元寶楓籽對小鼠腸道菌群生態的影響
2020-07-13 11:48:58孫朋浩薛玉環吳永繼朱曉巖趙善廷
食品科學 2020年11期
孫朋浩,薛玉環,吳永繼,任 瑋,鄭 偉,朱曉巖,趙善廷,*
(1.西北農林科技大學動物醫學院神經生物學實驗室,陜西 楊凌 712100;2.西北農林科技大學資源環境學院,農業農村部西北植物營養與農業環境重點實驗室,陜西 楊凌 712100)
元寶楓為槭樹科槭屬植物,為中國特有樹種,其籽仁含油率可高達48%,最近研究發現元寶楓油中含有5%~6%的神經酸[1-4],元寶楓籽是一種營養豐富的天然食品,民間早有炒食元寶楓籽的習慣。2011年3月22日中華人民共和國衛生部第9號文件批準元寶楓籽油為新資源食品,自此元寶楓籽油正式進入我國食用植物油行列。已有研究表明元寶楓籽油具有促進大腦發育、修復受損神經元、抑菌及抗腫瘤的作用[2-4]。
2000年,諾貝爾生理學/醫學獎獲得者Lederberg提出:人是由人體自身的真核細胞及與人共生的微生物細胞共同組成的“超級生物體”[5]。2016年Sender等重新估算了人體腸道細菌的數量,發現其與人體細胞的數量比約為1.3∶1[6]。這些細菌編碼330多萬個基因,大約是人類基因組150 倍,可謂人類的“第二基因組”[7]。腸道菌群與宿主進行著活躍的代謝交換和共代謝,參與機體的免疫調劑、神經調節、物質消化和吸收等諸多方面內容[8-9]。腸-腦軸是腸和腦之間的信息交流系統途徑,越來越多的研究發現,腸道菌群能夠通過腦-腸軸影響腦的活動甚至宿主行為[8]。
將元寶楓籽作為食品,如何影響腸道菌群?腸道菌群在消化系統中扮演了重要的角色,宿主腸道菌群的建立起始于出生后,到了成年時期,腸道菌群已經基本穩定,對非內源性的細菌具備了高度的抗定植能力[9-11]。腸道菌群的結構組成受到多種因素的影響:遺傳背景、生活環境、飲食等,其中飲食結構對于菌群結構變化的影響最為深刻[12-15]。飲食可通過多種機制影響腸道菌群的組成和活性,目前認為,飲食可以給予某些特定的能利用其成分的細菌選擇性生長優勢,從而成為改變腸道微生態的主要驅動力[12-15]。早期的體外實驗發現元寶楓籽具有抑菌的作用[3],本實驗旨在探究元寶楓籽作為食品對宿主腸道菌群的調整作用以及對有害菌的抑制作用。
如何觀察腸道菌群的組成結構及其變化?微生物純培養技術是研究微生物學的經典方法,然而,微生物的數量和多樣性巨大,至今僅有極小部分物種被成功分離培養[16]。隨著高通量測序技術的發展和測序價格的下降,標記基因分析方法已經成為微生物組學分析的有力工具[17-19]。本實驗通過Illumina HiSeq 2500測序系統對細菌16S RNA編碼區域進行擴增注釋,以評估元寶楓籽作為食品對小鼠腸道菌群結構的影響以及對腸道內有害菌的抑制作用。
1 材料與方法
1.1 動物、材料與試劑
