利用同源重組技術構建Hoxa5、BMP6真核表達載體及共轉染成纖維細胞表達
2020-08-31 03:12:20張夢瑤劉開東賀建寧
華北農學報 2020年4期
關鍵詞:信號
李 晶,張夢瑤,劉開東,柳 楠,賀建寧
(1.青島農業大學 動物科技學院,山東 青島 266109;2.曲阜市畜牧獸醫技術服務中心,山東 曲阜 273100;3.青島畜牧獸醫研究所,山東 青島 266121)
敖漢細毛羊是我國很早培育出的細毛羊品種,它主要分布在東北部地區,是我國最具有代表性的細毛羊之一[1],其羊毛是紡織工業的重要原料。敖漢細毛羊毛囊的結構和特性直接決定毛發品質和商業價值[2],毛囊擁有著相對復雜的結構,它的形成是由多種細胞來協同完成的。毛囊通過一系列的分化生長最后成為羊毛,其生長發育也有些復雜,它是由多種因子來相互調節,因此,通過研究相關因子來探討毛囊的生長發育顯得尤為重要。
Hox家族在生物體內是一類與發育調控相關十分重要的基因。Hox家族這類轉錄因子都是高度保守的[3],對毛囊細胞增殖分化也起著十分重要的作用[4-5]。研究發現,在皮膚和毛囊生長發育過程中Hox家族基因起到了主導作用[6],Hoxa5主導了皮膚細胞增殖、分化與凋亡[5],對毛囊退行期進行了調控,并誘導了細胞分化[7]。動物形態的進化和多樣性與Hox基因數目、序列及其表達方式密切相關,已有研究結果表明,當Hox家族基因發生變化時,動物在被毛生長時會表現出缺陷[8]。BMP6基因是TGF-β超家族中的一員[9-10],TGF通路中的許多轉錄因子都受BMP6調控,均表現出上調和下調趨勢。楊燕燕等[11]在生殖發育上對BMP6進行了研究,它控制著羊的多胎,尤其是在發情期基因表達相當明顯。也有研究表明,BMPs通路中的轉錄組因子對毛囊發育主要表現出抑制作用,而它的下游靶標BMPRs主要表現出促進的作用[12-13]。程旭等[14]通過對鐵調素在BMP6-HJV-SMAD信號通路的作用,驗證了BMP6的重要性。由此可見,Hoxa5和BMP6基因都能抑制毛囊生長,但二者之間的關系鮮有報道。
對于家畜皮膚毛囊的一些研究大多集中在單個基因的功能驗證上,還有一些是與骨的調節、繁殖等與皮膚毛囊無關的研究,本試驗主要的側重點是利用同源重組技術構建表達載體通過與單、共轉染比較基因表達量的變化來研究Hoxa5和BMP6之間的作用關系,旨在為更深層次上研究毛囊功能基因奠定基礎。
