豬圓環病毒2型感染人源細胞的轉錄組分析
2020-07-17 06:18:46劉小花徐鵬張昕薇歐陽婷吉衛龍劉曉慧任林柱
中國比較醫學雜志 2020年6期
劉小花徐 鵬張昕薇歐陽婷吉衛龍劉曉慧任林柱
(吉林大學動物科學學院,長春 130062)
豬圓環病毒(porcine circovirus,PCV)是圓環病毒科圓環病毒屬中的一員,為緊密的球形無包膜單鏈環狀DNA病毒。根據基因組特點,可將PCV分為 PCV1型(porcine circovirus 1,PCV1)、2型(porcine circovirus 2,PCV2)和3型(porcine circovirus 3,PCV3)[1]。對PCV的研究初期,我們普遍以為這類病毒只能寄生在豬體或豬源細胞中。但隨著科研人員的進一步研究發現,在非洲綠猴腎細胞及其他細胞中也發現了PCV的寄生,并且能夠進行分離與傳代[2]。PCV2與PCV1不同,它能夠引起豬圓環病毒病(Porcine circovirus disease,PCVD)或豬圓環相關病毒病(Porcine circovirus associated disease,PCVAD)[2-3]。突出表現在豬皮炎腎病綜合征(Porcine dermatitis and nephropathy syndrome,PDNS)、斷奶仔豬多系統衰竭綜合征(Post-weaning muhisystemic wasting syndrome,PMWS)、豬呼吸道疾病綜合征(Porcine respiratory diseases complex,PRDC)和母豬繁殖障礙(Reproductive failure)等疾病中,給養豬業造成了巨大的經濟損失[2]。
PCV2感染會引起豬源細胞內一系列的信號改變,這些信號包括PCV2增殖、細胞凋亡和自噬的啟動以及抗病毒應答的誘導。已有研究發現許多與細胞信號通路相關的蛋白質可作為抗病毒效應的靶標[3]。研究發現,PCV1能夠感染人源細胞,且能夠引起細胞超微結構的改變[4-5],但PCV2是否也能感染人源細胞的問題一直存在爭議。Hattermann等人[6]在2004年用帶有 PCV2全長的質粒感染293T、HeLa等人源細胞后,發現在所有轉染細胞中都擴增出了PCV2的DNA,并且發現了病毒抗原的表達;隨后,轉染3 d后的細胞出現病變,細胞形態變為圓形,上清液中死亡細胞和細胞碎片數量增加,但在兩周之后,病毒顆粒消失,證明PCV2不能在人的細胞中穩定存在[6]。往后其他研究人員的調查顯示,沒有證據表明PCV2接觸者的血清中存在PCV2特異性抗體,表明PCV2在人類細胞是有限存在[7-9]。直至2019年,我們用PCV2感染12種人的細胞,包括單核細胞、上皮以及成纖維細胞,結果發現PCV2都能夠穩定感染這些細胞,且產生可感染性的病毒粒子[10]。但關于人源細胞感染PCV2之后所引起的免疫反應及抗病毒效應知之甚少。
