石楠銹孢銹菌重寄生菌的鑒定及產毒培養基篩選
2020-08-04 03:02:22隋國強張登云孔磊陳玉惠李靖
生物技術通報 2020年7期
隋國強 張登云 孔磊 陳玉惠 李靖
(云南省林木生物技術重點實驗室 西南林業大學,昆明 650224)
重寄生現象(Mycoparasitism)是指自然界一種真菌寄生在另一種真菌上并完成生活史的現象。重寄生菌則是一類寄生在另一種真菌上的真菌[1]。重寄生作用作為植物病害的主要生防機制,越來越受到各國植物病理學家的重視。植物銹病的重寄生現象非常普遍,在生態自然控病和植物銹病生物防治中占有重要地位。以往研究主要集中在銹菌重寄生菌的篩選、鑒定及生物學特性等方面[2-4]。對于重寄生的機制目前認為主要包括酶和毒素兩方面[5]。真菌毒素(Mycotoxin)是由植物病原真菌及病原重寄生真菌產生的一類對寄主植物和病原真菌有毒的代謝產物,即真菌與寄主植物,重寄生菌與寄主真菌互作中的重要致病因子[6]。目前對于重寄生菌所產毒素的研究報道較少,多從重寄生菌對植物病原菌能產生很強的拮抗作用來推測其所產毒素的存在[7]。
擬盤多毛孢屬(Pestalotiopsis)含有對許多植物內生或致病的真菌。Steyaert[8]于1949年首先提出擬盤多毛孢屬,并根據分生孢子的形態特征將其與盤多毛孢屬(Pestalotia)分離。自此傳統擬盤多毛孢屬的分類鑒定主要依靠形態特征。但是依據形態特征對擬盤多毛孢進行分類有很大的局限性,無法明確種間界限及親緣關系;不同學者對同一物種的形態特征界定和看法不同,且主要以菌株的寄主植物命名,導致一個菌種多個名字,不利于擬盤多毛孢菌株的開發及利用[9]。近年來,分子手段被廣泛用于擬盤多毛孢屬的鑒定和分類。Jeewon等[10]基于ITS序列分析評估形態特征的系統發育重要性,使用ITS1-5.8S rRNA-ITS2識別物種。建立了一個系統發育樹,并顯示最終的形態特征是中性細胞的色素沉著和附肢尖端的形態。為進一步解決分類學問題,Liu等[11]使用形態學和ITS1-5.8S rRNA-ITS2和β-tubulin 2基因序列描述了一個新物種Pestalotiopsis hainanensis,并構建了相似的系統進化樹。Hu等[12]分析了來自ITS區域和β-微管蛋白基因的組合數據集DNA序列數據,發現組合基因更適合于解決分類學關系。
石楠銹孢銹菌(Aecidium pourthiaea)是石楠(Photinia prionophylla)葉銹病的病原菌,主要危害3年生以下的石楠幼樹及幼苗[13]。前期研究從患有銹病的球花石楠葉片上分離得到3株重寄生擬盤多毛孢,對3株菌的形態特征進行了描述并初步確定其為擬盤多毛孢屬,同時發現它們對石楠銹孢銹菌的銹孢子有很強的破壞作用[14]。本研究利用單基因及多基因復合分析并結合形態特征,對3株擬盤多毛孢進行種的鑒定,旨在為石楠銹病的生物防治提供菌種資源;同時篩選重寄生擬盤多毛孢的最佳產毒培養基,為石楠銹孢銹病的重寄生擬盤多毛孢毒素的分離鑒定及生物農藥的開發提供理論依據。
1 材料與方法
1.1 材料
1.1.1 菌株 3株重寄生擬盤多毛孢PG52、PG53和PG90分離自患有銹病的球花石楠葉片上,并保存于4℃[14]。茶藨生柱銹菌銹孢子采自云南省昆明市東川區二二二林場;石楠銹病銹菌孢子采自西南林業大學苗圃中。
1.1.2 培養基 6種產毒培養基:PDA培養基、改良 Fries培養基、查彼(Czapek)培養基、理查(Richard)培養基、PSKA培養基和改良M-1-D培養基[15]。