1 材料和方法
1.1 試驗材料
健康的30日齡胎羊由青島畜牧所奧特種羊場提供;TRIzol、KpnⅠ、倒置顯微鏡、熒光定量PCR等均購自Bio-Rad公司[9]。
1.2 試驗方法
1.2.1 引物設計 選取綿羊Hoxa5基因(GenBank登錄號為NM-001009431.1)CDS區 和BMP6基因(GenBank登錄號為NM-001110277.1)CDS區,軟件導入pcDNA3.1表達載體的序列,通過Snap Gene設計Hoxa5基因和BMP6基因上下游引物,序列分別如下:
F(Hoxa5):5′-ATGCGTTAACTCAGGAATCGGATGCATCGGTCAAGCTGGCAT-3′;R(Hoxa5):5′-TGGCCTTAAAATCTCCGAGGAAATACTCGCTCACGCGGTAG-3′。F(BMP6):5′-TTACGGGCATCGGTAGGGAAATTCGTATTGCATGCAAAGCT-3′;R(BMP6):5′-TGTCCGAGACCTGCACATCCACGGCGGCCCTTCAGCATACCCCTA-3′。擴增體系:20 μL,cDNA 1 μL,上下游引物各1 μL,Mix 17 μL。
1.2.2 pcDNA3.1與Hoxa5、BMP6基因同源重組 對Hoxa5、BMP6進行切膠,用KpnⅠ對pcDNA3.1進行單酶切,體系:KpnⅠ 1 μL,pcDNA3.1 5 μL,Buffer 5 μL,ddH2O 39 μL。通過電泳對BMP6、Hoxa5基因片段及pcDNA3.1進行膠回收,SoSo連接體系:BMP6、Hoxa5基因上、下游引物各1 μL,pcDNA3.1 1 μL,SoSo 5 μL,ddH2O 3 μL,轉化DH5α,過夜對菌液進行振蕩培養,將菌液生長良好的送往公司進行測序[12-13]。
1.2.3 胎兒成纖維細胞培養 在細胞室將30日齡胎羊從母羊子宮內取出,先用75%的酒精對胎兒進行消毒,再用PBS將胎兒表面的酒精沖洗掉。在培養皿中用手術刀將胎兒的頭部和四肢、內臟等全部剔除,只用胎羊的軀干,將其剪碎,分布在培養皿中,在每一個肌肉塊上面加2滴已經預熱的胎牛血清,放進CO2培養箱(37 ℃,5.0% CO2),24 h對其進行觀察并且換液[15-16]。
