白樺BpJMJ18基因啟動子克隆及表達分析
2020-07-14 01:00:18王萬奇齊婉竹趙秋爽付鵬躍曲冠證趙曦陽
植物研究 2020年5期
關鍵詞:分析
王萬奇 齊婉竹 趙秋爽 曾 棟 劉 軼 付鵬躍 曲冠證 趙曦陽*
(1.東北林業大學林木遺傳育種國家重點實驗室,哈爾濱 150040; 2.吉林師范大學生命科學學院,四平 136000; 3.北華大學林學院,吉林 132013)
JMJ類蛋白是通過組蛋白的去甲基化以及與其他染色質修飾互作等方式,參與許多基因表達和染色質活動的調控,進一步影響生物生長發育、代謝和環境響應等生物學過程[1]。JMJ類蛋白成員數目眾多,按照JmjC結構域的序列分析,可以將其分為7大類[2]:JARID類,具有H3K4去甲基化活性[3];JMJD2亞家族,是H3K9和H3K36的去甲基化酶[4];JHDM2類,能夠去除H3K9me2[5];JMJD6類,只含有JmjC結構域,能夠去除精氨酸甲基化[6];UTX/JMJD3和JHDM1是動物所特有的兩類蛋白[7];而植物所特有的一類蛋白中,JMJ14成員是H3K4去甲基化酶基因,該基因在調控擬南芥開花中扮演重要的角色,能夠與兩個NAC家族轉錄因子互作來影響植株的生長發育[8]。此外,JmjC結構域屬于Cupin超家族。JmjC結構域蛋白可能是催化組蛋白修飾的蛋白質羥化酶。同時,通過對本研究的JMJ18蛋白序列分析發現,該序列除含有FRY、FYRC、JmjC、JmjN結構域外,也含有C5HC2類的鋅指結構域,而該結構域極有可能與特定序列的DNA結合,發揮類似轉錄因子的作用。
JMJ蛋白功能最初是在動物中發現的,它能影響小鼠(Mus musculus)的大腦和心臟發育[9~10],而植物中的JMJ蛋白雖然沒有動物當中研究的那么廣泛且深入,但就前人已有的研究發現JMJ蛋白能夠參與植物器官發育、生殖過程、激素應答及DNA甲基化等多個生物學過程[11]。植物中最早發現的具有組蛋白去甲基化酶活性的是IBM1(即JMJ25),它能影響植株的許多表型[12];且到目前為止,JMJ蛋白的研究多數集中在擬南芥(Arabidopsisthaliana)和水稻(Oryzasativa)等草本植物中,例如:JHDM家族的JMJ12基因與擬南芥的花期有密切的關系[13];水稻中的組蛋白去甲基化酶JMJ705可以通過提高抗性基因的表達來去除H3K27的甲基化,進而提高水稻對白葉枯病的抗性[14];JMJ706基因突變后能夠引起水稻花器官發育異常,進而影響花器官的形態建成[2];OsJMJ718基因在水稻的生殖發育階段也能夠呈現不同的動態表達模式[15];JMJ703突變后能夠影響植物細胞分裂,從而使莖干和幼穗發育受到影響[16]。綜上所述,不同植物中JMJ基因功能不盡相同,但均表明該基因與植物生長發育密切相關。