RNA-seq技術是近些年發展較快的一種高通量測序技術,具有測序量大且快速、應用范圍廣等優勢[11]。本文通過PCV2感染HeLa細胞后的轉錄組測序分析,將篩選出來的差異表達基因(differentially expressed genes,DEGs)做進一步分析,預測其相關的信號通路以及生物學功能,最后篩選出與抗病毒反應相關的基因,并通過實時定量PCR方法進一步驗證,以期為人源細胞對PCV2及其他病毒產生抗病毒效應的研究提供參考。
1 材料和方法
1.1 實驗材料
PCV2株CC1(GenBank登錄號JQ955679)是從患有PMWS癥狀的豬組織標本中分離得到,并由本實驗室培養及儲存[12]。人源細胞系MCF-7、HeLa和A549購自上海通威生物科技公司[10]。
1.2 主要試劑與儀器
TRNzol-A+試劑(天根生化科技有限公司,中國北京);Agilent 2100生物分析儀(Agilent Technologies,Santa Clara,CA,USA);TruSeq鏈mRNA LT樣品制備試劑盒(Illumina,San Diego,CA,USA);磁性寡糖(dT)珠(Anchored oligo(dT)23primers,Sigma);Illumina HiSeqTM 2500平臺(Illumina,美國)。
1.3 實驗方法
1.3.1 RNA制備與RNA序列測定
簡單來說,接毒前1 d分板,待MCF-7、HeLa和A549細胞長至50%~60%時,用感染復數為1的PCV2株CC1毒液感染細胞,1 h后棄掉病毒液,用新鮮培養液輕輕洗滌1~2遍,然后加入正常培養液繼續培養,未感染組同期進行,72 h后用TRNzol-A+試劑提取 RNA。總 RNA溶于50μL去 RNase的ddH2O中,并在-20℃下保存。
為制備cDNA文庫,用去RNase的DNaseⅠ處理總RNA,然后用磁性寡糖(dT)珠純化mRNA,并用Agilent 2100生物分析儀檢測RNA完整性。RNA完整性指數(RINs)>8的可進行后續分析。純化的mRNA用TruSeq鏈mRNA LT樣品制備試劑盒構建文庫。然后,在Illumina HiSeqTM 2500平臺上對這些文庫進行測序。
1.3.2 差異基因的功能注釋與富集分析
利用Blast2 GO軟件將所有的UniGenes比對到各大平臺的數據庫中,最后在Nr數據庫可注釋到最大的比對率。再將測序獲得的所有DEGs進行基因功能注釋(Gene Ontology,GO)分析和分類,獲得轉錄本的GO功能注釋信息。DEGs比對GO數據庫后,獲得 GO條目(term)。通過超幾何檢驗(Phyper),將校正P值(correctedP-value)<0.05的GO term定義為DEGs中顯著富集的GO term。將DEGs應用京都基因與基因組百科全書(Kyoto The Encyclopedia of Genes and Genomes,KEGG)數據庫預測其參與的代謝途徑(correctedP-value≤0.05為篩選標準)。再利用分子相互作用分析軟件(STRING)預測DEGs相互作用網絡,用 Cytoscape軟件構建蛋白相互作用網絡。