8 周齡清潔級雄性昆明小鼠,體質量約28~30 g,購于第四軍醫大學,生產許可證號:SCXK(軍)2002-005。
元寶楓籽購于遼寧彰武,炒制。
DNA提取試劑盒 上海MO BIO Laboratories公司;HiSeq Rapid SBS Kit v2測序試劑盒、文庫構建TruSeq DNA PCR-Free Sample Prep Kit 美國Illumina公司。
1.2 儀器與設備
Qubit 2.0實時熒光定量聚合酶鏈式反應儀 美國Thermo Fisher公司;HiSeq 2500系統 美國Illumina公司。
1.3 方法
1.3.1 小鼠分組
將小鼠隨機分為對照組(C)和實驗組(Y),每組各10 只。在實驗組的日糧中每天加入1.5 g元寶楓籽,連續飼喂2 周,對照組正常飼喂。
1.3.2 小鼠糞便的采集
將小鼠置于干凈空曠的鼠籠中,待糞便排出后,迅速用無菌鑷子夾取糞便于無菌1.5 mL EP管中并置于冰盒中保存;同一批樣品采集完成后,統一將裝有糞便的EP管用液氮處理,暫時保存于-80 ℃冰箱中。
1.3.3 糞便樣品預處理
稱取220 mg糞便于2 mL微量離心管中,加入1.4 mL裂解緩沖液,漩渦混勻,70 ℃孵育5 min,重復2 次。吸取1.2 mL上清液至2 mL微量離心管,加入1 片InhibitEX,立即漩渦1 min至InhibitEX片完全懸浮,室溫下孵育1 min,吸附破壞樣品中的DNA破壞物質和PCR抑制劑,離心3 min,吸取所有上清液至1.5 mL離心管,再離心3 min。
1.3.4 DNA提取
根據試劑盒說明書,從預處理后的樣品中提取總細菌DNA。通過A260nm/A280nm和A260nm/A230nm的比率評估DNA質量和數量,然后將DNA儲存在-80 ℃條件下,待進一步處理。
1.3.5 16S rDNA測序分析
用通用引物(F:5’-ACTCCTACGGGAGGCAGCA-3’;R:5’-GGACTACHVGGGTWTCTAAT-3’)結合適配序列和條形碼序列擴增細菌16S rRNA基因的V3~V4區,進行聚合酶鏈式反應(polymerase chain reaction,PCR)擴增。最后用Quant-it?dsDNA-HS試劑對所有的PCR產物進行定量,并匯集在一起。在HiSeq 2500平臺(2×250 對末端)上對純化的匯集樣本進行高通量測序。
1.3.6 高通量測序數據分析
根據下機序列之間的重疊關系,將HiSeq測序得到的雙端序列數據拼接成一條序列Tags,同時對序列的質量和合并的效果進行質控過濾。使用QIIME 1.8.0[20]軟件中的UCLUST[21]算法對Tags在97%的相似度水平下進行聚類并獲得操作分類單元(operational taxonomic units,OTU),并基于Silva分類學數據庫對OTU進行分類學注釋。后續結果分析依靠QIIME和R等軟件,對不同組間和組內的樣本進行Alpha多樣性分析和Beta多樣性分析。
