白樺MYB基因響應激素及鹽旱處理的表達研究
2020-07-14 01:00:18劉佳欣劉慧子石晶靜
植物研究 2020年5期
關鍵詞:分析
劉佳欣 劉慧子 石晶靜 于 穎 王 超
(林木遺傳育種國家重點實驗室,東北林業大學,哈爾濱 150040)
已知植物莖的伸長生長受多種內源激素的調控,如生長素吲哚乙酸(IAA)、赤霉素(GA)、油菜素內酯(brassinosteroid,BR)和乙烯(ethylene,ETH)等都促進莖的伸長生長[9]。而外源激素的施加能夠改變內源激素的含量,并影響莖生長發育。IAA、GA3和IBA浸種的兩兩交互作用對云南松胚軸和根長具有極顯著或顯著的差異影響[10]。外源激素GA3與ABA對烤煙莖尖處內源激素GA、ABA、IAA含量以及株高、節間距伸長產生影響[11]。噴施外源激素GA3可調節矮稈基因型煙草莖尖中各內源激素的含量,與正常株高基因型“K326”的各激素含量相接近,從而促進莖生長[12]。從外界信號到性狀改變的過程是如何通過一系列的分子響應完成的?哪些基因參與了其調控的分子途徑?分子機制如何?是研究激素調控植物生長發育的重要研究內容之一。
MYB轉錄因子是植物轉錄因子家族中最大的家族之一[1],不僅在植物生長發育調控中調控生長發育的節奏以及器官的形成,而且可以響應非生物脅迫誘導從而影響代謝產物的形成與激素信號傳導等[2~4]。轉錄因子的主要作用方式時通過與下游啟動子上的順式作用原件相結合的方式來調控靶細胞基因的表達從而形成同源、異源二聚體,也可與其他蛋白互作成為某種活化形式參與JA(jasmonate acid)、SA(salicylic acid)、ABA(abscisic acid)等信號傳導途徑,形成基因表達的調控網絡。遺傳學分析發現ABA誘導花青素合成部分依賴于MBW復合體中的核心轉錄因子,如TTG1、TT8及MYB75等[5]。水曲柳FmMYB5基因對IAA(indole-3-acetic acid)、ABA、GA3(gibberellin)、JA和SA 5種激素誘導都存在顯著響應且都是正調控作用[6]。核桃JrEFM1可被ABA、JA、SA處理不同程度地誘導[7]。