1.3.3 定量PCR分析基因表達
獲得PCV2感染72 h以及未感染細胞的總RNA后,并用FastQuant RT-Kit(天根生化科技有限公司,中國北京)進行反轉錄。所得cDNA用Bio-Rad IQTM多色實時 PCR檢測系統和 Luna?Universal qPCR Master Mix(新英格蘭生物實驗室,美國)進行SYBR Green定量實時PCR。
擴增20μL體系,上下游引物各0.5μL,cDNA 2μL,2×Luna通用 qPCR混合物 10μL,ddH2O 7μL,采用以下循環條件:95℃預變性60 s,95℃變性15 s,并在60℃下退火而后延伸30 s,40個循環。實驗至少重復3次。
2 結果
2.1 PCV2感染人源細胞的轉錄組分析
本研究利用Illumina HiSeq平臺共測了2組樣品(每組樣品有三個重復),每組樣品平均有6.40 Gb的數據。共檢測到表達的基因數為15402個,其中已知基因14817個,預測新基因有585個。
對感染72 h及未感染PCV2的HeLa細胞進行的轉錄組RNA序列分析,結果如表1所示。在感染PCV2的HeLa細胞中共鑒定出387個DEGs,包括267個上調基因和120個下調基因(表1,圖1A)。在387個DEGs中,291個基因與生物過程有關。因此,選取生物過程(biological process)的差異基因做進一步GO分類分析。結果顯示共有212個基因上調,79個基因下調。此外,調控基因可分為15個功能組別(圖1B、1C),表明PCV2感染可上調或下調多種人類基因。在上調基因中,占比前四的組別分別是生物調控(biological regulation)、刺激應答(response to stimulus)、細胞過程(cellular process)以及代謝過程(metabolic process),分別占上調基因總數的15%、11%、10.8%和10.7%(圖1B)。在下調基因中,同樣是生物調控、刺激應答、細胞過程和代謝過程這幾個功能組別占比較高,分別為16.9%、8.7%、17.2%和10.6%(圖1C)。
2.2 PCV2感染細胞中DEGs的GO和KEGG途徑富集分析
為了了解DEGs在PCV2感染細胞中的主要途徑,我們進行了GO富集和KEGG通路分析,并預測了DEGs可能參與的生物學相互作用。GO富集結果表明,最常見的GO term是應激反應(97 DEGs)和免疫系統過程(80 DEGs)(圖2A)。
此外,將DEGs進行KEGG通路分析以進一步評估它們各自的功能。圖2B展示了PCV2感染的人類細胞中的前20個(按Q value從小到大排序)富集途徑。在HeLa細胞對PCV2感染的應答中,NOD樣受體信號途徑(NOD-like receptor signaling pathway)有21個DEGs,單純皰疹病毒感染(Herpes simplex infection)相關的途徑中有22個DEGs被高度富集(圖2B)。