1.4 數據統計分析
實驗數據統計分析以及繪圖使用SPSS 25.0以及R 3.5.3軟件完成。采用方差分析法進行組間差異顯著性比較。
2 結果與分析
2.1 體質量變化
實驗過程中,每天記錄小鼠體質量,將連續3 d的數據合并取平均值,進而統計實驗組和對照組的體質量變化趨勢,結果發現實驗組和對照組小鼠體質量變化趨勢相近,無統計學差異。
2.2 測序數據質量評估
將HiSeq測序得到的雙端序列數據使用FLASH 1.2.7軟件拼接成一條Tags后,對拼接后的Tags使用Trimmomatic 0.33軟件進行過濾,去除過長和過短的序列,得到高質量的Tags;使用UCHIME 4.2鑒定并去除嵌合體,最終得到有效數據。實驗組和對照組中每個樣本測序后的有效序列數均在6×104條左右,表明本實驗的測序質量較高,可以用于后續的分析。
2.3 OTU分析結果
使用QIIME 1.8.0軟件中的UCLUST算法對預處理后的Tags在97%的相似度水平下進行聚類、獲得OTU。通過聚類后,實驗組和對照組的樣品聚類后的平均OTU數目均在400以上,表明樣品序列的聚類結果比較可靠,可用于下一步樣品序列的注釋,并且再一次表明本次實驗的測序質量比較高。
2.4 Alpha多樣性變化分析結果

如圖1所示,實驗組和對照組的ACE平均指數和Chao1平均指數相近,表明兩組內腸道菌群的物種數量比較相近;相比于對照組,實驗組的Simpson平均指數要高一些,而Simpson平均指數與Alpha多樣性成反比;兩組的Shannon平均指數相近。綜合這4 個指數可以發現:實驗組和對照組中腸道菌群的物種數量比較相近,實驗組菌群組成的均勻度要低于對照組。但是對于這4 個指數,組間并未表現出統計學差異。
2.5 Beta多樣性變化

圖2 小鼠腸道菌群組間多樣性比較Fig. 2 Comparison of the diversity of intestinal flora between groups
使用QIIME軟件進行Beta多樣性分析,比較不同樣品在物種多樣性方面存在的相似程度。通過樣品相關性熱圖(圖2A)和UPGMA聚類樹(圖2B)對所有樣品進行聚類分析,發現組內個體聚類較為集中統一,而組間聚類分離比較明顯,表明在元寶楓籽的干預下,實驗組和對照組小鼠的腸道菌群結構發生了比較明顯的改變。
為了進一步描述不同樣品的物種多樣性差異,對數據進行了主坐標分析(principal co-ordinates analysis,PCoA)(圖2C)。通過PCoA分析可知,實驗組和對照組數據在第一解釋因素和第二解釋因素構成的象限下發生明顯的分離(PERMANOVA:P<0.041;R2=0.0939 46),這表明實驗組和對照組的腸道菌群的物種結構組成存在顯著性差異。