擬南芥AtMYB51受到水楊酸強烈誘導,AtMYB34則對茉莉酸甲酯最為敏感,處理后表達水平顯著提高[8]。這些研究說明植物MYB轉錄因子廣泛參與外源激素調控的植物發育中。種子萌發到下胚軸和胚根的形成是植物發育、形態建成的重要生物學過程,受到不同激素的調控,研究該過程響應激素處理的MYB轉錄因子表達模式,對研究木本植物發育及形態建成的分子調控具有一定的研究價值。
白樺(Betulaplatyphylla)是我國北方重要的經濟和園林綠化樹種之一[13],其具有材質潔白細膩,抗旱耐瘠薄的優良特性,適于做紙漿材,單板材和觀賞樹木。是研究林木重要性狀形成分子機制的良好材料,也是重要的改良樹種。本研究以白樺為材料,選擇MeJA(Methyl jasmonate)、ETH、ABA和KT對野生型白樺種子進行噴灑處理,比較不同處理條件下白樺苗與對照苗胚軸及根發育情況;利用實時熒光定量PCR對經過不同激素噴灑、NaCl和Mannitol處理下的白樺BpMYBs基因的表達量進行分析。分析其響應激素處理的表達模式,為了進一步明確BpMYB家族基因的信號調控及功能提供依據。
1 材料與方法
1.1 實驗材料
將野生型白樺種子用流水沖泡2 d,然后種到人工營養土壤(腐殖質∶蛭石∶珍珠巖3∶1∶1)中覆膜保持濕度(3 d后將膜揭掉),放置于生化培養箱內,第二天開始進行激素處理,分別用10 μmol·L-1的MeJA、100 μmol·L-1的ETH、20 μmol·L-1的ABA和50 μmol·L-1的KT對白樺種子進行噴灑處理,同時以噴灑清水處理作為對照,期間每隔1 d噴灑1次。待白樺苗長到4周大小時,將不同激素處理后的白樺苗完整的從土壤中取出,用清水將根部的土壤洗凈,平鋪在干凈的玻璃平板上進行觀察,并照相。用于基因表達分析的材料迅速用液氮處理,放于-80℃冰箱內保存備用。取8周大小的白樺苗,分別由200 mmol·L-1NaCl和100 mmol·L-1Mannitol處理6、12、24和48 h,用水處理白樺苗作對照進行基因表達分析。
1.2 RNA提取及cDNA合成