1.2.4 細胞轉染 細胞進行傳代以后,等密度再次達到95%左右時,借助轉染試劑對重組的載體進行瞬時轉染。取脂質體20 μL,PBS 480 μL,加入到無菌的離心管中并混勻,靜置5 min。進一步進行轉染,轉染時分為2組,分別是共轉染組和單轉染組,共轉染組中加入Hoxa5、BMP6質粒各20 μL,單轉染組加入Hoxa5質粒40 μL或者BMP6質粒40 μL,2組均加入PBS 460 μL,具體詳細操作步驟參照Lipofectamine2000說明書[17]。
1.2.5 細胞轉染后的qPCR 重組載體轉染成纖維細胞后需要進一步的增殖培養,當密度達到95%左右時,對成纖維細胞的RNA進行提取,并將其反轉錄成cDNA[18]。使用Snap Gene軟件對Hoxa5、BMP6和GAPDH基因進行qPCR引物設計,序列見表1。
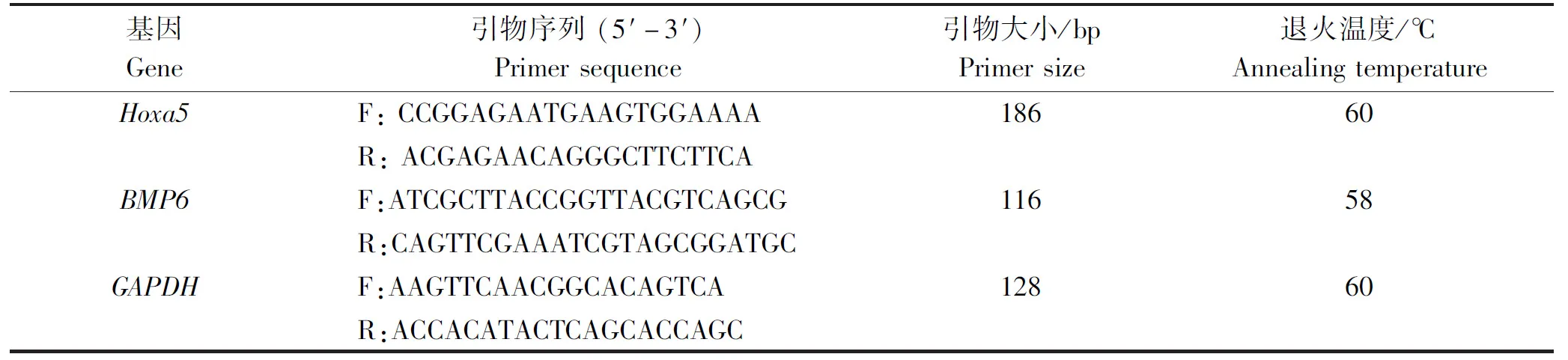
表1 引物信息Tab.1 Primers information
qPCR反應條件:95 ℃反應7 min;95 ℃ 40 s,60 ℃退火40 s,35個循環;72 ℃ 30 s,4 ℃保存。每組3個生物學重復,對數據利用Ct(2-ΔΔCt)公式進行計算,得出Hoxa5、BMP6的相對表達量。借助SPSS 20.0軟件對數據進行差異顯著性的分析,以P<0.01作為差異顯著性判斷標準[17]。
1.2.6 Hoxa5、BMP6蛋白表達的檢測 提取成纖維細胞中的組蛋白,進行Western Blot,首先進行轉膜,后進行封閉,時長2 h;進行一抗、二抗的孵育,均對抗體進行2 000倍的稀釋。孵育完成后利用1∶1的曝光液進行曝光拍照[17]。
2 結果與分析
2.1 Hoxa5、BMP6基因PCR擴增
以cDNA為模板,添加引物進行PCR 擴增獲取Hoxa5、BMP6基因片段。1%瓊脂糖凝膠電泳結果表明(圖1),Hoxa5基因條帶在圖中804 bp處,BMP6基因條帶在圖中537 bp處,都與目的基因CDS區大小吻合,可用于后期的試驗。
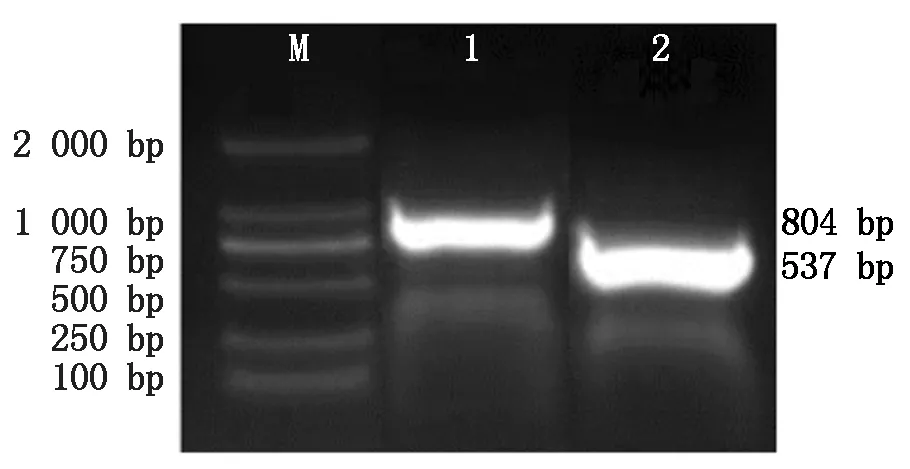
M.2000 Marker;1.Hoxa5基因擴增片段(804 bp);2.BMP6基因擴增片段(537 bp)。M.2000 Marker;1.Hoxa5 product(804 bp);2.BMP6 product(537 bp).