考慮到實驗數據的高維情況,為了彌補PCoA的不足,進行了偏最小二乘法(partial least squares regression,PLS-DA)分析(圖2D)。圖2D結果發現實驗組數據和對照組數據依然可以發生很好的分離,因而認為在元寶楓籽作為食物的影響下,實驗組和對照組的腸道菌群的物種結構組成存在顯著性差異。
2.6 菌群結構分析
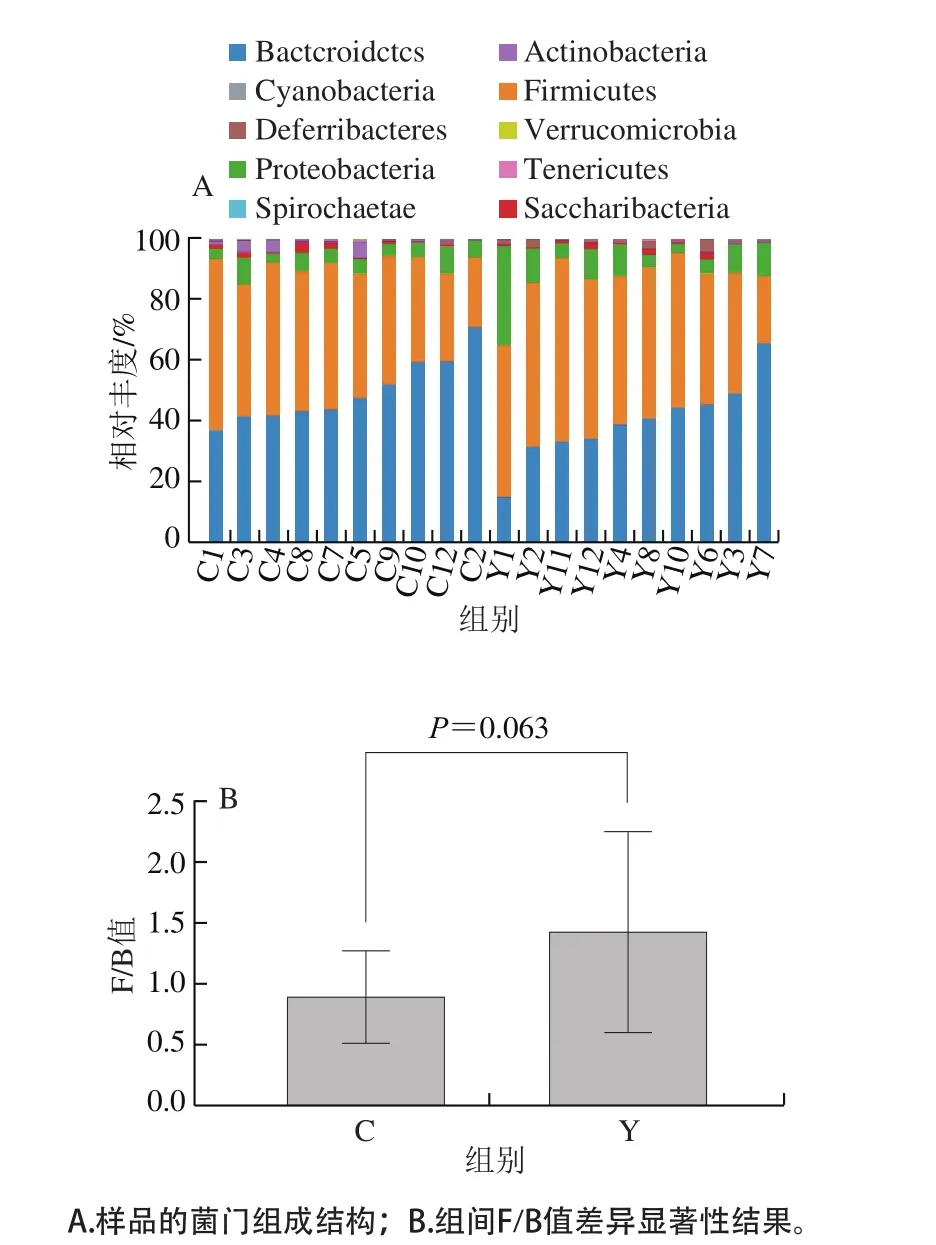
圖3 門分類學水平的細菌結構組成Fig. 3 Bacterial community structure at phylum level
將OTU的代表序列與微生物參考數據庫(Silva數據庫)進行比對,進行分類學注釋得到不同分類學水平的結果。圖3展示的是門分類學水平的分類結果,可以看出:無論是實驗組還是對照組,厚壁菌門(Firmicutes)和擬桿菌門(Bacteroidetes)均為其優勢菌門;比較組間豐度變化發現,相比于對照組,實驗組中厚壁桿菌門相對豐度上升而擬桿菌門的相對豐度下降,比較厚壁桿菌門和擬桿菌門相對豐度比值(F/B)的變化,發現實驗組F/B平均值要高于對照組(P=0.063)。
圖4是實驗組和對照組中各個樣品屬水平的分類學結果,比較組間和組內數據發現:首先是組內數據變異比較大,同一菌屬在不同樣本對象的表達豐度差異變化比較大,因此也難于直接比較組間的數據差異。對于不同組間菌屬的表達豐度差異,借助非參數檢驗統計以及機器學習等方法進行分析,這一點在后面會詳述。但是可以看出:Lactobacillus、Lachnospiraccac NK4A136 group、Bacteroides、Prevotella-1以及Alloprevotella菌屬在對照組和實驗組中均為優勢菌屬。
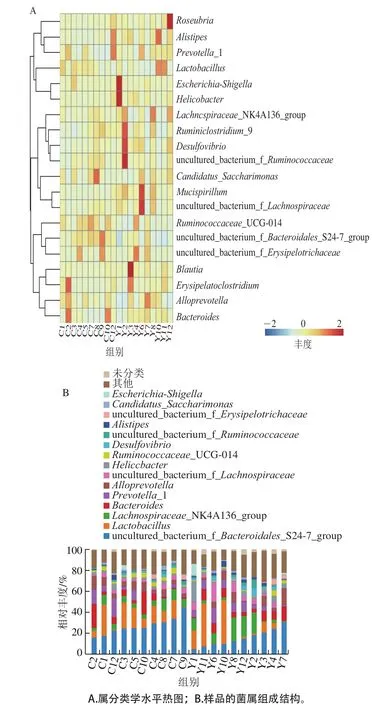
圖4 屬分類學水平的細菌結構組成Fig. 4 Bacterial community structure at genus level
2.7 組間差異物種鑒定
組間差異顯著性分析主要用于發現不同組間具有統計學差異的生物對象。圖5A為線性判別分析效應量(linear discriminant analysis effect size,LEfSe)分析結果(本次實驗分析的LAD值設為2.0)[22-23]。圖5B是隨機森林分類器篩選后門水平的評分結果,圖5C是隨機森林分類器篩選后屬分類水平的評分結果。將LEfSe和隨機森林的分析結果結合,篩選出重要性評分高且存在組間顯著性差異的對象,認為這些是存在于組間有意義的差異物種(圖6、7)。


圖5 小鼠腸道菌群組間差異顯著性分析Fig. 5 Significance analysis of difference in intestinal flora between groups