使用CTAB法提取白樺中的總RNA,利用反轉錄試劑盒(Takara,日本)進行一鏈cDNA的合成。反應體系為20 μL:2×TS Reaction Mix10 μL,TransScript? RT/RI Enzyme Mix 1 μL,Anchored Oligo(dT)18 Primer(0.5 μg·μL-1) 1 μL,Total RNA 50 ng至5 μg,gDNA Remove 1 μL,補充RNase-Free Water至20 μL。反應條件:在PCR儀(Biometra)中,42℃條件下,反應30 min;85℃條件下,反應5 s。實驗結束后,將cDNA于-20℃冰箱保存備用。
1.3 基因克隆及生物信息學分析
對白樺應拉木轉錄組數據進行分析,篩選鑒定出轉錄組中的BpMYB基因序列,利用NCBI的ORF finder程序進行編碼區分析并鑒定其完整的CDS序列。以野生型白樺cDNA為模板,根據白樺MYB基因CDS序列設計特異引物(見附表1)進行RT-PCR克隆。克隆產物送華大(北京)公司進行測序。利用MEGA(V.6)對白樺MYB基因編碼的蛋白與擬南芥已知MYB蛋白序列進行系統進化分析。
1.4 熒光定量RT-PCR分析
根據白樺MYB基因CDS序列,選取200 bp左右長度序列設計特異引物(見表1)進行熒光定量PCR克隆。分別以不同脅迫處理和水對照的白樺苗cDNA為模板進行熒光定量PCR試驗,其中以Tubulin(GenBank number:FG067376)和Ubiquitin(GenBank number:FG065618)為內參引物,設置3次重復(包括3次生物學重復以及3次技術重復)。實時熒光定量RT-PCR反應體系20 μL,cDNA 2.0 μL,Primer F(10 μmol·L-1) 1.0 μL,Primer(10 μmol·L-1) 1.0 μL,SYBR Green Realtime PCR Master mix 10.0 μL,補充ddH2O至總體積20 μL。實時熒光定量PCR反應程序:94℃ 30 s;94℃ 12 s,58℃ 15 s,72℃ 40 s,79℃讀板1 s,共45個循環。反應結束后,運用ΔΔCt方法對基因的相對表達量進行定量數據分析。

表1 BpMYB家族基因定量引物序列
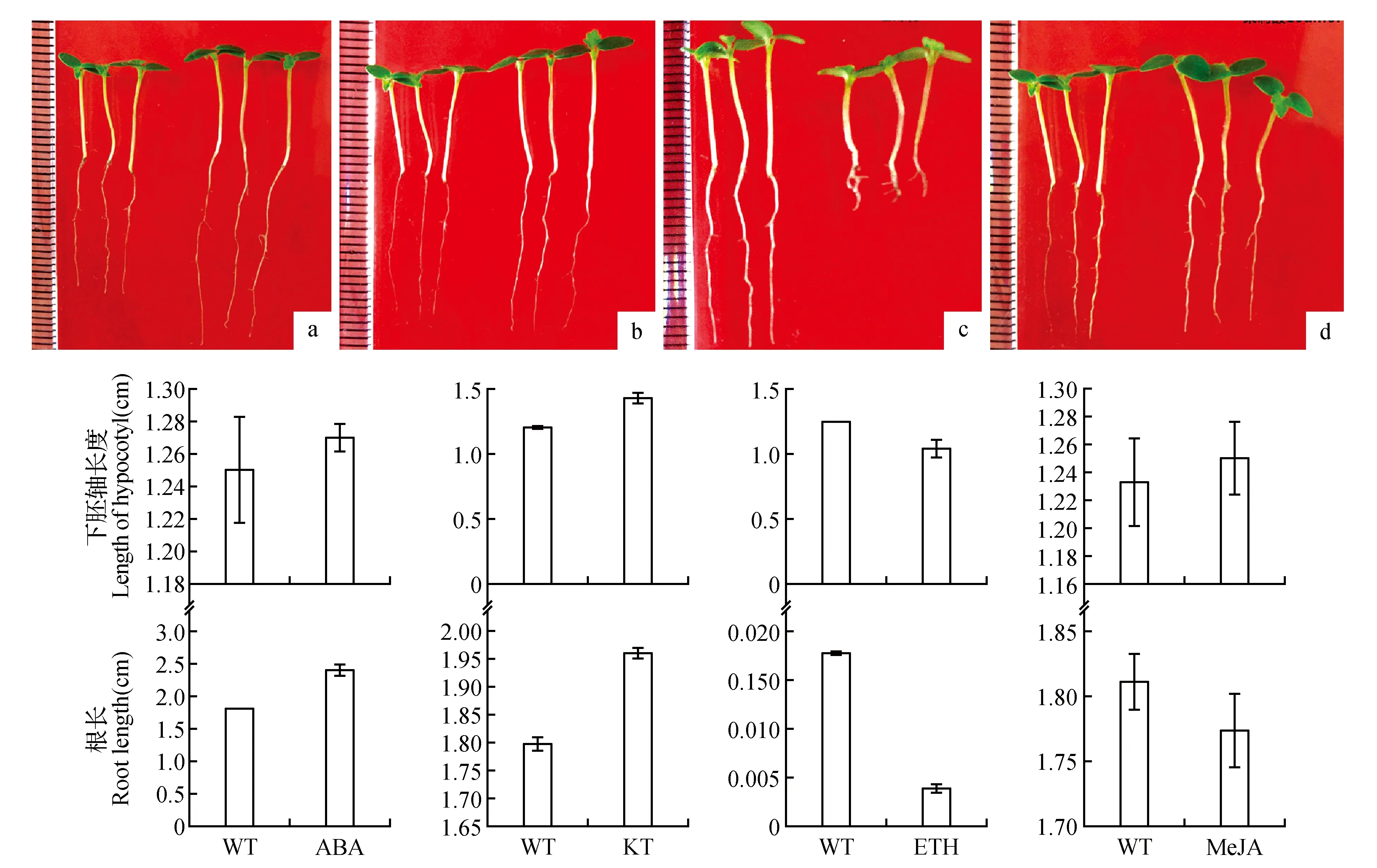
