珠江口黃茅海底質沉積中介形類的生態類型及其分布特征
2020-09-10 15:43:10黃毅吳潔殷征欣關瑤倪錦鋒
海洋開發與管理 2020年6期
黃毅 吳潔 殷征欣 關瑤 倪錦鋒


摘要:文章通過對黃茅海底質沉積樣品中的介形類生態類型進行研究,探討其空間分布特征。研究結果顯示:研究區內出現的介形類可劃分為4種生態類型,總體表現為以近岸廣鹽型分子占主導地位,優勢度極高;淺海多鹽-真鹽型分子次之;而廣海真鹽型和淡水-微鹽型分子豐度很低。根據黃茅海河口灣的水體條件,可大致將研究區劃分為5種亞環境區,各區內的介形類生態類型組成也存在較大差別,表現出不同的空間分布特征。介形類分布受海水鹽度控制最大,其次為水動力條件。在水體鹽度小、水動力條件強的河流入海口及其影響區域,介形類豐度小,且生態類型單一;而在黃茅海河口灣南部海域,由于海水底層鹽度全年在15‰以上,水動力條件適宜,介形類豐度總體較高,生態類型增多。
關鍵詞:珠江口;黃茅海河口灣;介形類;生態類型;分布特征
Abstract:In this study,the ecological types of ostracod in bottom sediments of Huangmaohai estuary were analyzed to discuss their spatial distribution characteristics.The results showed that the ostracod could be divided into four ecological types in the study area.The dominant ecological type is the coastal euryhaline type,followed by the marine poly euhaline type.Besides,the abundance of the marine euhaline type and the freshwater? slightly brackish type are very low.According to the water conditions of Huangmaohai estuary,the study area could be divided into 5 sub-environmental areas,and the composition of the ecological types of ostracod in each area is also quite different,showing the different spatial distribution characteristics.The results showed that the distribution of ostracod is mainly controlled by the