白樺(Betulaplatyphylla)是樺木科(Betulaceace)樹種落葉喬木,是我國東北地區重要的闊葉落葉林及針葉闊葉混交林中常見樹種,其具有生長快、木材細膩、適應性強等特點,是東北地區木材生產的重要材料,具有極高的研究和利用價值[17]。由于白樺全基因組已被測定[18],很多基因功能的開發與驗證均以白樺為基礎材料,例如:楊洋以白樺為實驗材料來研究BpTCP基因在植物生長發育過程中發揮的作用,進而揭示BpTCP基因的生物學功能[19];李蕾蕾等[20]為了解GT14基因在生長發育過程中的功能及對環境的適應機制,選取白樺為材料,對該基因的啟動子進行了相關的研究;同時,王宇航[21]也以白樺的鋅指蛋白轉錄因子基因為研究對象,對基因的表達和功能進行了初步研究,但到目前為止對JMJ類基因在木本植物中的功能研究較少。本研究以白樺為研究對象,對BpJMJ18基因的啟動子進行克隆,通過構建植物表達載體,并用農桿菌介導的瞬時轉化法及原生質體瞬時轉化,研究其BpJMJ18基因的組織表達模式以及BpJMJ18蛋白定位區域,為后續進一步研究JMJ18基因在植物生長發育方面的功能奠定重要基礎。
1 材料與方法
1.1 實驗材料
于2016年4月在東北林業大學林木遺傳育種白樺強化育種基地,采集4年生的白樺優樹嫩葉建立組培體系,并獲得白樺組培苗。本研究選取生長1個月左右的組培苗進行白樺GUS組織化學染色,同時依次選取組培苗經盆栽生長2個月植株的頂芽、幼葉、成熟葉、初生莖、過渡莖、成熟莖及根進行不同組織部位的特異性表達分析。
1.2 主要試劑及菌株
TransStart?FastPfu Fly DNA Polymerase購自全式金;同源重組酶Mut Express? Ⅱ Fast Mutagenesis Kit V2購自諾唯贊公司;Plasmid mini Kit Ⅰ、Gel extraction kit購自杭州博日科技(Bioer)有限公司;DNA Marker DL5000、反轉錄試劑盒、SYBR? Premix Ex TaqTMⅡ、DNA連接酶、限制性內切酶HindⅢ等均購自TaKaRa公司(大連);大腸桿菌感受態(Trans1-T1)購自TransGen Biotech公司;農桿菌感受態(GV3101)購自上海唯地生物技術有限公司;pBI101-35S::GUS載體質粒由本實驗室提供,其他實驗試劑均為進口或國產分析純。
1.3 實驗方法
1.3.1 白樺BpJMJ18基因組織特異性表達分析
利用CTAB法[22]分別提取盆栽生長2個月白樺頂芽、幼葉、成熟葉、初生莖、過渡莖、成熟莖及根的RNA,利用TaKaRa公司的反轉錄試劑盒將其總RNA反轉錄為cDNA,反轉錄的cDNA稀釋10倍作為定量PCR的模板,選用18S基因作為內參,其在NCBI網站的登錄號為:MK388236.1,來檢測目的基因在不同組織部位的表達情況。定量PCR使用的相關引物如下表1所示,反應體系為:20 μL;其中2×SYBR Green 10 μL,ROX Dye Ⅱ 0.4 μL,正向引物(JMJ18-RT-F)和反向引物(JMJ18-RT-R)各0.8 μL,反應程序為:95℃ 30 s;95℃ 5 s;60℃ 35 s;40cycle;95℃ 15 s;60℃ 1 min;95℃ 15 s。每個樣品設置3次生物學重復,并通過2-ΔΔCt法進行數據分析。
表1 實時熒光定量PCR引物
Table 1 Primer sequence of the quantitative real-time PCR
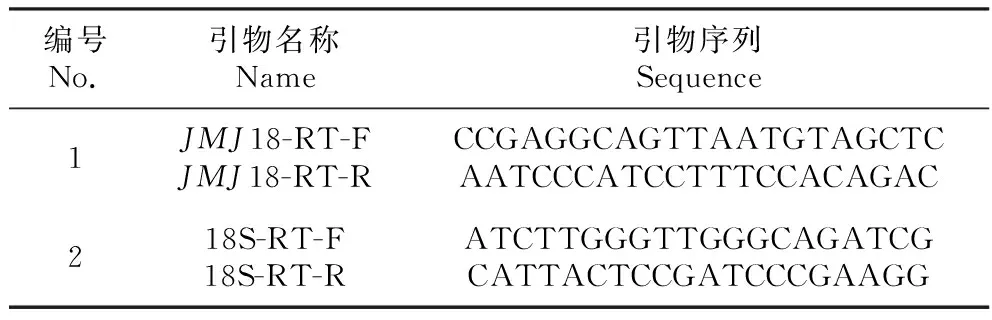