圖1 不同激素處理的白樺幼苗表型分析 a.20 μmol·L-1 ABA;b.50 μmol·L-1 KT;c.100 μmol·L-1 ETH;d.10 μmol·L-1 MeJAFig.1 Phenotype of birch seedlings treated with different hormones a.20 μmol·L-1 ABA;b.50 μmol·L-1 KT;c.100 μmol·L-1 ETH;d.10 μmol·L-1 MeJA
2 結果與分析
2.1 激素處理下白樺表型觀察
對白樺種子分別用10 μmol·L-1的MeJA、100 μmol·L-1的ETH、20 μmol·L-1的ABA和50 μmol·L-1的KT進行噴灑處理,4周后觀測其生長性狀。從苗的表型上來看(見圖1),經過ETH噴灑處理的白樺苗變化最為明顯,它的下胚軸比未處理的白樺下胚軸短,根長顯著減小,約是未處理白樺苗的20%;經過ABA處理的白樺苗,下胚軸比未處理的白樺苗的下胚軸稍長,根長顯著增加,比未處理的根長30%;經過KT處理的白樺苗的下胚軸和根均比未處理的白樺苗的下胚軸和根長;經過茉莉酸處理的白樺苗下胚軸和根比對照稍長。
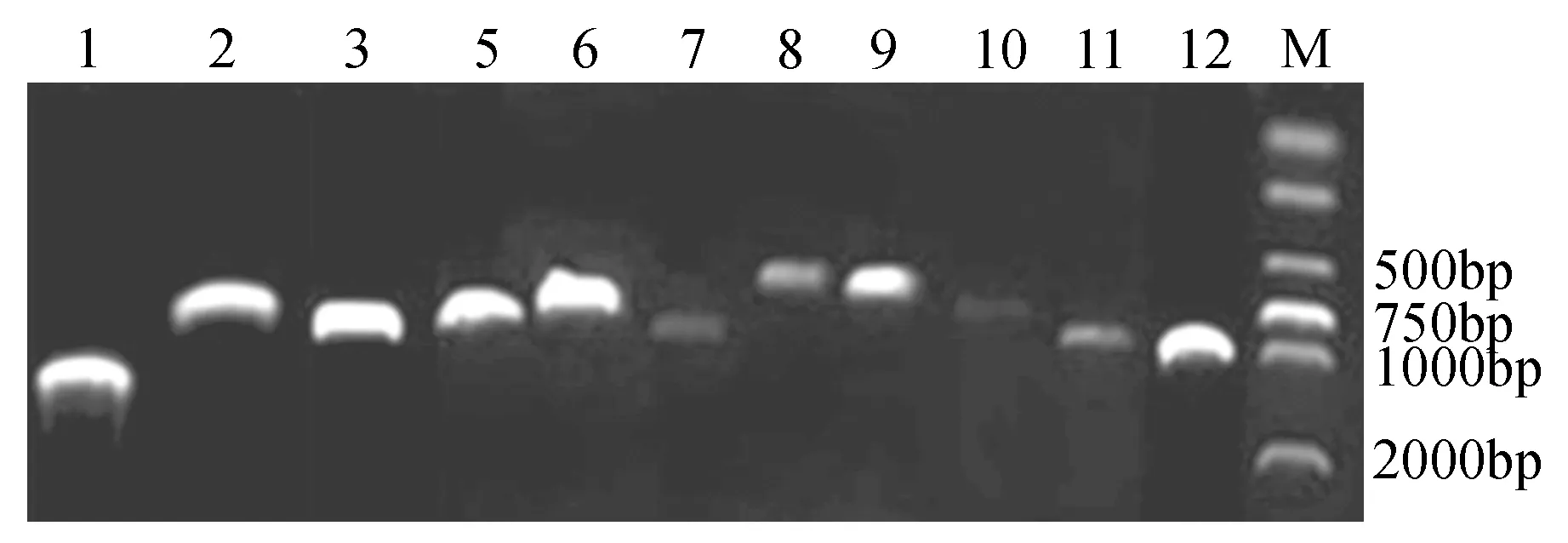
圖2 白樺MYB基因RT-PCR克隆Fig.2 RT-PCR cloning of birch MYB gene
2.2 白樺MYBs基因克隆及系統進化分析
通過對所選白樺MYB基因進行RT-PCR,成功擴增出11條目的片段,將這11條MYB目的片段利用膠回收試劑盒(Omega)進行回收,用PMD18-T載體進行連接并且成功轉化大腸桿菌,從中隨機挑取陽性菌斑,利用11條MYB基因的特異性引物進行菌液PCR,PCR結果顯示目的基因條帶正確(見圖2),將菌液送至公司測序,測序結果與基因序列比對一致,成功克隆得到11條具有完整ORF的白樺MYB序列,這些基因編碼區堿基數在624~1 203 bp。根據克隆順序命名為BpMYB1、2、3、5、6、7、8、9、10、11和BpMYB12。本研究增加了一條研究前期克隆的BpMYB4,共12條。