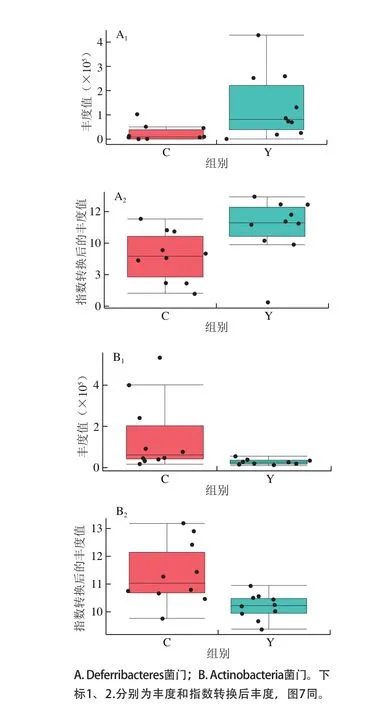
圖6 小鼠腸道菌群組間門水平差異物種Fig. 6 Difference in intestinal flora between groups at phylum level
圖6 為門分類學水平的組間差異對象結果,分別為脫鐵桿菌門(Deferribacteres)和放線桿菌門(Actinobacteria);其中,實驗組中脫鐵桿菌門的平均豐度要高于對照組,而放線桿菌門的平均豐度要低于對照組。由于門分類學水平下包含許多菌屬,且不同菌屬對宿主的影響不盡相同,所以盡管發現了這兩個差異的門水平對象,卻難以直接描述這種差異變化對宿主的影響。但是這種差異現象是應該被注意到的,因為它往往表征的是一種趨勢性變化,而這種變化趨勢往往具有比較好的表現性和重復性,易于不同的實驗結果之間進行比較和解讀。



圖7 小鼠腸道菌群組間屬水平差異物種Fig. 7 Difference in intestinal flora between groups at genus level
圖7 為屬分類學水平組間差異種屬,實驗組中Mucispirillum、Rikenella和Anaerotuncus的平均豐度要高于對照組,而Staphylococcus、Acinetobacter和Corynebacterium_1的平均豐度要低于對照組。Mucispirillum被報道作為結腸黏膜層的健康標記物,它在病原微生物感染早期表現為豐度下降,在恢復期(病原體被清除之后)豐度回到正常水平;Staphylococcus和Acinetobacter均是被廣泛報道的條件致病菌,當機體抵抗力降低時易引起機體感染,是造成醫院內感染的重要機會致病菌之一,在2017年世界衛生組織公布的十二大耐藥性細菌中,Acinetobacter赫然排名在級別1——嚴重耐藥性的第一名。
2.8 共性網絡分析
共性網絡分析通過將不同屬性的對象抽象為節點,并用連接來展示不同節點之間的關系,通過量化以節點和連接為組件的網絡結構指數,從而能夠在統一的框架下尋找復雜系統的共性。在紛雜的共性網絡里面,很難確定某幾個節點之間確切的關系,但是可以借助其尋找聯系緊密的特定群組或者反應群體的結構組成特征。
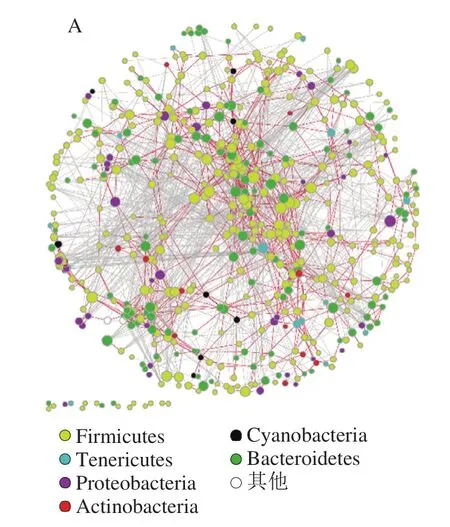

圖8 小鼠腸道菌群網絡分析圖Fig. 8 Network analysis of intestinal flora in mice
由圖8可知,無論是對照組還是實驗組,厚壁菌門和擬桿菌門構成了主要的節點聚合中心,但是對照組中厚壁菌門和擬桿菌門的節點分布相對比較分散均一,而實驗組中擬桿菌門和厚壁桿菌門呈現出兩極分離的趨勢,這提示實驗組中門水平的物種結構關系已經發生了改變。
2.9 腸型的鑒定