編號No.引物名稱Name引物序列Sequence1JMJ18-RT-FJMJ18-RT-RCCGAGGCAGTTAATGTAGCTCAATCCCATCCTTTCCACAGAC218S-RT-F18S-RT-RATCTTGGGTTGGGCAGATCGCATTACTCCGATCCCGAAGG
1.3.2 白樺BpJMJ18基因啟動子的克隆
根據已知的BpJMJ18基因在擬南芥中的序列,其在擬南芥Tair網站的命名為At1g30810,與白樺基因組數據庫進行對比,克隆了白樺BpJMJ18基因上游1 932 bp的啟動子區域,并設計帶有酶切位點的引物,引物由吉林省庫美生物科技有限公司合成。上、下游引物序列分別為BpJMJ18-promoter-F:5′-FATCAAGCTTAGTCAAAGCATCCTATGAAAACCCAGC-3′(下劃線處為HindⅢ酶切位點);BpJMJ18-promoter-R:5′-ATCAAGCTTAGTCAAAGCATCCTATGAAAACCCAGC-3′(下劃線處為HindⅢ酶切位點);以白樺組培苗葉片基因組DNA為模板擴增全長啟動子序列,PCR反應體系為50 μL,含10 μL 5×pfu Buffer、4 μL 2.5 mmol·L-1High pure dNTPs、1.0 μL上游引物(BpJMJ18-promoter-F)、1.0 μL下游引物(BpJMJ18-promoter-R)、2.0 μL模板、1.0 μL pfu酶、31 μL ddH2O。反應條件為95℃預變性2 min;95℃變性20 s,57℃退火20 s,72℃延伸1 min,40個循環;72℃延伸5 min。PCR反應完成后將產物經1.0%的瓊脂糖凝膠電泳檢測。
1.3.3 白樺BpJMJ18基因啟動子序列分析
利用PLACE數據庫(http://www.dna.affrc.go.jp/PLACE)和Plant CARE數據庫(http://bioinformatics.psb.ugent.Be/webtools/plantcare/html)在線分析軟件,對BpJMJ18基因進行啟動子順式作用元件分析。
1.3.4 pBI101-BpJMJ18pro::GUS載體構建及瞬時轉化
通過1.3.1中引物擴增目的片段,連接pEASY-Blunt載體并轉化大腸桿菌Trans1-T1,隨機挑取6個單克隆,進行PCR檢測并送至擎科生物公司測序保存。將測序結果正確的菌液擴大培養,利用限制性內切酶HindⅢ對pEASY-Blunt-BpJMJ18-promoter和pBI101-35S::GUS質粒進行單酶切,分別回收目的片段,利用TaKaRa公司的SolutionⅠ進行連接并轉化大腸桿菌。隨機挑取單克隆進行PCR檢測,將獲得的陽性轉化子菌液送公司測序,保存測序正確的重組質粒,命名為:pBI101-BpJMJ18pro::GUS。最后采用液氮凍融法將重組質粒轉入農桿菌GV3101中,挑取單克隆經菌液PCR驗證轉化子。
參考李萌[23]等瞬時轉化白樺的方法,并修改如下:將pBI101-BpJMJ18pro::GUS菌種接種到7 mL含卡那霉素的液體LB培養基中,28℃ 180 r·min-1震蕩過夜進行活化培養;第二天接種活化好的菌液于100 mL含卡那霉素的液體LB培養基中,28℃ 180 r·min-1震蕩培養至OD600=0.5,收集菌體,用1/2MS液體培養基將菌體進行重懸,使其OD600=0.8,取長勢良好的白樺組培苗放入制備好的菌液中,25℃ 120 r·min-1培養2 d,每間隔16 h更換1次1/2MS液體培養基。