2.3 參與PCV2感染的DEGs相關網絡分析
利用 Cytoscape軟件(http://www.cytoscape.org/,Version 3.6.1)的生物網絡基因功能注釋插件(Biological Networks Gene Ontology,BinGo)對基因的GO注釋進行了超幾何檢驗,P<0.001。如圖3A所示,DEGs在炎癥反應(inflammatory response)、細胞因子刺激反應(response to cytokine stimulus)、白細胞趨化性(leukocyte chemotaxis)、免疫應答(immune response)和病毒應答(response to virus)等生物過程中高度富集,表明這些生物過程與PCV2感染人類細胞的過程有關。
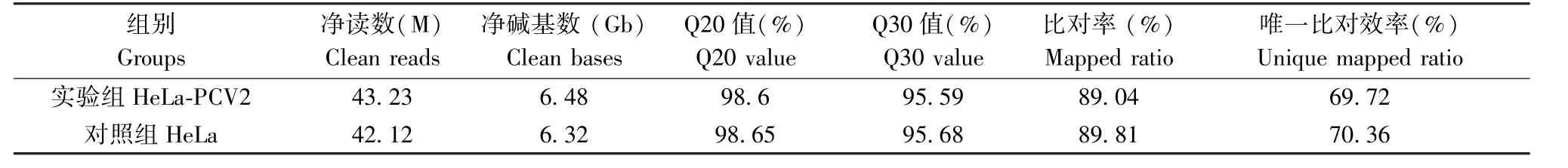
表1 RNA-Seq數據匯總Table 1 Summary of RNA-Seq data
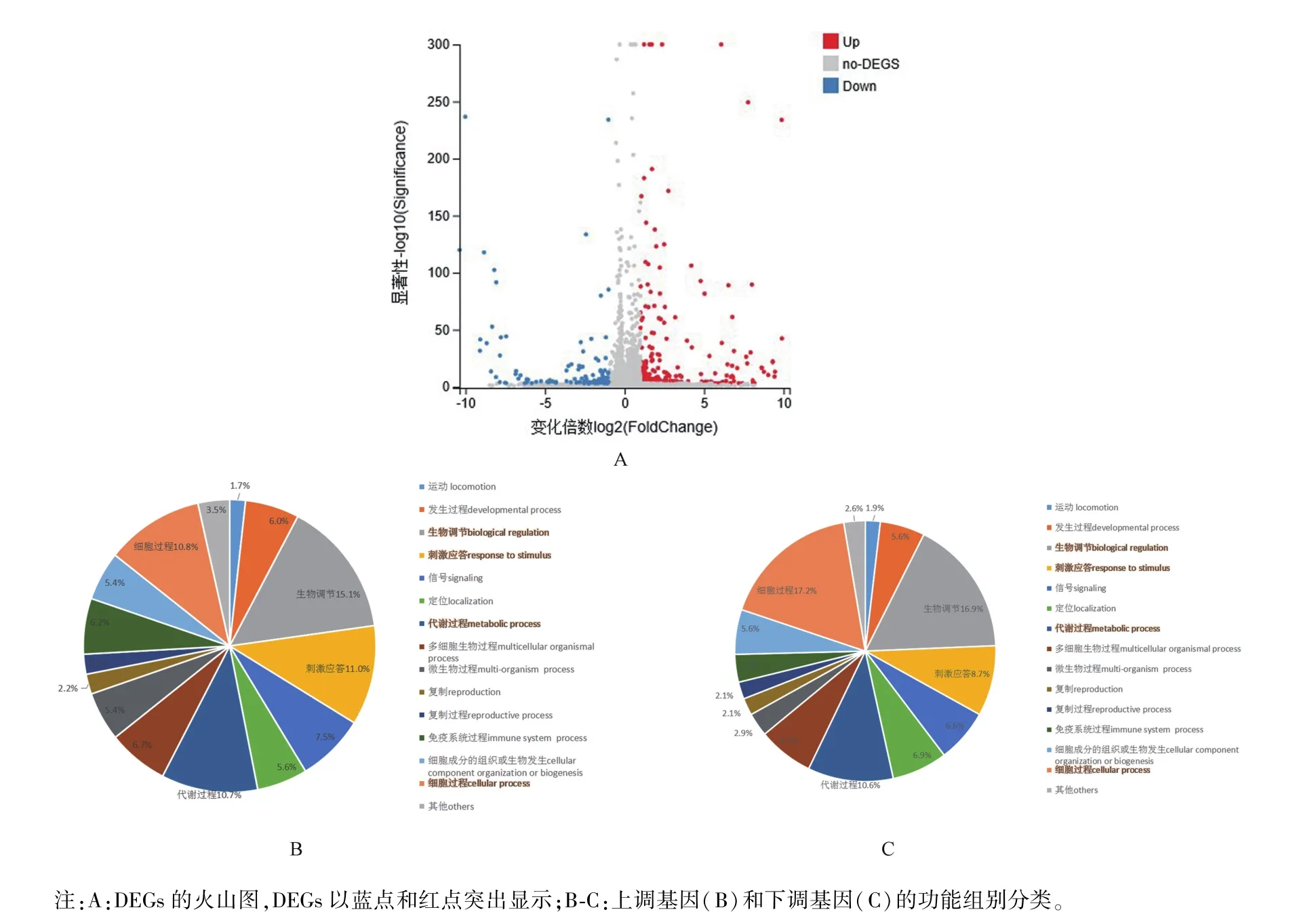
圖1 PCV2感染與未感染細胞基因差異表達分析Note.A,Volcano plot of DEGs.Significantly DEGs were highlighted in blue and red dot.B-C,Categories of the up-regulated genes(B)and the down-regulated genes(C).Figure 1 Differential expression analyses of human genes between mock and PCV2-infected cells

圖2 PCV2感染與未感染的HeLa細胞中GO富集的前20個生物過程Note.A,GO enrichment analysis.B,KEGG enrichment analysis.Circles indicate numbers of enriched genes and colors mean the Q value.Figure 2 Top 20 of GO terms between mock and PCV2-infected human cells
此外,利用STRING網站分析了與病毒應答相關的DEGs潛在的相互作用網絡。如圖3B所示,24個DEGs在PCV2感染后下調,包括 IRF9、IFI44L、DDX58、GBP1、GBP3、IFIT2、IFIT1、IFIT3、IL6、IRF1、IRF7、LGALS9、MX1、MX2、OAS1、OAS2、PLSCR1、DDX60、CCL5、 IFIH1(MDA5)、 DHX58(LGP2)、ZC3H12 A(MCPIP)、RSAD2和 ISG15(圖 3B)。
2.4 實時qPCR對DEGs的確認
