葉綠素熒光成像技術下的柑橘黃龍病快速診斷
2020-07-25 03:49:10翁海勇何城城許金釵青家興葉大鵬
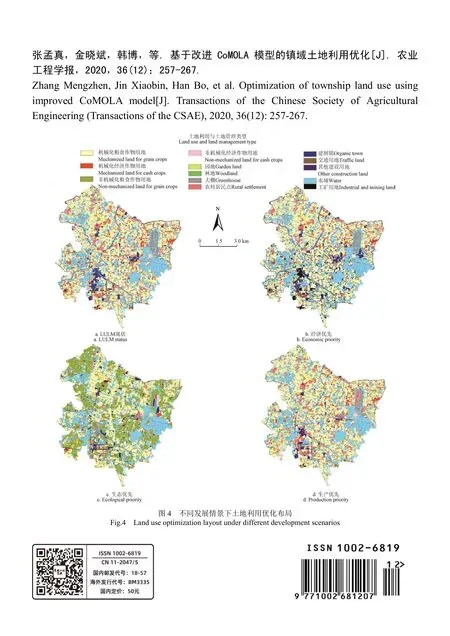
農業工程學報 2020年12期
翁海勇,何城城,許金釵,劉 浪,青家興,萬 亮,葉大鵬※
(1. 福建農林大學機電工程學院,福州 350002;2. 現代農業裝備福建省高校工程研究中心,福州 310002;3. 浙江大學生物系統工程與食品科學學院,杭州 310058)
0 引 言
柑橘黃龍病(Huanglongbing, HLB)是由一種僅限于韌皮部內寄生的候選韌皮部桿菌(CandidatusLiberibacter spp.)引起的細菌性病害,主要通過亞洲木虱或非洲木虱在田間傳播,具有極強的傳染力[1]。柑橘植株感染黃龍病后,一般經歷從幾個月到幾年不等的潛伏期之后才會顯現典型的黃龍病癥狀,如斑駁黃化和紅鼻子果[2]。然而在潛伏期時,病原菌已經在寄主樹體內增殖,而染病樹會發展成新的傳染源,進而引發黃龍病在果園中的擴散。通常柑橘黃龍病菌侵染會擾亂柑橘葉片正常的糖代謝功能,導致葉片的淀粉和可溶性糖含量異常累積[3-4]。Albrecht等[5]研究發現,柑橘黃龍病病原菌會影響宿主碳水化合物代謝相關基因表達,其中涉及淀粉合成的酶(AGPase)的基因表達量被上調,然而與淀粉分解酶(β-淀粉酶)相關基因的表達則被抑制。柑橘黃龍病目前尚不能被有效根治,常用控制的策略主要包括挖除感染黃龍病樹體、減少果園木虱數量和及時噴灑農藥等措施[6]。因此,及時、快速和準確診斷感染黃龍病柑橘樹體,對果園疫情防控具有重要的意義。
目前,柑橘黃龍病診斷的方法主要包括人工經驗法和聚合酶鏈式反應(Polymerase Chain Reaction, PCR)。由于人工經經驗判斷的方法主觀性強,檢測的準確率僅為59%左右[7],而PCR技術雖然準確率高,但也存在檢測效率低,成本高,以及對操作人員的專業水平要求高等缺點。光學成像技術作為一種先進的植物病害檢測方法,具有快速、操作簡單、無損等優點,在植物病害高通量檢測中具有重要的應用前景。Hawkins等[8]采用傅里葉紅外衰減全反射光譜儀采集了干燥柑橘葉片粉末信號,對染病葉片的識別正確率為95%。Weng等[9]采集健康和黃龍病葉片的可見/近紅外高光譜反射圖像,識別的正確率為92.6%。劉燕德等[10]利用高光譜成像技術結合機器學習算法,實現了柑橘葉片黃龍病準確率為94.4%的診斷。梅慧蘭等[11]分析了 4種不同病狀柑橘葉片高光譜反射圖像并比較了不同數據預處理方法,結果表明,經平滑濾波(Savitzky Golay, SG)或一階微分預處理后模型得到了96.4%的判別準確率。Deng等[12]利用無人機遙感高光譜成像技術分析了黃龍病病原菌對葉片光反射能力的影響,并采用支持向量機實現了柑橘黃龍病的快速診斷。Sankaran等[13]利用手持式熒光儀采集了健康和染病顯癥柑橘葉片的穩態熒光信號,利用決策樹算法獲得了97%的識別正確率,但對未顯黃龍病癥柑橘葉片的識別正確率僅為48.2%。Cen等[14]采用葉綠素熒光成像技術檢測柑橘黃龍病,總體正確率為 97%。盡管上述研究獲得了較好地識別效果,但僅針對的是顯現黃龍病癥柑橘葉片的識別。由于柑橘黃龍病致病性復雜,因此有必要從光吸收水平上探究病原菌對宿主光合作用過程中光能吸收、分配和利用的影響。