將這12條基因編碼的蛋白與107條擬南芥MYB家族基因進行系統進化分析,結果顯示可以分為6大組,其中BpMYB1和BpMYB6聚為一組,BpMYB3、10、12聚為一組,BpMYB2、4、5和BpMYB8蛋白序列相似性最高,BpMYB7、9和BpMYB11進化關系較近。BpMYB1與AtMYB61、AtMYB50進化關系比較近,屬于R2R3類型的家族成員;BpMYB12與編碼葉軸MYB結構域蛋白AtMYB91進化關系比較近;BpMYB4l與AtMYB4進化關系比較近;BpMYB5與種子萌發過程中的原花色素(proanthocyanidin)相關基因AtMYB123進化關系比較近;BpMYB7與影響氣孔開關的AtMYB60進化關系比較近;BpMYB11與AtMYB30進化關系比較近;BpMYB9與AtMYB84進化關系比較近(見圖3)。
2.3 白樺MYBs基因響應激素處理的表達模式
以不同激素、NaCl和甘露醇處理白樺苗的cDNA為模板,以同期未經處理的白樺苗為對照,利用實時熒光定量PCR技術對白樺中BpMYBs基因的表達量進行分析。
經過20 μmol·L-1ABA的處理后,只有BpMYB4、8上調表達,但差異不顯著,其他BpMYB基因全都下調表達,除BpMYB11和BpMYB12下調表達差異非常顯著外,其他基因下調的倍數較茉莉酸和乙烯利處理下調的少。經過50 μmol·L-1KT處理后,除了BpMYB11依然處于下調表達以外,其他的BpMYB基因全部呈現出上調表達,其中BpMYB2、8上調倍數最高。經過100 μmol·L-1ETH處理后,所有基因的表達模式出現較為明顯的統一趨勢,全部都呈現出下調表達,除了BpMYB2與BpMYB8下調表達量差異不顯著外,其他的BpMYB基因全都高度下調表達。其中BpMYB11的下調表達量差異最大。經過10 μmol·L-1MeJA處理后, 除了BpMYB2和BpMYB8上調表達
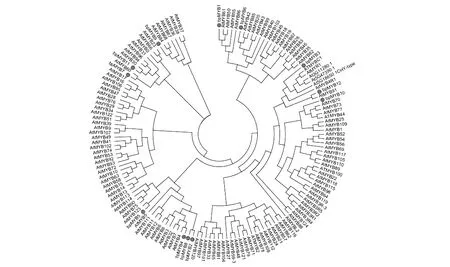