為了進一步驗證上述RNA-Seq數據,選擇了部分與病毒應答相關的屬于刺激應答過程的DEGs,使用實時qPCR進行驗證。如圖4A所示,與未感染細胞相比,細胞感染PCV2之后,所選擇的所有DEGs顯著下調,但與RNA-Seq數據結果相反。
根據免疫系統過程(immune system process)、信號(signaling)和刺激應答(response to stimulus)三個生物學過程的GO富集,選擇了在PCV2感染HeLa后差異最顯著的前12個DEGs進行RT-qPCR驗證,并以MCF-7細胞做相同處理進行進一步驗證。如圖4B所示,在感染PCV2的HeLa細胞中,F8A2的表達沒有差異(FC<1.5);然而,CXCL3、CSF3、TNFAIP3和 ITGB2的表達水平明顯升高,HIST1H2AK、CXCL1、FBXO17、HBA1、SAA1、PTX3和PI3的表達水平明顯降低(圖4B)。此外,感染PCV2的 MCF-7細胞中 HIST1H2AK、FBXO17、PTX3和PI3的表達水平同HeLa細胞一致,也明顯降低,而其他基因 F8A2、CXCL1、HBA1、CXCL3、CSF3、SAA1、ITGB2和TNFAIP3的表達水平明顯高于未感染細胞(圖4C)。這些結果表明,不同人源細胞的抗病毒作用(特別是對病毒的應答)不盡相同。
為了進一步證實這些結果,用PCV2感染A549細胞72 h,隨機選擇前文中提到的DEGs做進一步實時qPCR驗證。結果顯示,病毒感染A549細胞后,SAA1、HIST1H2AK、PI3、F8A2、CXCL1、IL-6 和IFI44 L水平較未感染細胞明顯降低,與HeLa細胞相比,除F8A2結果不一致之外,其他基因變化趨勢均相同。
3 討論
PCV2對豬源細胞的易感性,對全世界養豬業造成了嚴重的威脅,但該病毒是否存在公共衛生風險是目前首要關心的問題。隨著以豬源器官為重點對象的異種移植技術的不斷興起與日趨成熟,PCV2能否感染人源細胞成為飽受爭議且需亟待解決的問題[2]。我們之前的研究中初步證明PCV2能夠感染人源細胞[10],這對于接觸此病毒的研究人員以及接受器官移植的患者來說都是一個非常大的安全隱患。除了對PCV2的防控措施做好之外,研究其抗病毒效應也顯得極為重要。
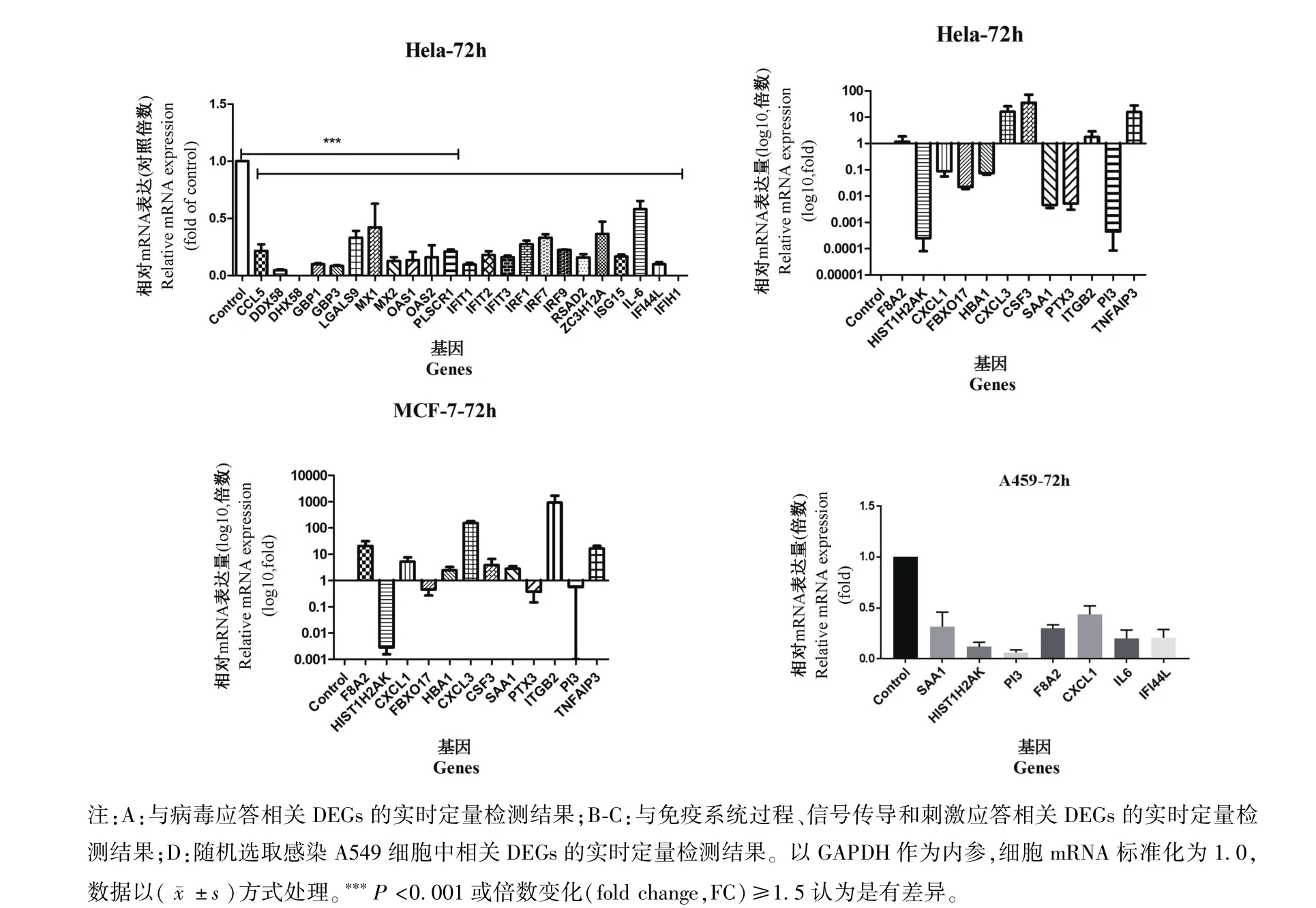
圖4 qRT-PCR結果Note.A,The results of Real-time PCR of related DEGs from response to virus.B-C,DEGs of the immune system process,signal and response to stimulus.D,Randomly selected some related DEGs from the infect A549 cells.Normalized with GAPDH,the celluar mRNA standardization was 1.0,***P<0.001 or FC≥1.5 was considered as the significance.Figure 4 The results of qRT-PCR