因此,本研究以感染黃龍病不同程度的柑橘葉片為研究對象,比較健康和染病葉片葉綠素熒光參數以及碳水化合物含量的差異,分析病原菌對宿主光能吸收和分配的影響及碳水化合物含量和葉綠素熒光參數關系,建立基于葉綠素熒光成像技術的柑橘黃龍病診斷模型,從而為實現柑橘黃龍病的快速診斷提供理論知識和技術支持。
1 材料與方法
1.1 試驗樣本
為探究感染黃龍病不同程度的柑橘葉片對病原菌在光合作用水平上的響應差異,以及建立柑橘黃龍病判別模型,本試驗于2016年8月從浙江臨海前岙村柑橘園中選擇經實時熒光定量多聚核苷酸鏈式反應(Real-time Quantitative Polymerase Chain Reaction, qPCR)鑒定過的健康和染病植株各3株,并于同年的11月再次采集已經標記過樹體的葉片。采樣時,從每棵植株的東南西北 4個方位分別剪下 3條枝條,并立即用蘸水的棉花包住枝條的切口,裝入自封袋后放入保鮮盒,減緩枝條上的水分散失。在采集葉綠素熒光圖像之前,再從枝條下取下葉片。本次試驗共收集了480片柑橘葉片(240片健康和240片染病)作為研究對象。其中,染病的柑橘葉片在7月和8月份均沒有癥狀,9月到11月份葉片的斑駁黃化癥狀逐漸明顯。
1.2 葉綠素熒光圖像采集
本研究利用葉綠素熒光成像技術分析宿主在光合作用水平上對病原菌的響應。本次試驗所采用的葉綠素熒光成像系統(FluorCam, Photon Systems Instruments, Brno,Czech Republic)主要包括分辨率為696×520 像素的CCD相機、焦距6 mm的鏡頭(VS Technology, Tokyo, Japan)、測量光和光化光1(620 nm),在距離光源20 cm位置,可均勻提供 0~306μmol/(m2·s)光強、飽和光(>8 000 K),可提供 0~4 053μmol/(m2·s)光強、高度可調節的樣品臺、暗箱。葉綠素熒光圖像采集前,首先對柑橘葉片進行25 min暗適應,以便使光合器官回到初始狀態,確保能準確地測量到葉片的原初光化學反應。光化光的強度設置與果園中平均光照強度相當,約為100μmol/(m2·s),飽和光強度則為1 500μmol/(m2·s)。本試驗所用的柑橘葉綠素熒光測量程序由 PSI公司提供(https://fluorcams.psi.cz/)。圖 1顯示了本試驗所用的柑橘葉綠素熒光測量程序,除了暗適應(t=5.56 s)和穩態時刻(Lss,t=92.24 s)分別施加1個飽和光外,在光適應過程中的L1時刻(t=32.24 s)、L2時刻(t=42.24 s)、L3時刻(t=52.24 s)和L4時刻(t= 72.24 s)和暗弛豫過程中D1時刻(t=122.24 s)、D2時刻(t=152.24 s)和D3時刻(t=182.24 s)也分別施加了1個飽和光,飽和光的施加時間點如圖1所示。

圖1 葉綠素熒光測量程序Fig.1 Measurement procedure of chlorophyll fluorescence
1.3 葉片碳水化合物含量和實時熒光定量聚合酶鏈式反應(PCR)分析