圖3 白樺與擬南芥MYB蛋白系統進化分析 擬南芥MYB家族蛋白序列來源于TAIR網Fig.3 Phylogenetic analysis of MYB protein in birch and Arabidopsis The protein sequence of MYB family in Arabidopsis is derived from TAIR website
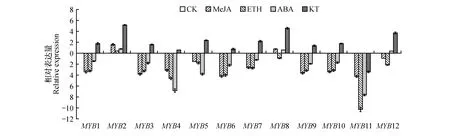
圖4 激素處理下白樺MYBs基因的表達模式分析Fig.4 Analysis of expression pattern of MYBs gene in birch under hormone treatment

圖5 200 mmol·L-1 NaCl處理下白樺MYBs基因的表達模式分析Fig.5 Analysis of the expression pattern of MYBs genes in white birch under 200 mmol·L-1 NaCl treatment
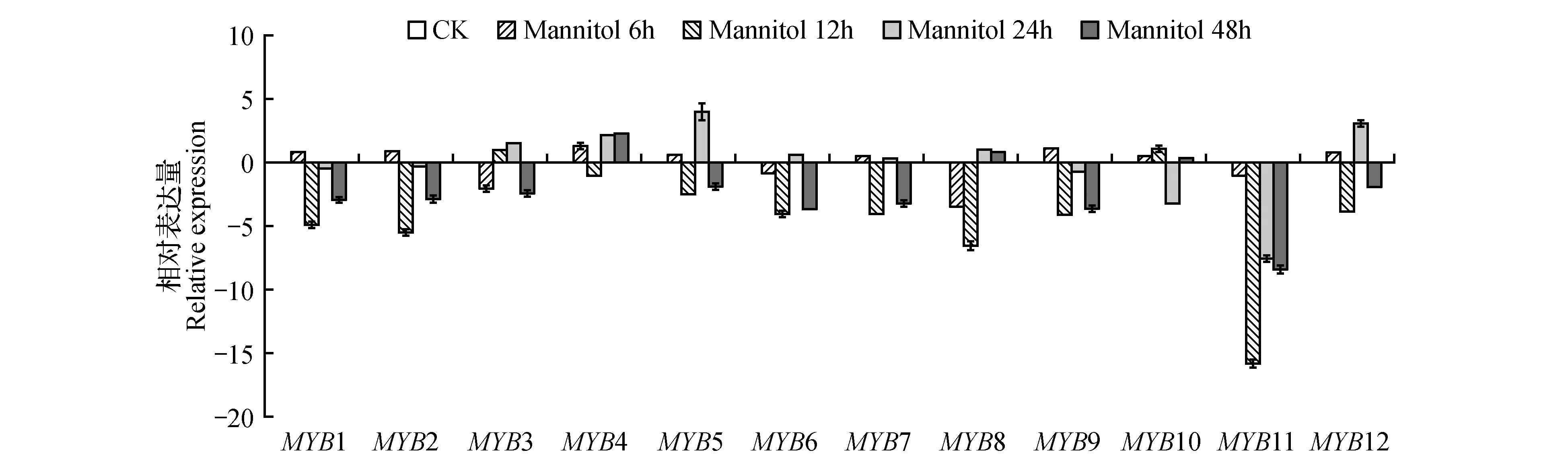
圖6 100 mmol·L-1 Mannitol處理下白樺MYB基因的表達模式分析Fig.6 Analysis of the expression pattern of MYBs gene in birch under 100 mmol·L-1 mannitol treatment
外,其余基因全部為下調表達,其中BpMYB5和BpMYB12的下調表達量相對較少,其他基因都呈現出高度下調表達趨勢。
經過高濃度NaCl脅迫處理后,BpMYB4基因最先響應脅迫誘導。BpMYB9、10和BpMYB12表達差異不明顯。BpMYB11主要表現出顯著的下調表達模式,表達量最高可下調317倍。其他大部分基因表現出先下調后上調的表達模式,在48 h表達上升,包括BpMYB1、2、3、6和BpMYB7。BpMYB8主要在處理48 h被誘導。