salinity of seawater,followed by the hydrodynamic condition.In the estuaries of river with low salinity and strong hydrodynamic conditions,the abundance of ostracod is low and the ecological type is single.But the area in the southern estuary is different which shows the higher abundance and more ecological types of ostracod because of the bottom salinity of seawater is above 15‰ all year and the hydrodynamic conditions are suitable.
Key words:Pearl River Estuary,Huangmaohai Estuary,Ostracod,Ecological types,Distribution characteristics
微體動物介形類是生存環境范圍較廣的甲殼動物,常可見于海水、半咸水和淡水環境。介形類因其能敏感地反映水體鹽度、溫度、水動力條件、底質等環境參數[1],在地質學尤其是在分析海陸交互的河口地區環境上,具有重要的研究意義。因此,微體動物中除了有孔蟲繼續被充分研究和利用外,基于微體動物介形類的研究方法也成為探討河口地帶環境變化的手段之一。
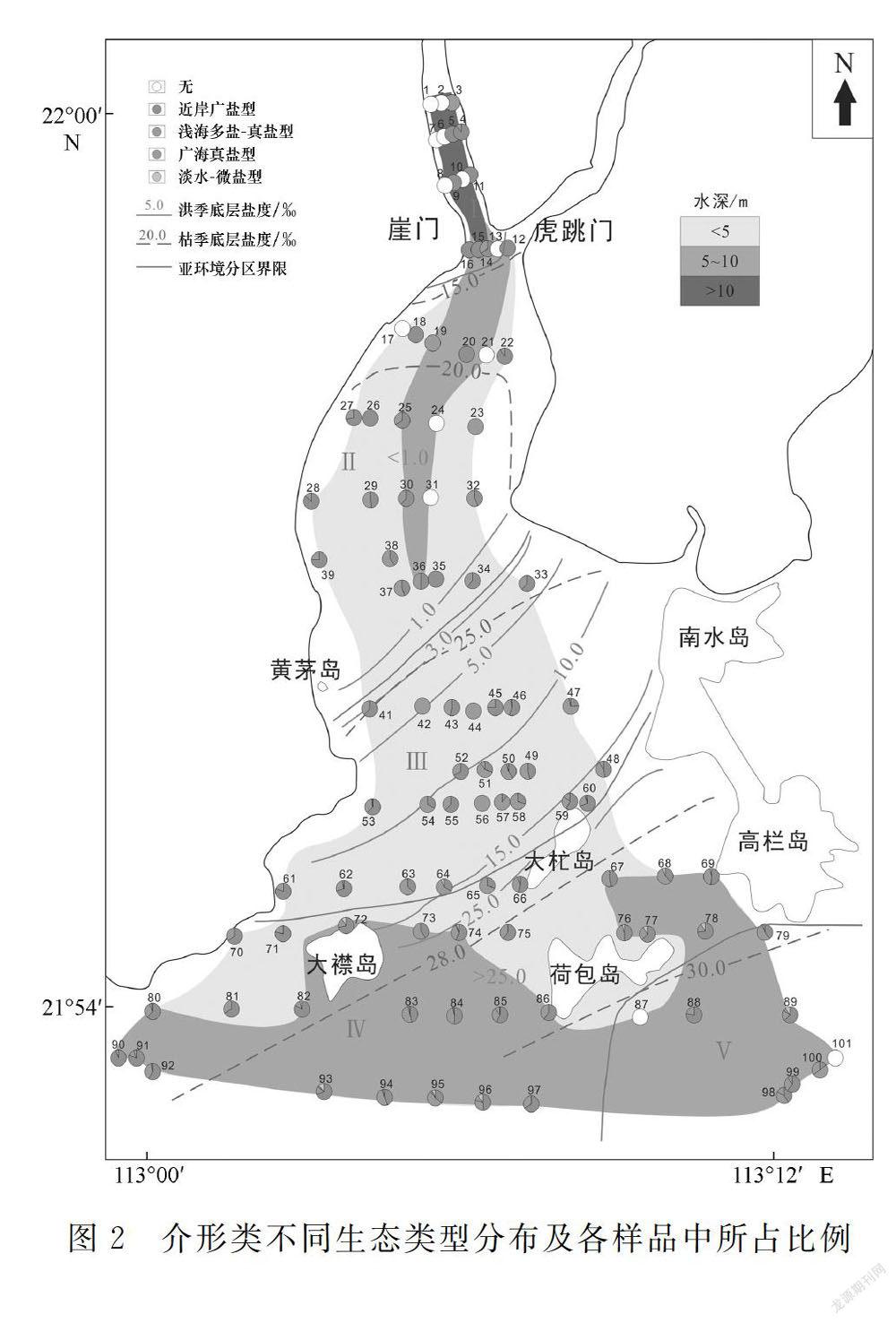
近年來對珠江口地區介形類微體動物的研究前人已有涉及[2-12],主要集中在伶仃洋海域表層沉積物或者三角洲鉆孔樣品。吳潔等[13]利用黃茅海有孔蟲和介形類的去趨勢對應分析,對控制微體動物分布的主要環境因素進行了初步研究。本研究在此基礎上,對黃茅海的底質沉積樣品中的介形類進行進一步分析,主要側重于介形類生態類型及其空間分布特征。底質沉積樣品取樣站位如圖1所示,具體取樣和樣品處理過程參見文獻[13],鹽度參見文獻[14]。
1 研究區概況
珠江擁有世界上最復雜的河口系統,匯集西江、北江和東江等河網水系后,通過珠江口獨特的8個口門入海[15]。珠江口發育有2個喇叭形河口灣,分別為東部的伶仃洋和西部的黃茅海,其中黃茅海灣頂端有虎跳門和崖門2個口門匯入,伶仃洋有4個口門(虎門、蕉門、洪奇瀝、橫門)匯入,而磨刀門和雞啼門直接注入南海[16]。
黃茅海河口灣三面環山,呈NNW-SSE走向,海域面積約543 km2[15]。該海域內島嶼眾多,通過島嶼峽口與南海相連[17],灘與槽交錯,西灘、東灘、攔門沙淺灘的水深均在5 m以下,北部主槽、大襟島和荷包島之間的中口深槽的水深基本大于5 m,形成了黃茅海海域“三灘兩槽”的水底地形格局[15,18]。