本試驗選擇美國戴安公司的離子色譜儀進行葉片可溶性糖含量的測量。色譜柱為Dionex CarboPACTM PA1分析柱(4 mm×250 mm)和(4 mm×50 mm)保護柱,柱溫保持30 ℃;淋洗液選擇流速為1.0 mL/min的NaOH,濃度為200 mmol/L;注入體積為10μL;檢測模式為電化學檢測。參考電極選擇Ag/AgCl;最后通過標準曲線來計算葉片中葡萄糖、果糖和蔗糖的含量。淀粉的含量則采用碘-淀粉顯色法。利用打孔器采集葉片對角4片面積為36.29 mm2的葉片小圓盤,去除葉綠素后,磨碎成懸浮液,煮沸,稀釋20倍后,取200μL的均勻懸浮液與15μL的2%碘液反應,20 min后,測量595 nm的吸光度值,最后利用淀粉溶液標準曲線計算出葉片的淀粉含量。同時,采用qPCR技術對每個月份的葉片健康狀態進行鑒定,引物為 OI1/OI2c(GCGCGTATGCAATACGAGCGGCA/GCCTCGCGACTTCGCAACCCAT)[15]。通過qPCR技術可以對沒有顯現黃龍病癥狀的柑橘葉片進行陰性或者陽性判斷,從而為進一步分析黃龍病對柑橘宿主光合作用的影響提供參考。
1.4 數據分析方法
為了使不同葉綠素熒光參數具有相同尺度,采用式(1)對原始的葉綠素熒光參數進行變換。

式中F′為變換后的葉綠素熒光參數,F為原始的葉綠素熒光參數,Fm和Fo分別為充分暗適應后的最大熒光和最小熒光。
通過主成分分析(Principal Component Analysis,PCA)將變換后的原始葉綠素熒光數據投影到一個新的坐標,獲取的前幾個主成分能夠反映大部分的原始信息,且相互獨立。各個成分的載荷系數(component loading coefficient)可用于解釋各個特征與對應因變量的關系,載荷系數絕對值越大,說明該特征對應變量越重要[16]。因此,成分的載荷可用于分析不同的葉綠素熒光參數對黃龍病病原菌侵染的響應。
葉綠素熒光信號與植物的光合作用能力密切相關,可以反映葉片的光化光淬滅和非光化光淬滅成分。因此,從葉綠素熒光圖像中獲取與健康葉片有區別的特征可作為判斷柑橘是否感染黃龍病的“指紋”特征。判別模型通過決策這些特征向量的歸屬,從而實現柑橘黃龍病的快速診斷。本研究選擇隨機森林(random forest)作為葉片中淀粉、蔗糖、葡萄糖和果糖的反演模型以及柑橘黃龍病的判別模型。隨機森林是由多個決策樹的分類器構成,通過將特征變量輸入每棵樹中進行分類,最后將若干個弱分類器的分類結果進行投票選擇[17]。為了使得隨機森林模型具有更好的非線性擬合或者分割能力,模型中決策樹的棵數節點處分類屬性的個數 2個參數需要優化。本研究設置樹的數量從 5~200,步長為 5,節點處分類屬性的范圍 2~98,步長為 2。采用 Kennard-Stone(KS)算法將數據集劃分成建模集和預測集。將不同的決策樹的棵數節點處分類屬性的個數進行組合,用預測集的決定系數來評估模型的性能。本研究中所使用的數據預處理方法和隨機森林模型在Matlab R2010a中完成。
2 結果與討論
2.1 柑橘黃龍病對柑橘葉片淀粉、蔗糖、葡萄糖和果糖含量的影響
圖 2顯示了健康和黃龍病柑橘葉片中淀粉、蔗糖、葡萄糖和果糖的含量。分析圖2a可知,感染黃龍病葉片中的淀粉含量(44.0±29.1)μg/mm2明顯高于健康葉片(20.3±5.7)μg/mm2,達到顯著性差異(P<0.05)。染病葉片中的淀粉的異常累積與淀粉合成能力被上調,而分解能力被下調有關。文獻表明,當柑橘感染黃龍病后,涉及淀粉合成酶(AGPase)的基因表達量被上調,淀粉分解酶(β-淀粉酶)的基因表達則被抑制[4]。葉片中淀粉的異常累積造成葉綠體類囊體膜裂解,影響葉綠體對光能的吸收、傳遞和轉化,降低葉片的光合作用效率[18]。