在甘露醇脅迫處理6 h時,除了BpMYB3、6、8、11外其余基因均為上調表達,但表達量不高,最高的也僅僅為3.1倍;在甘露醇脅迫處理12 h的時候,只有BpMYB3輕度上調表達,其余MYB基因全部都呈下調表達模式,且為高度下調表達;在甘露醇脅迫處理24 h的時候,只有BpMYB11呈高度下調表達,雖然BpMYB1、2、9也呈下調表達,但表達差異較小,其余基因均為上調表達;經過甘露醇脅迫處理48 h的時候,除了BpMYB4、8、10為上調表達外,其余基因均為下調表達,且為高度下調表達,但是,整體下調表達的強度比脅迫12 h的下調表達強度弱。BpMYB11在脅迫12 h后,都呈現出顯著的高度下調表達。
3 討論
在不同激素處理下,白樺4周幼苗表現出了不同的生長表型,ABA和KT處理,能夠促進白樺幼苗下胚軸及根的伸長,而ETH處理,顯著抑制白樺幼苗下胚軸及根的伸長,但是促進了下胚軸的增粗及側根的生長。MeJA處理效果不明顯。有研究表明高濃度(10-6~10-4mol·L-1)的ABA和MeJA均抑制花生根和下胚軸的生長;低濃度(10-7mol·L-1)的ABA作用不明顯,MeJA在低濃度下對生長表現出明顯的促進效應[14]。高濃度(10 μmol·L-1)ABA做為脅迫因素在短時間內對2種藜科抗逆植物的種子萌發和幼苗生長均有促進效應,而長時間作用則使2種植物產生不同的適應策略,這種差異可能由于2種種子大小不同所導致[15]。較低溫度下(5、10℃),植物材料的發芽率、根長、芽長均隨著ABA濃度的提高而降低,20℃時,1、5 mg·L-1的ABA處理對種子萌發均有促進作用[16]。本研究10 μmol·L-1ABA和50 μmol·L-1KT處理對白樺下胚軸和根生長起到促進作用,可能是因為白樺種子的特異性及早期短時間處理造成的。植物激素乙烯(ETH)調節植物生命周期許多方面,如種子萌發、根毛發育、根瘤形成、花衰老、凋謝和果實的成熟。ETH對胚軸伸長的作用依賴幼苗生長在光下或暗中,暗中ETH及ETH合成前體AC(C1-氨基環丙烷1-羧酸)抑制胚軸伸長[17]。本研究白樺種子播種于土壤中進行ETH處理,起到了抑制胚軸伸長的作用。
從整體趨勢來看,本研究的白樺MYB基因在4種不同激素的處理下的表達模式的整體趨勢基本一致:在10 μmol·L-1MeJA、100 μmol·L-1ETH以及20 μmol·L-1ABA的噴灑處理后,基因大部分都處于下調表達,但是其表達量有所差異,其中在脫落酸處理下的下調表達量較其他兩個激素處理后的下調表達量低,而經過乙烯處理后的差異表達量最高,呈高度下調表達模式,這種表達模式暗示了在ETH處理下,BpMYB基因的下調表達,在白樺幼苗下胚軸及根的生長抑制中起作用。