2.2 pcDNA3.1表達載體酶切純化
使用KpnⅠ將pcDNA3.1表達載體從環狀酶切成鏈狀,用1%瓊脂糖凝膠電泳檢測是否酶切成功(圖2)。1,2是對pcDNA3.1的KpnⅠ單酶切,pcDNA3.1長度為5 428 bp,圖中條帶所在位置約是5 428 bp,結果顯示酶切完成,回收條帶進行重組。
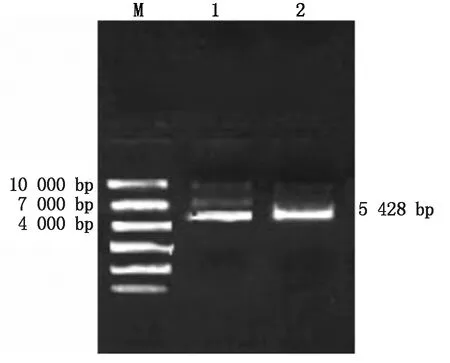
圖2 pcDNA3.1載體單酶切鑒定Fig.2 Single enzyme digestion identification of pcDNA3.1 vector
2.3 pcDNA3.1表達載體的重組及鑒定
將pcDNA3.1酶切成鏈狀后,用SoSo試劑盒將BMP6、Hoxa5分別與pcDNA3.1相連接,轉化,振蕩培養,進行菌液PCR驗證如圖3所示,在約804 bp處有明亮電泳條帶,是Hoxa5基因;537 bp處的是BMP6基因。菌液測序結果用Blast進行對比,結果如圖4,說明BMP6、Hoxa5沒有堿基的突變,已成功的連接到pcDNA3.1載體上。將連接好的重組載體從菌液中提取出來,如圖5所示,1-2為pcDNA3.1-BMP6重組質粒,大小為5 965 bp,3-4為pcDNA3.1-Hoxa5重組質粒,大小為6 232 bp,與圖中條帶相符。
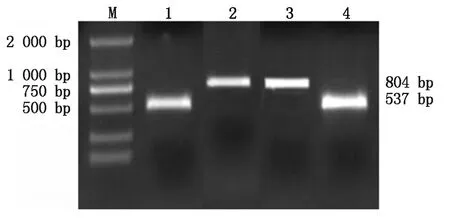
M.2000 Marker;1,4.BMP6菌液PCR;2,3.Hoxa5菌液PCR。M.2000 Marker;1,4.BMP6 bacterial PCR;2,3. Hoxa5 bacterial PCR.
2.4 細胞培養觀察
對胎兒成纖維細胞的生長情況進行觀察記錄(圖6),24 h能夠觀察到細胞已經開始貼到培養皿底部,周圍也有一些漂浮的組織。培養至96 h后,密度再次達到95%左右,此時即可對原代細胞進行傳代[18]。
2.5 RT-PCR檢測Hoxa5、BMP6基因mRNA表達
對引物進行驗證,1%瓊脂糖凝膠電泳結果(圖7)表明,引物擴增效果良好,沒有出現二聚體等雜物,在128 bp 處的是內參基因GAPDH,在186 bp處的是Hoxa5基因,在116 bp處的是BMP6基因,綜上可得引物均可用于實時熒光定量PCR試驗。