蔗糖作為光合作用另一主要產物,通過韌皮部從源器官轉運到庫器官。分析圖 2b,染病柑橘葉片的蔗糖變化趨勢與淀粉的變化趨勢一致,染病葉片(3.98±2.30 )μg/mm2中的蔗糖含量是健康葉片(2.48±1.12)μg/mm2的 1.61倍,二者差異達到顯著性水平(P<0.05)。柑橘黃龍病病原菌的侵染引起的宿主葉片蔗糖的異常累積可能是由于韌皮部遭受損傷后,蔗糖無法順利地從源器官轉運到庫器官。類似的現象也可以在僅限于韌皮部的病原菌(Phytoplasmas)侵染引起的椰子樹黃化病中觀察到[19]。進一步分析黃龍病病原菌侵染對宿主己糖(葡萄糖和果糖)代謝的影響可知(圖2c和圖2d),染病葉片中的葡萄糖(1.50±1.01)μg/mm2和果糖(0.72±0.65)μg/mm2含量明顯高于健康葉片的葡萄糖(0.91±0.64)μg/mm2和果糖(0.38±0.30)μg/mm2,分別是健康的1.65和1.86倍。這說明蔗糖水解成己糖代謝過程已經也受到了病原菌(CandidatusLiberibacter spp.)的影響。有文獻表明,微生物脅迫會使植物轉化酶的活性增加,會促進更多的蔗糖水解成葡萄糖和果糖[20-21]。與淀粉和蔗糖相比,葡萄糖和果糖的含量在不同季節的葉片中波動性更大,但是葡萄糖和果糖之間的含量差在染病和健康柑橘葉片中卻存在顯著性差異(P<0.05),說明了黃龍病病原菌對果糖和葡萄糖的選擇性不同,這與韌皮部寄生的病原菌對果糖具有偏好性相一致[22]。

圖2 健康和染病柑橘葉片的淀粉、蔗糖、葡萄糖和果糖含量Fig.2 Starch, sucrose, glucose and fructose in healthy and citrus Huanglongbing infected leaves
2.2 葉綠素熒光參數對病原菌侵染的響應分析
采用主成分分析(Principal Component Analysis,PCA)計算得到的載荷系數矩陣可反映出不同的葉綠素熒光參數對主成分的貢獻率,葉綠素熒光參數對應的載荷系數絕對值越大,貢獻越大,說明該參數對黃龍病病原菌侵染的響應更為敏感。前5個主成分(PC1、PC2、PC3、PC4和 PC5)的貢獻率分別為 41.80%、23.06%、11.92%、8.99%和4.5%,能夠表達原始變量的90.27%的信息。由于第1主成分的貢獻率最大,因此選擇分析第1主成分的載荷系數。由圖 3可知,充分暗適應后的最小熒光(Fo)、可變熒光與最小熒光的比值(Fv/Fo)、光適應和暗弛豫過程中的最小熒光(Fo_Ln/Dn),光系統II(Photosystem II, PSII)最大光量子效率QY_max(Fv/Fm)以及光適應和暗弛豫過程中的 PSII最大光量子效率(Fv/Fm_Ln/Dn)以及不可調制非光化光淬滅量子產額(ΦNO_Ln/Dn)對病原菌侵染的響應最為敏感。

圖3 第1主成分的載荷系數Fig.3 Loading coefficient of first principle component