1.3.5 GUS組織化學染色分析
將瞬時侵染后的白樺苗放置于GUS染液中(配方如下:100 mL 50 mmol·L-1磷酸鈉緩沖液(pH7.0),0.372 g 10 mmol·L-1EDTA·2 Na溶液,100 μL 0.1% Triton X-100,0.016 5 mg 0.5 mmol·L-1亞鐵氰化鉀,0.021 1 mg 0.5 mmol·L-1鐵氰化鉀,500 μL 1 mg·L-1X-Gluc),進行GUS染色,37℃條件下避光溫浴過夜培養,將植株置于脫色液中脫色6~8 h(脫色液∶無水乙醇∶冰乙酸=3∶1),期間及時更換脫色液。待脫色完全后,通過顯微鏡觀察BpJMJ18基因啟動子的表達情況并拍照保存。
2 結果與分析
2.1 白樺BpJMJ18基因組織特異性表達分析
為了研究白樺BpJMJ18基因在各個組織部位的表達情況,提取白樺頂芽、幼葉、成熟葉、初生莖、過渡莖、成熟莖及根的total RNA,進行熒光定量PCR檢測,結果如圖1所示。表明BpJMJ18基因在各個組織部位中均有所表達,且BpJMJ18基因的表達模式在根和幼葉中的表達水平高于其他組織部位,在成熟莖的表達量相對較低,上述結果說明BpJMJ18基因在植株的根和葉中表達更為廣泛。
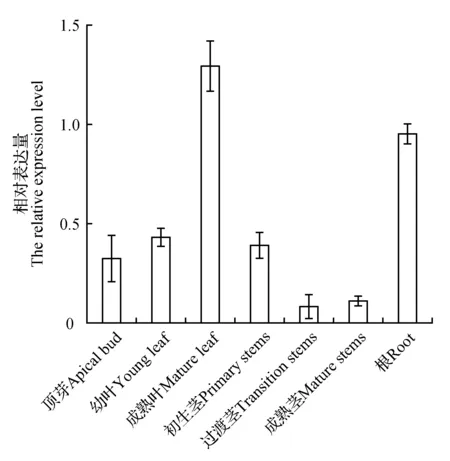
圖1 BpJMJ18基因在白樺不同組織表達分析Fig.1 Expression analysis of BpJMJ18 gene in different tissues of B.platyphylla
2.2 白樺BpJMJ18基因啟動子的克隆
通過上述組織特異性表達分析結果,利用PCR技術從白樺葉片中擴增出長度為1 932 bp的目的片段(見圖2A),膠回收目的片段后連接pEASY-Blunt載體,轉化大腸桿菌Trans1-T1感受態,隨機挑取6個單克隆進行菌液PCR檢測(見圖2B)。將菌液檢測成功的轉化子送公司進行測序,結果表明插入目的片段未發生堿基突變,成功克隆BpJMJ18基因啟動子。
2.3 白樺BpJMJ18基因啟動子的序列分析及元件預測
利用PLACE和Plant CARE數據庫對1 932 bp啟動子進行順式作用元件預測分析,結果如圖3所示,該序列中除含有TATA-box和CAAT-box等基本順式作用元件外,還具有光響應元件如G-Box、Box4、GT1-motif、TCT-motif;參與脫落酸反應的元件ABRE;MEJA反應的調節元件CGTCA-motif;厭氧誘導所必須的調節元件ARE;赤霉素和生長素響應元件P-box、TGA-element;除上述相關的響應元件外,BpJMJ18啟動子還具有分生組織表達相關作用元件CAT-box(見表2)。由此可以推測BpJMJ18啟動子可能在空間表達過程中受到光周期的調控、參與多種植物激素的響應,具有多種生物學功能,在植物生長發育過程中扮演不同角色。