1.1.3 引 物 ITS5(5'-GAAAGTAAAAGTCGTAACAAG-3')和 ITS4(5'-TCCTCCGCTTATTGATAT-3');LROR(5'-ACCCGCTGAACTTAAGC-3')和 LR5(5'-TCCTGAGGGAAACTTCG-3');bt2a(5'-GGTAAACCA AATCGGTGCTGCTTTC-3')和 bt2b(5'-ACCCTCAGTG TAGTGACCCTTGGC-3')[14]。
1.2 方法
1.2.1 分子序列測定 從已培養8 d的PDA平板菌落邊緣挑取直徑為0.5 cm的擬盤多毛孢菌塊,以3塊/瓶的接種量接入50 mL PD培養液中,于25℃下以120 r/min振蕩培養5 d。每處理設置3個重復。發酵液雙層濾紙抽濾,無菌水清洗菌絲后利用DNA提取試劑盒(上海生工)進行擬盤多毛孢DNA的提取,以ITS5和ITS4為引物擴增ITS基因;以LROR和LR5為引物擴增28S基因;以bt2a和bt2b為引物擴增beta-tubulin基因,對擴增產物依次進行瓊脂糖凝膠電泳、膠回收、序列測定和GenBank數據庫檢索。
1.2.2 三株擬盤多毛孢分類鑒定 篩選合適的對比 序 列[16], 利 用 PAUP.40b10軟 件, 輸 入 paup程序構建系統發育樹。通過前期觀察得到3株擬盤多毛孢菌落形狀、顏色和分生孢子等形態特征[14],對比《中國真菌志:擬盤多毛孢屬》[17]和Maharachchikumbura等[16]文章中菌株的形態特征,通過分子鑒定和形態特征判斷3株擬盤多毛孢的具體種名。
1.2.3 粗提物的制備 將活化后的3株擬盤多毛孢,從各菌落邊緣挑取直徑0.5 cm的菌塊分別接入6種(PDA培養基、改良Fries培養基、查彼Czapek培養基、理查Richard培養基、PSKA培養基和改良M-1-D培養基)中,25℃培養20 d后提取毒素粗提物。對使用不同培養基培養后的3種重寄生擬盤多毛孢菌株及其培養基,分別切成均勻小塊,并放入相應標有菌株和培養基的錐形瓶中。然后加入有機溶劑(乙酸乙酯∶甲醇∶乙酸=80∶15∶5)100 mL浸泡3 d后過濾,重復3次,過濾后合并濾液。用旋轉蒸發儀在60℃下萃取,然后加入12 mL溶解劑二甲亞砜(DMSO)溶解提取物1-2 min既得粗提物,冰箱中低溫保存備用。
1.2.4 銹孢子活力檢測 生物測定采用凹玻片法,即在滅過菌的凹玻片內注入100 μL毒素原液,分別取適量茶藨生柱銹菌和石楠銹孢銹菌銹孢子浸于其中,攪拌使銹孢子與液體充分接觸。處理一段時間后,定時取少許銹孢子分別以蒸餾水和0.4%臺酚藍為浮載劑制片,于顯微鏡下觀察孢子的顏色和形態變化。以未接種菌塊的培養液處理的銹孢子為對照。
1.2.5 最佳產毒培養基的篩選 按上述方法進行生測,以添加銹孢子壁為誘導物的培養基制得的粗提物處理銹孢子作為平行組,以未接入菌塊的培養液處理銹孢子為陰性對照,每處理設3個重復。通過臺盼藍染色法測定銹孢子的活性,利用倒置顯微鏡(LEICA DMi1)拍照紀錄。
2 結果
2.1 三株擬盤多毛孢的分子鑒定
系統發育分析采用PAUP. 40b10軟件,以最大簡約(MP)法對3株擬盤多毛孢菌株和來自[14]文章中報道的菌株beta-tubulin(TUB)序列進行分析并構建系統發育樹,結果顯示(圖1):3株擬盤多毛孢中PG52和PG90聚在同一個分支,支持率為63%,兩者又與菌種Pestalotiopsis kenyana和Pestalotiopsis Oryzae在同一個大的分支,支持率為98%;PG53與PG52、PG90分支相距較遠,且與Pestalotiopsis telopeae在同一個分支,支持率為59%。表明PG52與PG90親緣關系近,兩者與Pestalotiopsis kenyana和Pestalotiopsis Oryzae的親緣關系較近,而PG53與Pestalotiopsis telopeae親緣關系較近。