選擇上述敏感葉綠素熒光參數進一步分析黃龍病病原菌對宿主光系統 II結構功能及原初化學反應的影響。圖 4顯示了健康和染病的柑橘葉片在暗適應、光適應、穩態和暗弛豫過程中不同時刻的葉綠素熒光參數。最小熒光(Fo)反映了葉綠體的光系統II反應中心的結構功能[23]。分析圖4a中健康和染病葉片的最小熒光(Fo),黃龍病病原菌的侵染致使宿主葉片的Fo上升,說明了病原菌會損傷光系統 II反應中心結構,能夠阻礙激發能向光反應中心轉移。同樣也可以發現染病葉片在光適應和暗弛豫的最小熒光(Fo_Ln和Fo_Dn)高于健康葉片,在L1、L2、L3、L4、Lss、D1、D2和D3時刻分別是健康葉片的1.28、1.27、1.26、1.26、1.26、1.27、1.28和1.28倍。病原菌致使宿主Fo上升同樣可以在感染白粉病的黃瓜葉片中發現[24]。Fv/Fo反映了類囊體膜上光系統II中有活性的反應中心的數量和尺寸,染病葉片的Fv/Fo(3.69±0.93)顯著低于健康葉片(4.51±0.88),說明黃龍病病原菌會顯著減少宿主光系統II中有活性的光反應中心數量(數據未列出)。QY_max(Fv/Fm)值反映了PSII的最大光量子效率的大小,由于該參數對脅迫十分靈敏,所以被視為植物遭受脅迫的指示器,也常被用于分析脅迫引起的植物光合效率變化[25-27]。分析圖4b可知,健康和染病葉片的QY_max存在差異,染病葉片的QY_max值(0.77±0.05)低于健康葉片(0.80±0.04),說明病原菌會降低PSII的最大光量子效率。同樣也可以發現染病葉片在光適應和暗弛豫的最大光量子效率(Fv/Fm_Ln和Fv/Fm_Dn)低于健康葉片,說明病原菌的侵染降低了染病葉片的光化學淬滅能力。分析不可調制非光化光淬滅量子產額ΦNO_Ln/Dn可知(圖 4c)。染病葉片的ΦNO_Ln/Dn顯著大于健康葉片,表明染病葉片的PSII出現了不可逆的損傷,受到光抑制。由于葉片將吸收的光能轉化成用于實際光合作用光量子產額(ΦPSII)、可調制非光化光淬滅量子產額(ΦNPQ)以及不可調制非光化光淬滅量子產額(ΦNO)。

圖4 不同時刻健康和染病葉片的最小熒光、最大光量子效率和不可調式非光化光淬滅量子產額Fig.4 Minimum fluorescence, maximum quantum efficiency and quantum yield of non-regulated non-photochemical energy dissipation of healthy and infected leaves at different phases
ΦNPQ_Ln/Dn對病原菌侵染的響應敏感性低于ΦNO_Ln/Dn和ΦPSII_Ln/Dn。從三者的此消彼長的競爭關系可知,染病葉片光化學反應效率的降低主要由于宿主受到光抑制后,將吸收的光能轉化成了不可調式非光化光淬滅量子產額(ΦNO),說明黃龍病病原菌能夠改變宿主葉片對激發能的分配。染病葉片光合作用效率的降低可能與光合作用相關基因如 photosystem II 5-kDa蛋白、photosystem-I subunit O和chlorophyll A-B binding家族蛋白有關的基因表達被下調有關[28]。
2.3 葉綠素熒光參數與葉片的淀粉、蔗糖、葡萄糖和果糖含量的相關性分析
柑橘葉片的碳水化合物含量、葉綠素熒光特性與黃龍病病原菌的侵染明顯相關,而葉片的碳水化合物含量又與葉片的光合作用有關,因此有必要進一步分析淀粉、蔗糖、葡萄糖和果糖與葉綠素熒光參數之間的相關性。圖 5顯示了淀粉、蔗糖、葡萄糖和果糖與葉綠素熒光參數之間的Pearson相關系數。淀粉和蔗糖與葉綠素熒光參數的相關性變化趨勢較為一致,而葡萄糖和果糖與葉綠素熒光參數的相關性變化趨勢則較為一致,造成這一現象源于淀粉和蔗糖均屬光合作用同化物,而葡萄糖和果糖則均是蔗糖的水解產物。相較于蔗糖、葡萄糖和果糖,淀粉與葉綠素熒光參數的相關系數絕對值>0.6以上的數量最多,且最高正相關系數可達 0.78,最高負相關系數可達?0.74。蔗糖、葡萄糖和果糖與葉綠素熒光參數的最高正相關系數分別可達 0.59、0.64和 0.52,最高負相關系數可達?0.56、?0.64和?0.56。相關性分析結果表明,葉綠素熒光參數與淀粉、蔗糖、葡萄糖和果糖具有明顯相關性,具有定量反演柑橘葉片中淀粉、蔗糖、葡萄糖和果糖含量的能力。