2.4 pBI101-BpJMJ18pro::GUS載體構建
將測序正確的pEASY-Blunt-BpJMJ18pro菌液提取質粒,利用限制性內切酶HindⅢ對pEASY-Blunt-BpJMJ18pro和pBI101-GUS進行單酶切(見圖4A),回收目的片段,利用TaKaRa公司的SolutionⅠ進行連接,轉化大腸桿菌(Trans1-T1),隨機挑取單克隆進行菌液PCR檢測(見圖4B),檢測正確的轉化子送擎科公司進行測序,結果表明插入的BpJMJ18啟動子序列未發生堿基突變,至此植物表達載體構建完成,構建示意圖如圖4所示,并命名為pBI101-BpJMJ18pro::GUS。然后將pBI101-BpJMJ18pro::GUS質粒轉入農桿菌感受態GV3101中,隨機挑選單克隆進行PCR檢測(見圖4C)。
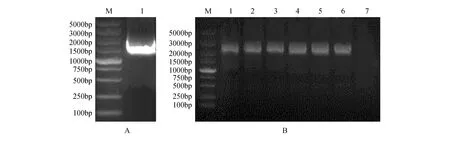
圖2 白樺BpJMJ18基因啟動子的克隆及鑒定 A. BpJMJ18啟動子的克隆(M. DNA marker DL5000;1.JMJ18啟動子的PCR擴增產物);B.pEASY-BpJMJ18pro載體在大腸桿菌菌液中的PCR檢測(M. DNA marker DL5000;1~6.pEASY-BpJMJ18pro轉化子的菌液PCR檢測;7.陰性對照)Fig.2 The clone and verification of BpJMJ18 gene promoter from B.platyphylla A. The cloning of BpJMJ18 promoter(M. DNA marker DL5000; 1. PCR amplification product of JMJ18 promoter); B. PCR detection of pEASY-BpJMJ18pro vector in E.coli(M. DNA marker DL5000; 1-6. PCR detection of pEASY-BpJMJ18pro transformants; 7.Negative control)

圖3 BpJMJ18啟動子作用元件示意圖Fig.3 BpJMJ18 promoter action element diagram
2.5 白樺BpJMJ18基因啟動子瞬時轉化及活性分析
為了進一步驗證BpJMJ18啟動子在白樺中的表達活性,我們將用農桿菌介導的瞬時轉化法侵染白樺組培苗,進行了3次重復試驗并通過GUS染色觀察其表達特征。結果如圖5A所示,發現BpJMJ18啟動子驅動的GUS基因在白樺的主根、側根、根尖、葉片的維管束和嫩莖中均檢測到表達,在老莖中表達水平較低,而未經過侵染的野生型白樺未檢測到GUS活性(見圖5:B~E),這與前人在擬南芥中進行GUS組織化學染色表現出相似的表達模式。說明BpJMJ18啟動子具有驅動GUS表達活性的能力,并在白樺植株的各個組織部位中發揮作用進而影響植株的生長發育。

表2 BpJMJ18啟動子中順式作用元件
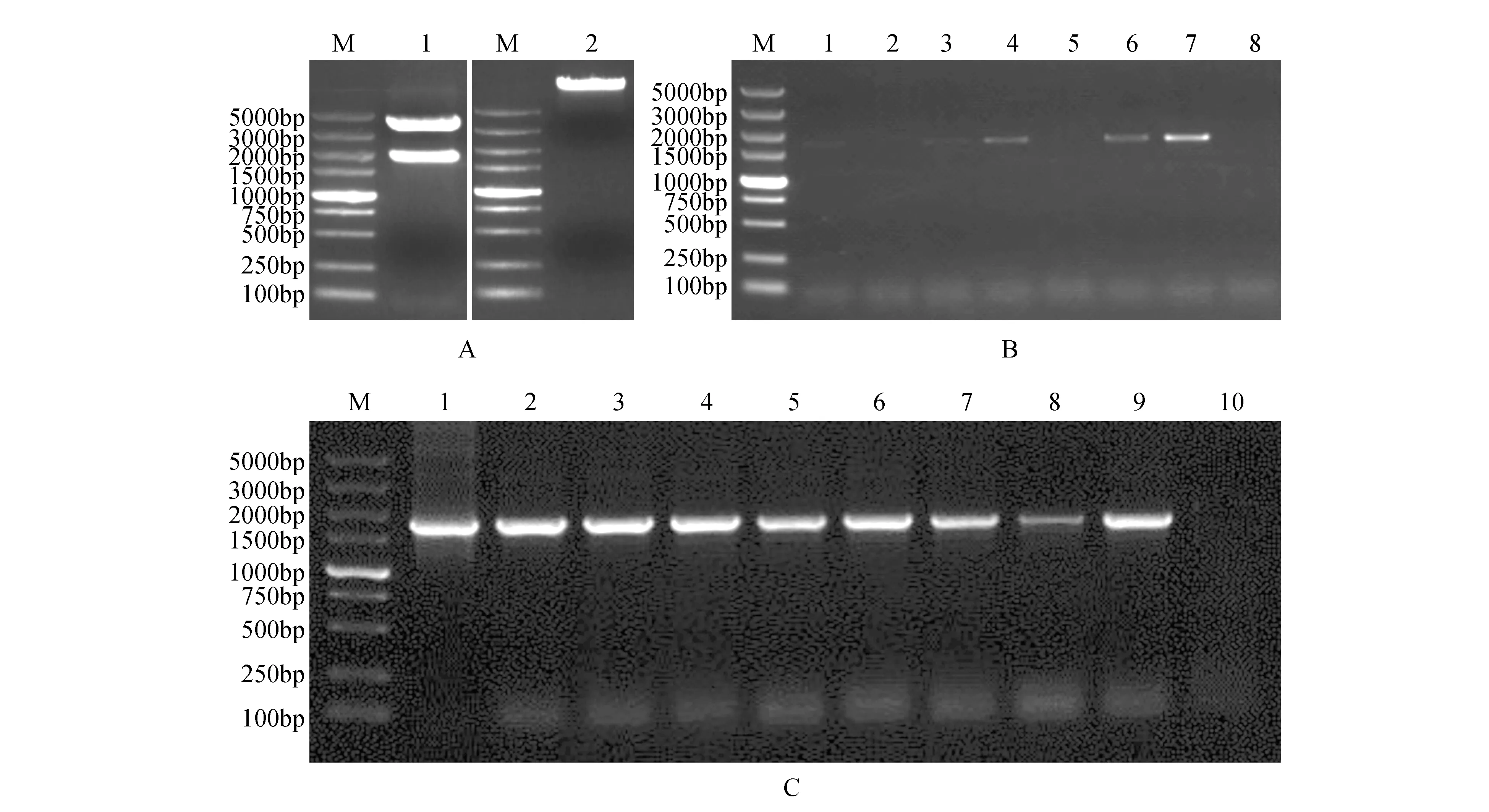