以最大簡約(MP)法對3株擬盤多毛孢菌株的ITS、LSU和TUB復合序列進行分析并構建系統發育樹,結果顯示(圖2):3株擬盤多毛孢中PG52與PG90聚在同一個分支,支持率為61%,又與Pestalotiopsis kenyana和Pestalotiopsis Oryza在同一個大分支,支持率為98%;PG53與PG52、PG90分支相距較遠,且與Pestalotiopsis telopeae在同一個分支,支持率為61%。表明PG52與PG90親緣關系近,兩者與Pestalotiopsis kenyana和Pestalotiopsis Oryzae的親緣關系較近,而PG53與Pestalotiopsis telopeae親緣關系較近。
對比發現ITS、LSU和TUB復合序列構建系統發育樹的結果與TUB序列系統發育樹的結果相同,又根據3株擬盤多毛孢的菌落形態、分生孢子基附肢的數量、孢子顏色和長度等形態特征對比結果,可知PG53菌株的形態與Pestalotiopsis telopea的形態描述基本一致;PG52和PG90菌株的形態特征與Pestalotiopsis kenyana的形態描述一致。
2.2 三株擬盤多毛孢對石楠銹病銹菌孢子的作用
3 株擬盤多毛孢均能在銹孢子堆上生長,取處理后的銹孢子(圖3-B-D)用倒置顯微鏡觀察,與未處理的銹孢子(圖3-A)對比,結果發現處理后的銹孢子內含物濃縮,有的銹孢子內含物釋放到孢子壁外,形成空殼,但銹孢子壁基本完整,初步判定其為毒素的作用。
2.3 三株擬盤多毛孢產毒培養基的篩選結果
2.3.1 石楠銹病銹菌孢子 PG52菌株:改良Fries培養基(圖4-B)和改良M-1-D培養基(圖4-F)的粗提物效果最好,處理染色后,銹孢子內和銹孢子壁都能染上深藍色。理查培養基(圖4-D)和PDA培養基(圖4-A)的粗提物效果稍次,PSKA培養基(圖4-E)和查彼培養基(圖4-C)最次。

圖1 三株擬盤多毛孢PG52、PG53和PG90的TUB序列建樹結果
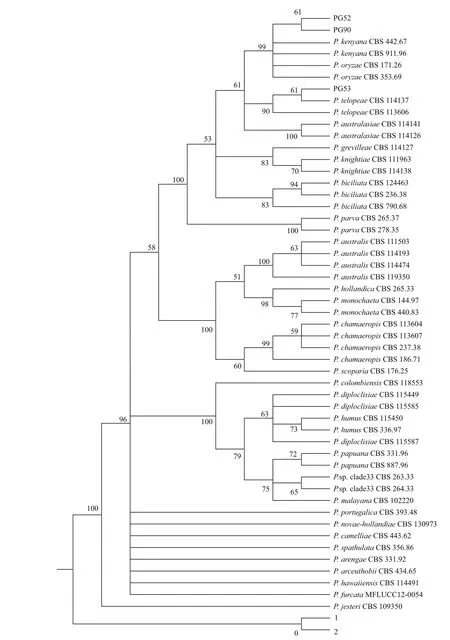
圖2 三株擬盤多毛孢PG52、PG53、PG90的三段序列組合建樹結果
PG53菌株:改良Fries培養基和改良M-1-D培養基的粗提物效果最好,處理染色后,銹孢子內和銹孢子壁都能染上深藍色。理查培養基和PSKA培養基稍次,PDA培養基和查彼培養基最次。

圖3 三株擬盤多毛孢對石楠銹病銹菌孢子的作用
PG90菌株:改良Fries培養基、理查培養基、查彼培養基和改良M-1-D培養基的粗提物效果最好,處理染色后,銹孢子內和銹孢子壁都能染上深藍色。PDA培養基和PSKA培養基效果稍次。
2.3.2 茶藨生柱銹菌孢子 PG52菌株:理查培養基(圖5-D)和改良Fries培養基(圖5-B)的粗提物效果最好,處理染色后,銹孢子內和銹孢子壁都能染上深藍色。PSKA培養基(圖5-E)和改良M-1-D培養基(圖5-F)的粗提物效果稍次。而PDA培養基(圖5-A)和查彼培養基(圖5-C)的粗提物則沒有效果,銹孢子仍然呈現黃色。
PG53菌株:理查培養基、改良Fries培養基和改良M-1-D培養基的粗提物效果最好,處理染色后,銹孢子內和銹孢子壁都能染上深藍色。PDA培養基的粗提物效果稍次,染色顏色不深。而PSKA培養基和查彼培養基的粗提物則沒有效果,銹孢子仍然呈現黃色。