圖5 淀粉、蔗糖、葡萄糖和果糖與葉綠素熒光之間的Pearson相關系數Fig.5 Pearson correlation coefficient between chlorophyll fluorescence parameters and starch, sucrose, glucose and fructose concentration
2.4 柑橘葉片中淀粉、蔗糖、葡萄糖和果糖含量的反演
分析表 1可知,淀粉、蔗糖、葡萄糖和果糖樣本中的建模集均包含了最大值和最小值,且建模集和預測集對應的含量值分布范圍較廣,建模集的含量范圍覆蓋預測集的含量范圍,不同數據集的碳水化合物含量平均值和標準偏差較為相近。因此按照這種方法來劃分建模集和預測集是合理的。

表1 各數據集中淀粉、蔗糖、葡萄糖和果糖含量的描述性統計Table 1 Statistical characteristics of starch, sucrose, glucose and fructose
為了使隨機森林算法具有良好的分類或者回歸能力,模型中決策樹的棵數節點處分類屬性的個數兩個參數需要優化。圖 6顯示了不同組合的森林中決策樹的棵數和節點處分類屬性的個數建立的隨機森林模型對淀粉、蔗糖、葡萄糖和果糖預測精度的影響。

圖6 隨機森林模型中決策樹的棵數和節點處分類屬性的個數對其預測能力的影響Fig.6 Effect of the number of trees and number for splitting at each node on the predicting performance of random forest model
從圖 6可以看出,森林中決策樹的棵數對模型預測能力的影響大于節點處分類屬性的個數。模型中決策樹的棵數從5增加至25時,淀粉、蔗糖、葡萄糖和果糖隨機森林模型的預測能力均得到較大幅度的提升,當棵數>25時,模型的預測能力逐漸趨于穩定。通過優化,淀粉隨機森林預測模型的決策樹的棵數和節點處分類屬性的個數分別為56和5時,預測集的決定系數最高,為0.91。但是進一步分析可知,當節點處分類屬性的個數出現微小變化時,模型的預測能力顯著降低,穩健性不足,如決策樹的棵數為5,節點處分類屬性的個數分別55和57時,決定系數分別為0.81和0.83,下降了11.0%和8.4%(圖6a)。因此,淀粉隨機森林預測模型的決策樹的棵數和節點處分類屬性的個數分別為 38和70,為 0.90。同理,蔗糖、葡萄糖和果糖隨機模型決策樹的棵數分別為90,190和55,節點處分類屬性的個數則分別為52、10和18(圖6b、6c和6d)。