圖4 BMP6(上)和Hoxa5(下)表達載體菌液的測序比對Fig.4 Sequencing comparison of BMP6 (up) and Hoxa5 (down) expression vector bacilli
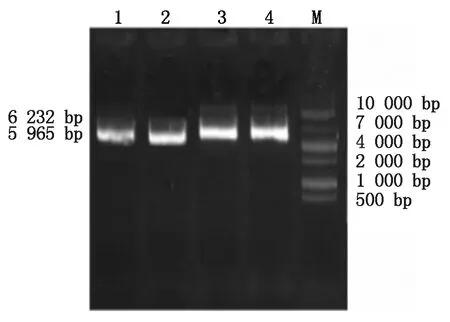
M.10000 Marker;1-2.pcDNA3.1-BMPR6;3-4.pcDNA3.1-Hoxa5。M. 10000 Marker;1-2. pcDNA3.1-BMPR6 recombinant plasmids;3-4. pcDNA3.1-Hoxa5 recombinant plasmids
圖 8,9分別為Hoxa5、BMP6和GAPDH的qPCR過程中的擴增曲線、熔解曲線,兩線效果顯示均良好,在qPCR過程中沒有出現非特異性產物。對2組的mRNA表達進行差異顯著性分析(圖10),分析顯示Hoxa5與BMP6共轉染后的Hoxa5基因相對表達量極顯著低于單轉染(P<0.01),而共轉染后的BMP6基因相對表達量在共轉染卻極顯著高于單轉染(P<0.01)。
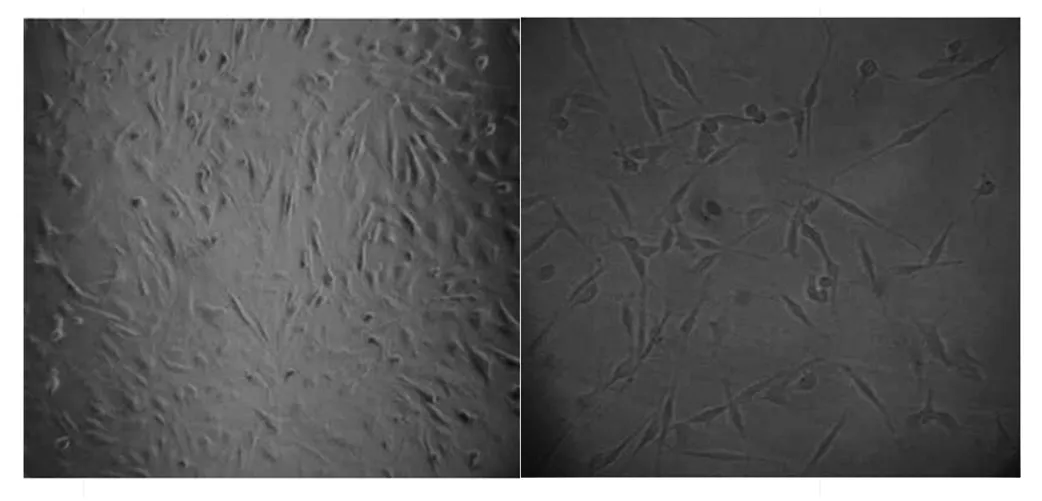
圖6 成纖維細胞原代(左)、傳代(右)培養(×100)Fig.6 Primary (left) and passage (right) culture of fibroblasts (×100)
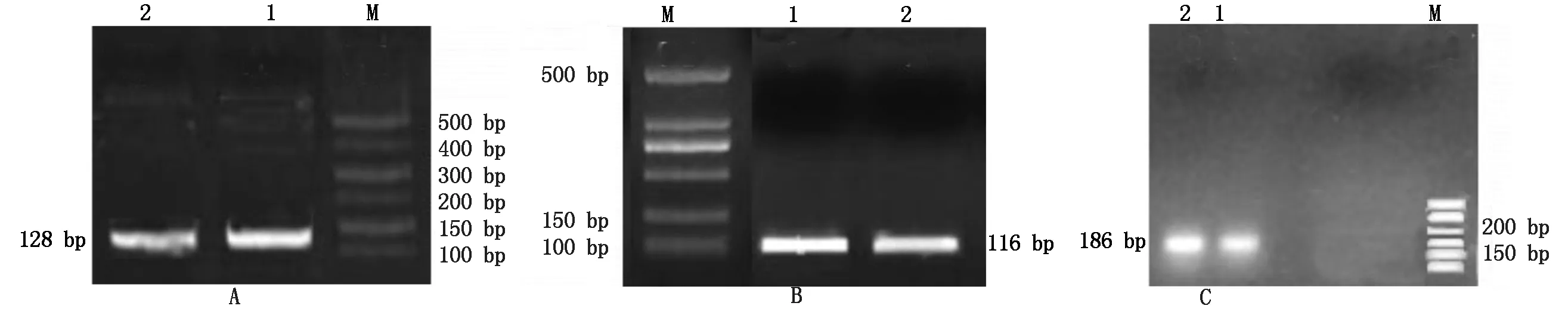
A:M.500 Marker;1-2.內參基因GAPDH;B:M.500 Marker;1-2.目的基因BMP6;C:M.500 Marker;1-2.目的基因Hoxa5。A:M.500 Marker;1-2 .Reference gene GAPDH;B:M. 500 Marker;1-2.Target gene BMP6;C:M.500 Marker;1-2. Target gene Hoxa5.