黃茅海河口灣屬于潮流作用優勢型河口,其潮汐為不規則半日潮性質,潮差由灣口向灣頂遞減[19-22],并具有“東進西出”的近岸水流格局[23]。由于受到熱帶海洋性季風氣候影響,河口地區的降雨量和徑流量呈現明顯的季節性,導致河口灣海域海水的鹽度值也具有顯著的季節性波動[24]。黃方等[14]研究顯示,黃茅海河口灣水體鹽度具有明顯的季節性變化,枯水期海水上涌,灣頂口門處鹽度可達15‰;豐水期受徑流下泄影響,黃茅海灣內北部鹽度較低。受黃茅海河口灣水流“東進西出”的影響,灣內的等鹽度線整體呈NE-SW走向。研究發現,黃茅海河口灣北部主槽的表層沉積物中砂含量最高,其次為東口,中口深槽含砂量中等,攔門沙淺灘次之,河口灣東西兩側的淺灘砂含量最低,表明該地區不同水域水動力條件具有明顯的差異[13]。
根據珠江口黃茅海海域鹽度、水深、水動力環境等方面的差異,可大致將該海域劃分為5個亞環境區(圖1)。①Ⅰ區位于河口灣頂,河流入灣處,沿主槽水深10 m以上,主要受徑流作用影響,水動力較強,枯季鹽度小于15‰,洪季底層水鹽度小于1‰。②Ⅱ區位于河口灣北部,攔門沙淺灘以北,枯季底層鹽度15‰~25‰、洪季底層鹽度小于3‰,受徑流影響強于潮流作用,沿主槽水動力條件仍很強,水深5~10 m,東灘和西灘水深1~3 m。③Ⅲ區位于攔門沙淺灘海域,潮流上溯流與徑流下泄流作用的交匯區,水動力條件相對較弱,水深3 m,兩側淺灘水深略淺,枯季底層鹽度25‰~27‰,洪季底層鹽度3‰~15‰。④Ⅳ區大致位于大襟島—大杧島一線以南,黃茅海灣口的南部海域,主要受潮流作用影響,枯季底層鹽度26‰~30‰,洪季底層鹽度15‰~25‰,水深5~10 m,近島嶼和陸地處水深較淺。⑤Ⅴ區位于黃茅海灣東口,水深通常大于10 m,潮水上溯流占優勢,且水動力條件較強,枯季底層鹽度大于30‰,洪季底層鹽度大于25‰。
2 黃茅海介形類生態類型
研究區底質沉積中共鑒定出介形類16屬20種,其中Neomonoceratina delicata、Sinocytheridea impressa和Bicornucythere bisanensis這3個屬種占比均大于17%,屬于優勢屬種,其余屬種優勢度較低,各屬種占比均小于6%,部分屬種僅零星出現,各屬種相對含量見表1。Liu等[25]曾對珠江口地區第四紀介形類的生態類型進行分類,本研究亦采用此分類方法進行劃分和討論。
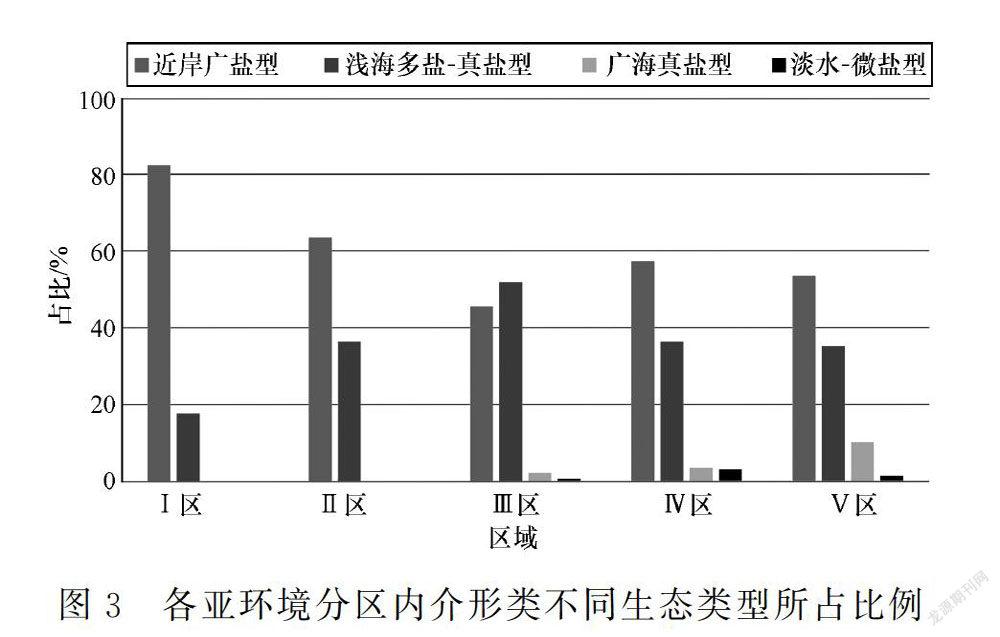
受黃茅海河口灣多變環境的影響,該地區的介形類總體表現為以近岸廣鹽型分子占主導地位,優勢度極高;淺海多鹽-真鹽型分子次之;而廣海真鹽型和淡水-微鹽型分子豐度很低。