圖 7顯示了采用各自最佳決策樹的棵數和節點處分類屬性的個數建立的淀粉、蔗糖、葡萄糖和果糖隨機森林模型建模型和預測集的預測性能。分析采用98個葉綠素熒光參數構建的淀粉隨機森林模型可知(圖 7a),建模集和預測集的決定系數分別可達0.94和0.90,校正均方根誤差(Root Mean Square Error of Calibration Set, RMSEC)和預測均方根誤差(Root Mean Square Error of Prediction Set, RMSEP)分別為12.51和9.57μg/mm2,剩余預測偏差(Residual Predictive Deviation,RPD)可達3.43,說明模型可對柑橘葉片中淀粉含量具有極好的預測能力。分析蔗糖隨機森林模型的實測值和估測值(圖7b),建模集和預測集的決定系數和分別可達 0.96和0.84,均方根誤差分別為0.56和0.86μg/mm2,RPD可達2.50,由此說明該模型可以精確的反演柑橘葉片中的蔗糖含量。同理,葡萄糖的隨機森林模型預測結果為0.94、為 0.85、RMSEC 為 0.29μg/mm2、RMSEP 為 0.38μg/mm2和 RPD為 2.57(圖 7c)。相較于淀粉、蔗糖和葡萄糖,采用葉綠素熒光技術構建的果糖隨機森林模型預測性能預測略低,但也仍然具有較好的預測能力,模型的為 0.94、為 0.82、RMSEC 為 0.17μg/mm2、RMSEP為0.21μg/mm2和RPD為2.39(圖7d)。上述結果表明,葉綠素熒光參數能夠較好的表征葉片碳水化合物含量,進而說明葉綠素熒光技術具有快速診斷柑橘黃龍病的潛力。

圖7 實測值與隨機森林模型預測值分布Fig.7 Distribution of measured values versus predicted values using random forest model
2.5 柑橘黃龍病診斷模型的構建
黃龍病病原菌引起宿主碳水化合物代謝異常,導致淀粉、蔗糖、葡萄糖和果糖的過量累積,損傷了宿主PSII反應中心,破壞了葉綠體的類囊體膜結構,減少宿主光系統II中有活性的光反應中心數量,導致宿主的光合作用效率降低。表2分別顯示了基于98個葉綠素熒光參數和碳水化合物代謝參數建立的隨機森林模型總體識別正確率。基于葉綠素熒光參數建立的隨機森林模型建模集和預測集的識別正確率分別為100%和97.5%。采用淀粉、蔗糖、葡萄糖和果糖建立的隨機森林模型建模集和預測集的識別正確率則分別為100%和98.75%。兩種參數建立的判別模型對柑橘黃龍病的總體識別正確率相當,說明葉綠素熒光成像技術可用于柑橘黃龍病的快速診斷。

表2 基于葉綠素熒光和碳水化合物代謝參數的隨機森林模型總體識別正確率Table 2 Overall accuracies of random forest models based on chlorophyll fluorescence and carbohydrate metabolism%
3 結 論
本研究以不同染病程度柑橘葉片為研究對象,從碳水化合物代謝水平上分析探究宿主對病原菌侵染的響應,結合葉綠素熒光成像技術分析了黃龍病病原菌對宿主光合作用的影響,并建立柑橘黃龍病快速診斷模型。結果表明,病原菌致使宿主葉片淀粉、蔗糖、葡萄糖和果糖異常累積。病原菌損傷了宿主的光系統 II(Photosystem II, PSII)反應中心,改變了熒光動力學特性,減少宿主光系統II中有活性的光反應中心數量,導致宿主的光合作用效率降低,降低了宿主將吸收光能轉化為光化學反應的能力。葉綠素熒光參數與葉片的淀粉、蔗糖、葡萄糖和果糖具有明顯的相關性。最后,采用葉綠素參數建立了隨機森林快速判別模型,實現了97.50%識別正確率。研究結果可為田間柑橘黃龍病的防治提供數據支撐和技術指導。