在50 μmol·L-1KT的噴灑處理后,基因的表達量全部上調,說明白樺應答KT誘導的BpMYBs基因呈現為正向的調控模式,有研究表明,過表達擬南芥MYBH基因能夠促進擬南芥下胚軸的伸長[19~20],本研究中這些上調的BpMYB基因可能在KT處理下白樺幼苗下胚軸及根的伸長中起作用。本研究BpMYBs基因對于不同外源激素信號響應不同,說明它們在不同激素信號途徑中起作用。其中有兩個表達模式較為特別的基因,一個是BpMYB2,在4種不同激素的處理下都處于一個上調表達的模式,但表達量有所差異,在激動素(KT)的處理下BpMYB2的表達量高度上調,可能與KT處理促進下胚軸及根發育相關;BpMYB2與BpMYB8在激素處理下的表達模式相似,這可能與它們系統進化分析聚為一組相關。另外一個基因是BpMYB11,它與BpMYB2正好相反,在4種不同激素的處理及鹽和旱脅迫處理下都處于一個下調表達的模式,而且均處于一個高度下調表達的趨勢,尤其是在100 μmol·L-1ETH噴灑處理后下調表達量極高,說明該基因受到外源激素信號的抑制。MYB轉錄因子在植物抗逆脅迫過程中根據是否依賴ABA信號途徑可將其分為依賴于ABA和獨立于ABA信號通路兩類[14]。本研究發現用20 μmol·L-1脫落酸(ABA)的噴灑處理后,BpMYB11、12表現為強烈的下調表達,說明這兩個基因可能依賴于ABA信號通路。在高濃度NaCl脅迫處理情況下,本研究大部分BpMYBs基因在脅迫處理初期下調表達,而在處理48 h被顯著誘導上調表達;BpMYB4、5和BpMYB12在鹽和旱脅迫處理早期即被誘導,并持續上調表達,可能和之前的信號傳導途徑有關[18]。總體上,系統進化分析聚為一組的基因有著相似的表達模式。
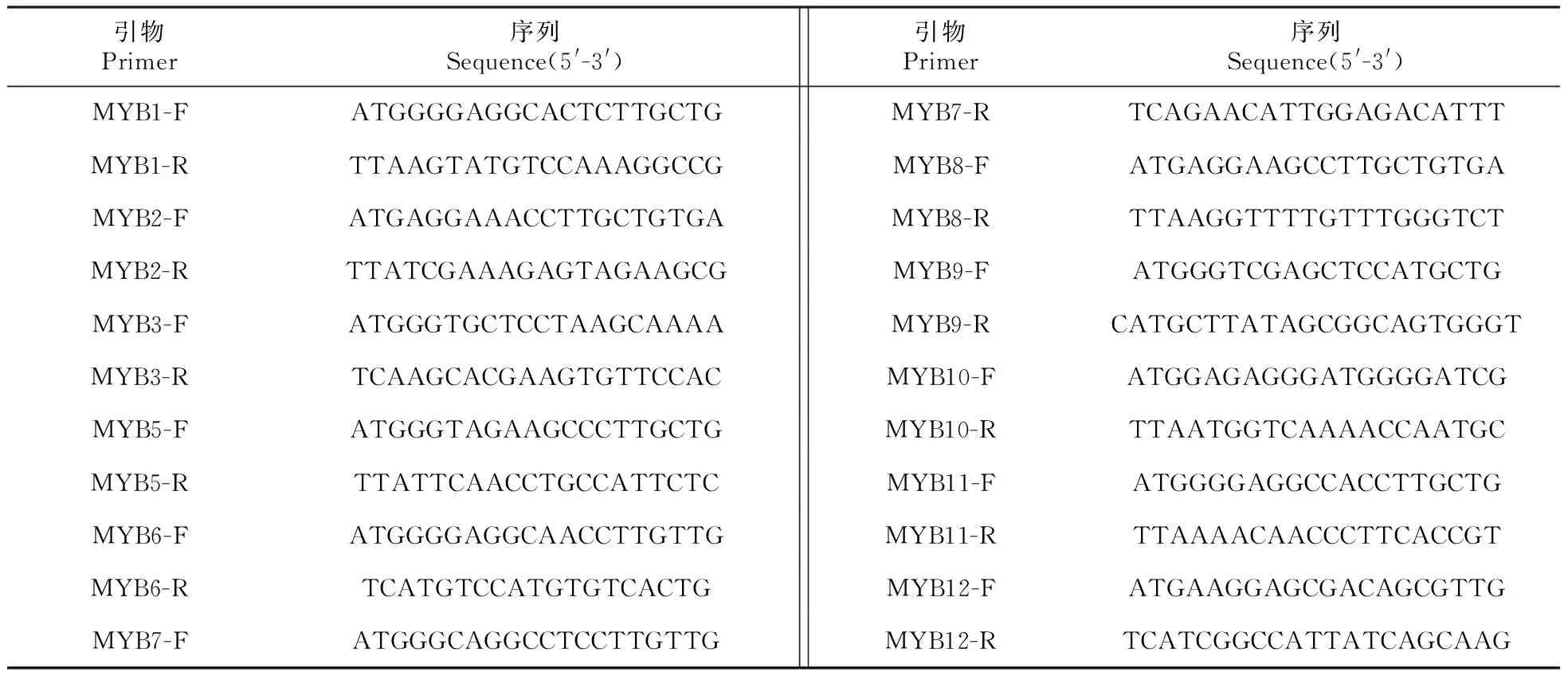
附表1 BpMYB家族基因克隆的引物序列
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