(1)近岸廣鹽型:該類型的介形類可適應鹽度2‰至真鹽,甚至高鹽條件,鹽度范圍最廣,多生存于小于20 m水深的沿岸水域,是黃茅海河口灣最為豐富的介形類。共發現5種,其殼體可占總數的54.92%。其中豐度最高的屬種是Neomonoceratina delica和Sinocytheridea impressa。僅此2個種的殼體就占這一類型的99%以上。此外,還零星出現Bicorncythere leizhouensis、Neomonoceratina dongtaiensis、Leptocythere ventriclivosa等屬種。
(2)淺海多鹽-真鹽型:這一類型介形類能適應相對較廣的鹽度范圍,多分布于多鹽至真鹽條件、水深20~50 m的海域,僅少數有時可出現在沿岸水域中。淺海多鹽-真鹽型介形類在黃茅海河口灣中的豐度僅次于近岸廣鹽型,共發現10種,占介形類殼體總數的39.41%。主要屬種包括Bicorncythere bisanensis、Alocopocythere kendengensis、Sinocythere sinensis、Neosinocythere elongata、Pistocythereis bradyformis、Keijella hodgii和Loxoconcha ocellata。其他豐度較低的屬種有Munseyella pupilla、Hemicytherura cuneata和Hemicytherura biannularis等。
(3)廣海真鹽型:該介形類類型主要是在正常鹽度的大陸架開闊海域,在河口地區其豐度和分異度通常都很低。廣海真鹽型的介形類在黃茅海河口灣發現4種,殼體僅占總數的3.22%。其中Stigmatocythere bona較為常見,其他屬種僅零星出現,包括Stigmatocythere rosemani、Cytheropteron miurense和Paracytherois angusta等。
(4)淡水-微鹽型:也可稱為陸相型,多生活于淡水環境,偶在微鹽水條件也可出現。該類型在黃茅海河口灣地區僅發現Candona屬的1種分子,豐度低,其殼體僅占總數的2.45%。
3 討論
珠江口表層沉積物中微體生物的空間分布可受水體鹽度、水動力條件和水深的控制[11,13]。由于受潮汐作用和徑流作用的雙重影響,不同位置的水體條件變化大,導致介形類豐度、生態類型組成變化大(圖2和圖3)。
(1)Ⅰ區:大多數樣品中不含任何微體生物,僅少數樣品見零星近岸廣鹽型介形類殼體,豐度小于50枚/100 g,且僅出現Sinocytheridea impressa和Neomonoceratina delicata 2個屬種。由于該區域水動力很強[11],生物難以生存和保存,包括介形類。此外,受徑流匯入的影響,該區域內底層水體鹽度非常低(洪季底層水鹽度小于1‰),也不利于介形類生存。
(2)Ⅱ區:介形類豐度一般在150枚/100 g以下,且殼體多為淺海多鹽-真鹽型小個體。受徑流下泄作用外圍的影響,該區域水動力仍然較強,且鹽度較低(洪季底層鹽度小于3‰),介形類仍較難生存。