圖4 植物表達載體pBI101-BpJMJ18pro::GUS的構建及驗證 A.限制性內切酶酶切產物的結果(M. DNA marker DL5000;1.pEASY-Blunt-BpJMJ18pro單酶切條帶;2.質粒pBI101-GUS的單酶切條帶);B.植物表達載體pBI101-BpJMJ18pro::GUS的菌液PCR檢測(M. DNA marker DL5000;1~7.pBI101-BpJMJ18pro::GUS菌液的PCR產物;8.陰性對照);C. GV3101-pBI101-BpJMJ18pro::GUS的菌液PCR檢測(M. DNA marker DL5000;1~9.農桿菌轉化子的菌液PCR檢測;10.陰性對照)Fig.4 The construction and verification of plant expression vector pBI101-BpJMJ18pro::GUS A.Electrophoretogram of restriction enzymes digestion(M. DNA marker DL5000; 1.Digestion of pEASY-Blunt-BpJMJ18pro with HindⅢ; 2.Digestion of pBI101-GUS vector with HindⅢ); B.PCR detection of plant expression vector pBI101-BpJMJ18pro::GUS(M. DNA marker DL5000; 1-7.PCR products of pBI101-BpJMJ18pro::GUS vector; 8.Negative control); C.PCR detection of GV3101-pBI101-BpJMJ18pro::GUS(M. DNA marker DL5000; 1-9.PCR products of transformants in Agrobacterium; 10.Negative control)

圖5 BpJMJ18在白樺中的活性分析 A.白樺組培苗;B.頂芽;C.莖;D.第三葉片;E.根Fig.5 BpJMJ18 promoter action element diagram A.Birch tissue culture seed; B.Apical bud; C.Stemt; D.Stem; D.Leaf; E.Root
4 討論
JMJ類家族蛋白含有JmjC保守結構域,編碼JmjC結構域蛋白的基因在許多生物體中代表了一個大的基因家族,如:人類中有30個基因,小鼠中有30個基因,酵母中有12個基因[24]。在植物中,通過生物信息學分析發現,21個擬南芥基因和20個水稻基因編碼JmjC結構域蛋白[25~26]。通過上述大量基因研究表明JMJ家族蛋白在不同的生物學過程中可能扮演一個重要的作用。因此,本研究從白樺基因組中鑒定出了一個編碼JmjC結構域的基因BpJMJ18,并分析了該基因的組織特異性表達模式,結果表明它在各個組織部位中均有所表達,且在根和幼葉中的表達水平高于其他組織部位,上述結果說明BpJMJ18基因可能參與了植物的生長發育過程。由于啟動子中的順式作用元件對于基因的時空表達及表達量都具有重要的調控作用[27~28],我們克隆了JMJ18基因的啟動子序列,通過在線軟件分析發現,該序列具有多個TATA-box和CAAT-box核心啟動元件,說明其具有典型啟動子的特征。