按照屬水平聚類分析方法劃分腸型,根據腸型分類結果發現:在對照組中20%的個體屬于1類腸型(ET B),80%的個體屬于3類腸型(ET F);在實驗組中,40%的個體屬于1類腸型(ET B),60%的個體屬于3類腸型(ET F)。
3 討 論
神經酸在大腦和神經組織中含量較高,常作為腦中白質的標志物,神經酸因為能夠修復受損的神經元并促進神經細胞再生而備受關注[2-3]。神經酸以往的來源主要依靠鯊魚,隨著國際社會禁捕鯊魚,各國政府和研究機構開始把方向轉向從植物中提取神經酸[24]。歐乞鍼從蒜頭果樹的種仁油中發現了神經酸,其質量分數高達67%[25]。但因蒜頭果樹屬國家二級保護植物,其開發利用受到一定限制。王性炎教授等從槭樹喬木元寶楓的籽中也發現了神經酸,其在元寶楓油中的含量在5%~6%[26]。元寶楓是中國含神經酸木本植物的一個特有樹種,目前仍是提取神經酸的主體資源[27]。研究發現,元寶楓油對大腸桿菌、枯草芽孢桿菌和黃曲霉菌具有明顯的抗菌作用[3]。
為了研究飲食對腸道菌群的影響,可以通過長期飲食習慣或飲食干預方法分別了解其長期及短期影響[28-32]。飲食作為腸道菌群的一種生態驅動力,使腸道菌群能適應其暴露的飲食結構,有研究發現:腸道菌群能夠反映宿主的飲食習慣、腸道結構和生理狀況[13-14]。本實驗將元寶楓籽作為食物飼喂小鼠,飼喂2 周后,檢測小鼠腸道菌群的組成結構的變化。根據高通量測序的結果,發現實驗組和對照組之間Beta多樣性發生了顯著性的差異,這表明在通過元寶楓籽進行飲食干預后,實驗組的腸道菌群組成發生了顯著性的改變,那么如何去評價這種改變呢?從兩個方面去嘗試分析:物種分類學上的差異改變,即腸道菌群的物種結構組成;腸道菌群功能組成上的變化。
首先進行LEfSe分析,LEfSe分析是基于非參數檢驗方法確定不同組間的差異對象,然后進行對象的亞類判定,建立線性判別分析模型,最終得到特定分類水平下的差異物種列表及差異對象的效應值。這種方法是基于統計學的顯著性差異來篩選對象的,那是否統計學差異就等同于生物學意義上的差異?為此引入了隨機森林分類器,隨機森林是基于決策樹的集成學習算法,通過對每個基本分類器的分類結果進行組合,來決定待分類樣本的歸屬類別[33]。通過對每一分類學水平內包含的對象進行重要性評分,按照從高到低排序,決定哪些對象是重要的,值得注意的是:這里的重要性不等同于統計學上的差異顯著性。通過結合這兩種分析篩選方法,在門水平和屬水平分別找出了組間差異的物種,在屬分類學水平的對象中,實驗組中Mucispirillu的平均豐度要高于對照組。正如結果分析部分中提到的,Mucispirillum是一種已經被報道的宿主健康狀態下的標志菌屬,其豐度變化常與腸道炎癥的發展進程相關。