(3)Ⅲ區:介形類豐度變化大(250~1 500枚/100 g),北部一般低于500枚/100 g,東側受大杧島影響,水動力較弱,豐度增加可達1 000枚/100 g以上。枯季受海洋影響顯著,洪季仍受下泄淡水影響,鹽度變化較大(3‰~25‰),適應力強的類型才能生存,區域內以淺海多鹽-真鹽型分子占主導,多數站位占50%,有的甚至達100%。淺海多鹽-真鹽型分子中Keijella hodgii比較豐富,有的樣品中可達33%,該種雖屬分布范圍較廣的淺海多鹽-真鹽型分子,但多分布于較淺水域。其次優勢類型為近岸廣鹽型,總體占45%左右。此外還偶見海相分子。
(4)Ⅳ區:該區域鹽度全年大于15‰,水動力條件中等,因此該區域是介形類最為豐富的區域,介形類豐度多為1 000~5 000枚/100 g,局部站位大于5 000枚/100 g。近岸廣鹽型多在50%以下;淺海多鹽-真鹽型分子減少,而廣海真鹽型分子比例增加。受黃茅海“東進西出”的影響,在黃茅海河口灣西口海域鹽度整體較東口低,且由于離岸較近,近岸廣鹽型占優勢。
(5)Ⅴ區:該區域內介形類豐度僅為中等,生態類型仍以近岸廣鹽型和淺海多鹽-真鹽型為主,但近岸廣鹽型相較其他水域略低,廣海真鹽型分子總體占比將近10%,部分站位可達34%。主要是因此處為黃茅海海灣東口,潮水常年從此處灌入黃茅海河口灣內,水動力條件較強,致使介形類豐度不高。同時,因該區域外側為開闊海域,水體底層鹽度常年大于28‰,廣海真鹽型分子得以發育。吳潔等[13]利用去趨勢對應分析,認為鹽度和沉積物含砂量(可代表水動力條件)是影響黃茅海地區介形類分布的主因。通過本次對介形類生態環境分布特征和分布環境的探討,印證黃茅海河口灣介形類分布受海水鹽度控制最大,其次為水動力條件。在水體鹽度小、水動力條件強的河流入海口及其影響區域,介形類豐度小,且生態類型單一;而隨著鹽度由北往南升高,水動力條件逐漸適宜,介形類的豐度逐漸增加,生態類型更加豐富,在黃茅海河口灣南部海域,由于海水底層鹽度全年在15‰以上,水動力條件適宜,介形類豐度和生態類型均達到最多。
4 結語
受黃茅海河口灣多變環境的影響,該地區底質沉積樣品中出現的16屬20種介形類可分為4種生態類型,總體表現為以近岸廣鹽型分子占主導地位,優勢度極高;淺海多鹽-真鹽型分子次之;而廣海真鹽型和淡水-微鹽型分子豐度很低。根據黃茅海口灣的水體條件可大致將研究區劃分為5種亞環境區,各區內的介形類生態類型組成也存在較大差別,表現出不同的空間分布特征。研究顯示介形類分布受海水鹽度控制最大,其次為水動力條件。在水體鹽度小、水動力條件強的河流入海口及其影響區域,介形類豐度小,且生態類型單一;而在黃茅海河口灣南部海域,由于海水底層鹽度全年在15‰以上,水動力條件適宜,介形類豐度總體較高,生態類型增多。
參考文獻
[1]WILLIAMS M A J,DUNKERLEY D L,DE DECKE P,et al.Quaternary environments[M].London:Arnold,1993.
[2]李淑鸞.珠江口底質中介形蟲埋葬群的分布[J].熱帶海洋,1985,4(4):43-52.
