比分娩當天濃度上調,增加的OA可能是和泌乳密切相關,升高的OA可能在嘧啶代謝中轉化為尿酶,從而降低對腎臟損傷,有利于維持奶牛分娩后機體健康。
4 結論
本研究通過分析分娩當天和產后第7天奶牛血漿代謝物,發現產后第7天與分娩當天相比NAA、天冬氨酸、PC和OA上調。這些代謝物變化有利維持奶牛分娩后機體健康。
猜你喜歡
現代臨床醫學(2022年4期)2022-09-29 07:38:00
音樂探索(2022年2期)2022-05-30 21:01:37
昆明醫科大學學報(2021年4期)2021-07-23 01:21:50
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
云南醫藥(2019年3期)2019-07-25 07:25:14
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
海南醫學(2016年8期)2016-06-08 05:43:00
醫學研究雜志(2015年9期)2015-07-01 17:28:15
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