Mucispirillum常在結腸遠端的外層腸黏膜表面形成集群,在病原菌屬進攻損傷腸黏膜的時候,Mucispirillum的豐度往往會下降;黏膜層完整的恢復開始于病原體清除后一段時間內,于此同時,Mucispirillum的豐度也會逐漸恢復到健康水平,因此,Mucispirillum能作為遠端結腸表層黏膜層健康水平的標記物,用于表征潛在的炎性結腸炎;其他細菌類型標志物,如梭菌屬和乳桿菌屬,在病原體消失后這些標志物豐度也會增加。于此同時,實驗中Staphylococcus和Acinetobacter的平均豐度要低于對照組,這兩種菌屬是已經被廣泛報道的機會性致病菌,Acinetobacter更是公認的嚴重耐藥菌屬。盡管這兩種菌屬在正常宿主機體內不發病,但是當宿主免疫水平降低或者遭受其他病理性因素影響時,這些菌屬往往會成為繼發感染的菌屬[34-35]。
腸道菌群行使多種代謝分解功能,每個功能的實現往往是多種細菌相互協同拮抗來實現的,因此,在考慮不同分類學水平上單個差異物種的同時,更應該去分析不同因素干預下菌群功能組成上的變化[36]。群體分型是一種有助于更好地理解人類身心健康等復雜生物學問題的有效方法,將這種方法應用于腸道微生物中,可以將不同的群落組成定義為腸型,腸型的意義在哪里呢?個體間微生物群落存在著巨大的差異,同時微生物群落變化還表現出不同的階段性,目前,在大群體和跨地理研究中,還很難簡單地描述如此復雜的生物現象,部分原因是有些菌屬在不同個體間表現為單峰,而另外一些菌屬又表現出雙峰或者更復雜的分布模式,因此確定微生物群落組成的結構模式及其背后的組裝機制是非常必要的[37]。對每一種腸型的特性研究,發現共存的微生物網絡中心存在一種驅動類群,即這一類微生物與給定的腸型最相關。在人類的微生物組學研究中,一種常用的腸型劃分方法是基于腸道菌群的屬分類學水平分類進行劃分,大致可以分為3 個腸型:腸型1(ET B)、腸型2(ET P)和腸型3(ET F)[38]。盡管動物的腸道菌群與人類不同,但是在控制實驗中,聚類方法可以將小鼠的腸道菌群劃分成明顯的集群,這表明,當沒有外部因素影響微生物群落時,更容易出現聚集的群落狀態[38]。因此,腸型并不是人體腸道菌群特有的,動物中同樣存在這些現象。將數據上傳到腸型在線分類器主頁(http://enterotypes.org/),得到腸型分類結果,發現實驗組中1類腸型的比例上升,3類腸型的比例下降。這個差異變化,在圖8中也得到了驗證。通過飼喂元寶楓籽影響了實驗組中小鼠群體的腸型結構組成,這為其他研究人員做相關平行研究提供了參比數據。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
音樂探索(2022年2期)2022-05-30 21:01:37
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
山東女子學院學報(2014年6期)2014-03-01 02:24:55