PG90菌株:理查培養基、查彼培養基、改良Fries培養基和改良M-1-D培養基的粗提物效果最好,處理染色后,銹孢子內和銹孢子壁都能染上深藍色。PDA培養基和PSKA培養基的粗提物效果稍次,染色顏色不深。該菌株沒有無效果的粗提物。

圖4 PG52菌株不同培養基粗提物處理石楠銹病銹菌孢子效果圖

圖5 PG52菌株不同培養基粗提物處理茶藨生柱銹菌銹孢子效果圖
3 討論
近年將分子系統學方法用于擬盤多毛孢分類中,用多個基因片段分別或共同建立分子系統樹,并結合形態學特征可以在一定程度上解決目前在形態分類上所遇到的問題。擬盤多毛孢屬的分生孢子在形態上是多種多樣的,基于單基因和多基因系統發育,以及形態特征,宿主關聯和地理分布等更加準確的鑒定出物種[18]。Jeewon等[19]在對擬盤多毛孢的第一個包容性系統發育研究中利用ITS序列數據,以評估分類學中擬盤多毛孢的形態學特征的系統發生意義。Hu等[12]在區分華山松和Ribesspp. 中的內生擬盤多毛孢種類時指出,TUB基因更好地解決了擬盤多毛孢的系統發育。TUB和ITS基因的組合可提供更好的系統發育分辨率,并建議將它們用于解決擬盤多毛孢的系統發育。Maharachchikumbura等[16]在擬盤多毛孢菌中測試了肌動蛋白、鈣調蛋白、谷氨酰胺合酶、3-磷酸甘油醛脫氫酶、ITS、LSU、18S nrDNA、RNA聚合酶II、TEF和TUB等,通過比較發現,分子鑒定效果優于形態學方法。在本研究中,基于ITS、TUB和LSU基因區域組合的系統發育物種識別在末端進化枝上提供了強力支持,為更好的鑒定擬盤多毛孢菌株奠定理論基礎。
銹菌是一類重要的植物病原菌,每年都給農業、林業生產造成巨大的損失[20-21]。防治銹病的方法包括化學防治、物理防治、生物防治和采用轉基因的手段,這些方法各有利弊[22]。近年來,對擬盤多毛孢致病機理的研究包括對其侵入寄主植物產生的相關酶的研究,及其所產毒素及在生產上的應用的前景等方面的研究[23-25]。目前,由石楠銹孢銹菌(Aecidium pourthiaeasyd.)引起的石楠葉銹病發病逐年加重,能引起石楠葉片、嫩梢的失綠和枯萎死亡,進而引起一些次生病蟲害的發生,昆明因氣候溫暖,該病害全年都有發生,嚴重影響了石楠的綠化和觀賞價值。從石楠銹孢銹菌孢子分離得到的3株重寄生擬盤多毛孢,通過研究證明,對石楠銹病銹菌孢子和茶藨生柱銹菌孢子均有強寄生作用,結合茶藨生柱銹菌孢子和石楠銹病銹菌孢子的實驗結果確定了3株擬盤多毛孢的最佳產毒培養基。研究結果為重寄生擬盤多毛孢在銹病生物防治中的應用及其次生代謝產物的研究奠定了良好的基礎。
4 結論
本研究分別利用ITS、LSU、TUB以及三者組合構建系統發育樹,結果顯示,PG52、PG90與Pestalotiopsis kenyana和Pestalotiopsis Oryza菌株在同一個分支,PG53為Pestalotiopsis telopeae菌株;同時結合形態特征,最終鑒定PG52、PG90為Pestalotiopsis kenyana菌 株,PG53為Pestalotiopsis telopeae菌株。分離得到的3株石楠銹孢銹菌重寄生擬盤多毛孢,其不僅能破壞石楠葉銹菌的銹孢子,也能破壞茶藨生柱銹菌的銹孢子,均使銹孢子的內含物溢出,導致銹孢子死亡。但因孢子壁基本完整,初步判定其為毒素的作用。PG52菌株的最佳產毒培養基為改良Fries培養基;PG53菌株的最佳產毒培養基為改良M-1-D培養基;PG90菌株的最佳產毒培養基為改良M-1-D培養基。