在本次研究中,選用HeLa細胞作為轉錄組分析的對象。在感染PCV2之后,使用Illumina HiSeq平臺一共測了 2個樣品,共注釋到 15402個UniGenes。感染PCV2之后有387個差異表達基因,267個上調基因和120個下調基因。Gene Ontology分為分子功能(molecular function)、細胞組分(cellular component)和生物過程(biological process)三大功能類。根據差異基因檢測結果進行功能分類。本研究主要選取生物過程做進一步分析。上調基因集中于生物調控(biological regulation),而下調基因集中于細胞過程(cellular process)。對291個生物過程有關的差異基因做進一步GO富集和KEGG通路分析發現,應激反應(97/291 DEGs)和免疫系統過程(80/291 DEGs)占比最高,表明在感染病毒之后可能通過調控細胞調控免疫因子來產生抗病毒效應的。KEGG通路分析發現最富集顯著的兩條途徑是NOD樣受體信號途徑(21/170 DEGs)和單純皰疹病毒感染(22/170 DEGs)。用BinGo插件對生物過程所有差異基因的GO注釋進行超幾何檢驗發現,其中對病毒應答(response to virus)的差異表達基因被高度富集,而這個過程的基因多數都與抗病毒研究相關。繼而對其中24個已知的DEGs進行實時定量PCR檢測,結果顯示這些病毒應答相關基因在感染之后全部下調。這與RNA-seq結果分析是相反的,實驗重復了三次,結果較為可信,差異原因可能是由于RNA-seq是大規模篩選的,反映的是樣本整體的基因表達變化趨勢,所以存在一些基因與qPCR的結果不一致,需要挑選大量的基因驗證。所以又在不同的細胞中選取了另外一些與免疫系統過程(immune system process)、信號(signaling)和刺激應答(response to stimulus)三個生物學過程相關的基因做進一步驗證。結果表明,PCV2感染可調節人源細胞基因的表達,雖然不同細胞各基因變化有差異,但大致趨勢相同,與RNA-seq結果有一些出入,這在其他一些研究中也出現過[13-14],其原因可能是由于Illumina HiSeq平臺測序雖然比較精準,但存在識別序列通常較短,有一定的錯誤率從而會出現較高假陽性的問題[15]。
在預測的病毒應答相關基因中,變化差別較大的基因有:人粘液病毒抗性2(human myxovirus resistance 2,MX2/MXB)、鳥苷酸結合蛋白 1/3(guanylate binding protein 1/3,GBP1/3)、四磷酸肽重復序列1(tetratricopeptide repeats 1,IFIT1)、維甲酸誘導基因-I(retinoic acid-inducible gene-I,RIG-I/DDX58)以及干擾素誘導的44樣蛋白(interferoninduced protein 44-like,IFI44L)。