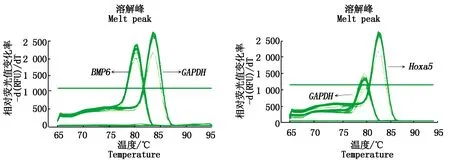
圖8 Hoxa5、BMP6和GAPDH 熔解曲線Fig.8 Hoxa5,BMP6 and GAPDH melting curves
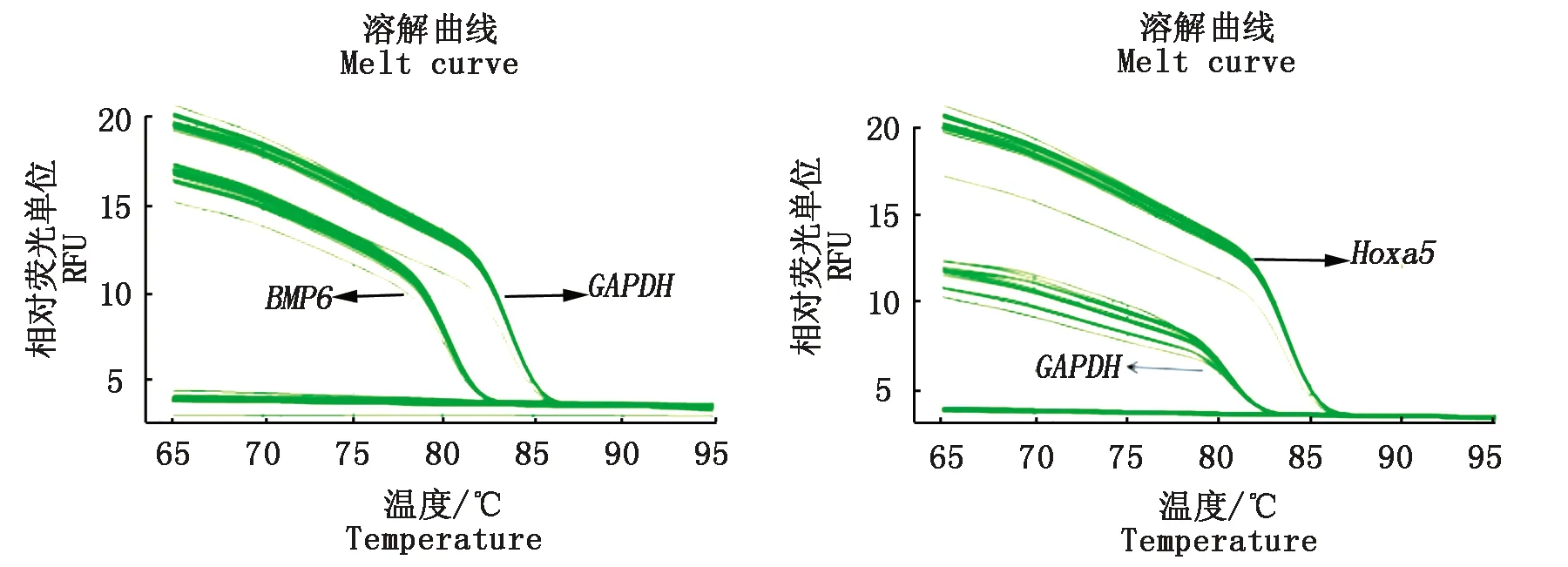
圖9 Hoxa5、BMP6和GAPDH擴增曲線 Fig.9 Hoxa5, BMP6 and GAPDH amplification curve
2.6 Hoxa5、BMP6蛋白表達的檢測
對Hoxa5、BMP6基因的共轉染、單轉染蛋白的表達(圖11)進行比較分析,從灰暗度以及條帶的大小,使用ImageJ和SPSS軟件進行分析,Hoxa5與BMP6共轉染后的Hoxa5蛋白表達量極顯著低于單轉染過程(P<0.01),而共轉染的BMP6蛋白表達量卻極顯著高于單轉染過程(P<0.01)(圖12),這與mRNA相對表達量的結果一致。
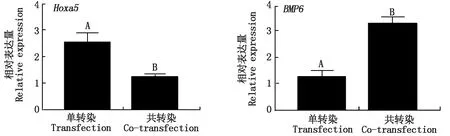
不同大寫字母表示差異極顯著(P<0.01)。圖12同。Different capital letters indicate extremely significant difference(P<0.01).The same as Fig.12.