啟動子除具有典型的核心啟動元件外,還有眾多與基因功能相關的調節元件。因此,在相近基因的啟動子序列中,應該存在很多一致的順式作用元件。張俊飛等[1]克隆分析玉米JMJ15基因啟動子發現,在啟動子序列內含有多個光響應、激素響應和逆境響應等相關元件。湯小鳳[29]克隆的楊樹PtrJMJ25基因啟動子中對光處理有很強的響應,存在光響應元件。本研究也得到了類似的結果,在BpJMJ18基因啟動子序列內發現多個光響應元件(G-Box、Box4)和激素(P-box、TGA-element)、逆境響應(ABRE、CGTCA-motif)相關的元件。其中,ABRE和CGTCA-motif分別是與植物生長相關、逆境響應相關的元件,這些結果預示著BpJMJ18基因對于植物的生長發育具有重要的調節作用。此外,研究BpJMJ18啟動子序列相關的順式作用元件為揭示BpJMJ18基因功能具有重要的參考價值。因此本文主要圍繞BpJMJ18基因啟動子是否具有啟動活性功能展開。
目前,能夠準確分析啟動子活性和組織表達特異性的方法是穩定遺傳轉化法,例如:Eun-HyeHong[26]等人通過穩定轉化法將JMJC家族中AtJMJ3、AtJMJ4、AtJMJ6、AtJMJ7、AtJMJ8、AtJMJ9、AtJMJ10、AtJMJ11等基因的啟動子轉入擬南芥中,發現該家族基因均在擬南芥的種子、花、葉片、莖的分生組織和根中有所表達。同時觀察GUS組織化學染色發現不同基因在相同組織表達部位也有所差異。Hongchun Yang[30]等人將AtJMJ18基因的啟動子穩定轉化到擬南芥中,GUS染色發現該基因在擬南芥的根韌皮部、莖、子葉、幼葉、成熟葉和花中均有表達,在根和葉片中表達更廣泛。但是此方法操作復雜、周期較長且必須獲得相應的轉基因株系才能進行下一步的相關研究。相對于此方法而言,本文所使用的瞬時轉化法能夠在短時間內就可以分析哪些啟動子序列是調節基因表達所必需的;它簡單、方便,并且能夠快速鑒定啟動子的活性及相應的表達部位。本研究采用瞬時轉化法將含有BpJMJ18啟動子序列的植物表達載體轉化到白樺中,并對瞬時侵染后的轉基因植株進行GUS染色分析。結果表明,BpJMJ18基因能夠驅動GUS報告基因在主根、側根、根尖、葉片的維管束和嫩莖中均檢測到特異表達并且在成熟莖中有較弱的表達,這和Hongchun Yang[31]等發現的擬南芥根的韌皮部、莖、子葉、幼葉、成熟葉和花中均有所表達的研究結果類似。綜上研究結果說明白樺BpJMJ18基因的啟動子序列具有典型的高等植物啟動子特征,能夠驅動GUS基因在瞬時轉化的白樺中特異定位表達,表明該基因的啟動子具有啟動活性,并且存在組織特異性。
綜上所述,本研究利用PCR技術克隆白樺BpJMJ18基因啟動子,通過順式作用元件預測分析及瞬時轉化啟動子驅動報告基因表達對BpJMJ18探其功能提供了新思路,為進一步的研究BpJMJ18基因對白樺生長發育方面的影響奠定理論基礎。對于BpJMJ18基因啟動子的研究發現它在植物生長發育過程中可能參與了不同的生物學過程,具體的功能還需要進一步的實驗驗證。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