大量文獻報道,上述基因均是與細胞抗病毒反應、細胞免疫反應或細胞增殖密切相關的。例如,Mx2在感染細胞中與HIV-1相互作用,并在逆轉錄和組裝成完整病毒基因組過程的早期階段阻斷感染[16-18]。GBP1是多形性膠質母細胞瘤(glioblastoma multiforme,GBM)中表皮生長因子受體(epidermal growth factor receptor,EGFR)的新靶基因,在體內可促進GBM腫瘤的生長和侵襲,但在體外對GBM細胞增殖無明顯影響[19-20]。另外,GBP3通過抑制病毒轉錄和復制參與抗流感活性[21]。在表達IFIT1的細胞中觀察到較高水平的Ⅰ型干擾素(type I interferon,I-IFN),IFIT1能感應到RNA的甲基化狀態,并通過與缺乏2′-O甲基化的5′帽結構結合而抑制病毒[22],也能通過抑制翻譯起始的步驟發揮其抗病毒功能[23-24]。RIG-I和黑色素瘤分化相關基因5(Melanomadifferentiation-associated gene 5,MDA5)是作為RNA病毒限制因子的胞漿RNA解旋酶,有研究證明了RIG-I樣受體(The RIG-I-like receptors,RLRs)限制了卡波西肉瘤相關皰疹病毒(Kaposi’s sarcomaassociated herpesvirus,KSHV)的裂解再活化[25];Dvorak等人[26]用PCV2感染豬源間充質樣細胞系后發現,細胞主要通過RIG-I/MDA5通路產生大量的I-IFN來達到抗病毒應答的效果。IFI44L是I型干擾素刺激基因(type I interferon-stimulated gene,ISG),屬于IFI44家族[27],經IL-28A和IFN-a處理抑制細胞丙型肝炎病毒(Hepatitis C virus,HCV)復制后,IFI44L表達顯著增加[28]。但是,由于PCV2在感染豬源細胞的過程中具有免疫抑制作用[29-30],而病毒應答過程中的一些基因大多參與了免疫應答過程(如 ISG15、IL6、IRF1等),因此,推測PCV2感染人源細胞也可能具有免疫抑制作用或抑制相關抗病毒因子表達的作用,最終導致上述基因的mRNA水平在感染72 h后降低,但這些推測還有待進一步的研究證實。
總之,用PCV2感染宮頸癌細胞HeLa之后引起細胞內大量基因表達情況改變,利用RAN-seq技術系統性分析了胞內mRNA的差異變化,多數DEGs參與了免疫系統過程、應激反應以及免疫應答過程。本文著重檢測了與病毒應答以及免疫系統過程等相關的DEGs的mRNA水平變化,這些DEGs可能與細胞抗病毒反應或致病機制有關。目前,正在以這些基因編碼的蛋白質為研究對象,分析其在感染PCV2之后的活性及功能變化以及這些蛋白對PCV2和其他病毒感染的影響。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