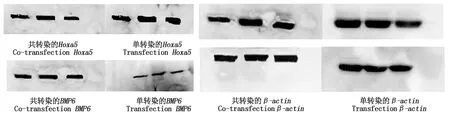
圖11 Hoxa5、BMP6蛋白表達 Fig.11 Hoxa5 and BMP6 expression protein
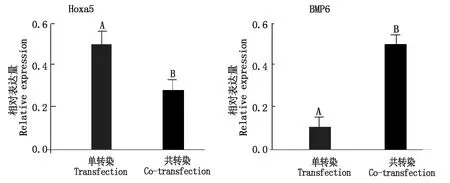
圖12 Hoxa5、BMP6蛋白相對表達量Fig.12 Relative expression of Hoxa5 and BMP6 protein
3 討論
許多研究表明,毛囊的發育依靠一系列的信號分子,這些信號分子在真皮細胞和上皮細胞間穿梭著,介導了這2個細胞群體間的相互作用,毛囊能夠穩定的進行發育全靠這兩者的介導。毛囊的形成過程中離不開所謂的原始信號,此信號來源于真皮下[18],并且在真皮下去激活Wnt信號[19-20]。目前,許多信號通路都參與毛囊的發育,最常見的有Wnt通路、FGF家族、BMP家族、TGF家族[21]等。Botchkarev等[22]利用小鼠來研究BMPs在皮膚中的表達位置以及表達時期,表明BMPs家族基因對毛囊的發育存在著一定的抑制作用。BMP6基因和BMP4一樣同屬于BMP家族,是一種抑制毛囊發育的信號分子,Hoxa5基因是TNF家族中尤為重要的基因,同樣也是一種抑制毛囊發育的信號分子,兩基因對毛囊的發育均有抑制作用[23]。在信號通路中,BMP家族中的信號分子對TNF通路中信號分子有干涉作用,有些是輔助基因的表達,有些是抑制基因的表達。現在對單個基因功能的驗證普遍存在,多個基因相互作用還是尤為少見,本研究通過利用同源重組手段來驗證多基因相互之間的作用,幫助篩選相關基因,有助于后期育種和羊毛生產。
為探討Hoxa5、BMP6基因在細胞水平上是否存在互作效應,本研究利用同源重組技術,快速構建了pc-DNA3.1-Hoxa5、pc-DNA3.1-BMP6表達載體,并通過共轉染、單轉染后基因mRNA、蛋白表達量的比較分析,發現Hoxa5基因有促進BMP6基因表達,BMP6基因有抑制Hoxa5基因表達的現象。本研究在構建表達載體過程中所用同源重組技術較傳統的T克隆技術節約了時間,減少了堿基突變;同時由分子水平到細胞水平得到了Hoxa5和BMP6基因的互作關系,一方面驗證了與羊毛生長有關的基因,為信號通路的解析添加了論證,另一方面為進一步在個體水平上驗證其功能奠定基礎。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