[3]汪品先,閔秋寶,卞云華,等.河口有孔蟲的搬運及其古環境意義[J].海洋地質與第四紀地質,1986,6(2):53-65.
[4]趙泉鴻,汪品先,張清蘭.南海北部陸架底質中介形蟲的分布[J].海洋學報,1986,82(5):590-602.
[5]趙泉鴻,汪品先.中國淺海現代介形蟲的數量和屬種分布[J].海洋與湖沼,1988,19(6):553-561.
[6]陳木宏.珠江崖門口沉積物中有孔蟲分布的定量分析[A].中國科學院南海海洋研究所.南海海洋科學集刊第9集[M].北京:科學出版社,1989:29-37.
[7]羅憲林,李春初,牟崇鍵.伶仃洋有孔蟲生物群和埋藏群的比較及河口沉積作用分析[J].海洋學報,2001,23(4):69-74.
[8]劉春蓮,FRSICH F T,董藝辛,等.珠江三角洲PRD 05孔的高分辨率介形類記錄與晚第四紀古環境重建[J].古地理學報,2008,10(3):313-322.
[9]李濤,向榮,李團結.珠江口表層沉積物底棲有孔蟲分布及環境指示[J].海洋地質與第四紀地質,2011,31(6):91-98.
[10]吳潔,劉春蓮,張素青,等.珠江三角洲東部晚第四紀微體動物群的古環境意義[J].中山大學學報(自然科學版),2014,53(6):45-52.
[11]WU J,LIU C,FRSICH F T,et al.Foraminifera as environmental indicators and quantitative salinity reconstructions in the Pearl River Estuary,southern China[J].The Journal of Foraminiferal Research,2015,45(3):205-219.
[12]吳月琴,劉春蓮,楊小強,等.珠江三角洲中部全新世以來的微體動物群記錄與古環境重建[J].海洋地質與第四紀地質,2019,39(2):31-43.
[13]吳潔,劉春蓮,賈良文,等.珠江口黃茅海河口灣表層沉積物中有孔蟲和介形蟲的分布及其影響因素[J].古地理學報,2013,15(3):413-422.
[14]黃方,葉春池,溫學良,等.黃茅海鹽度特征及其鹽水楔活動范圍[J].海洋通報,1994,13(2):33-39.
[15]吳超羽.黃茅海河口小尺度動力結構及其沉積作用[J].中山大學學報(自然科學版),1995,34(2):86-94.
[16]李平日,喬彭年,鄭洪漢,等.珠江三角洲一萬年來環境演變[M].北京:海洋出版社,1991.
[17]應強,曹民雄,孔祥柏.黃茅海海域內泥沙淤積范圍的確定[J].水科學進展,1997,8(1):48-53.
[18]賈良文,羅軍,任杰.珠江口黃茅海攔門沙演變及成因分析[J].海洋學報,2012,34(5):120-127.
[19]呂頌輝,齊雨藻,錢宏林,等.南海港灣浮游植物與赤潮生物研究Ⅱ:廣海灣[J].海洋通報,1993,12(2):57-62.
[20]張心鳳,詹杰民.黃茅海波-流共同作用下的三維懸沙數值模擬[J].計算力學學報,2009,26(6):840-845.
[21]張心鳳.黃茅海水域河床長期演變趨勢預測[J].武漢大學學報(工學版),2014,47(5):591-598.
[22]吳創收,劉歡,武亞菊,等.黃茅海河口沿程異常潮差:I-理論模型研究[J].海洋科學進展,2010,25(4):436-444.
[23]楊雪舞,于紅兵,孫宗勛,等.黃茅海河口灣現代動力地貌體系和沖淤過程分析[J].熱帶海洋,1997,16(1):49-59.
[24]DONG L,SU J,WONG L A,et al.Seasonal variation and dynamics of the Pearl River Plume[J].Continental Shelf Research,2004,24:1761-1777.
[25]LIU C,HUANG Y,WU J,et al.Miocene-Pliocene planktonic foraminiferal biostratigraphy of the Pearl River-Mouth Basin,northern South China Sea[J].Journal of Palaeogeography,2012,1:43-56.
