松江鱸IL 15 Rα的結(jié)構(gòu)特征與表達(dá)分析
2020-10-05 04:58:10林日錦隗涵涵鐘金妙劉瑩瑩
海洋漁業(yè) 2020年4期
關(guān)鍵詞:分析
林日錦,隗涵涵,鐘金妙,劉瑩瑩
(山東大學(xué)(威海)海洋學(xué)院,山東威海 264209)
白細(xì)胞介素15(IL 15)是4α螺旋細(xì)胞因子家族中的主要成員之一[1],通過與膜受體復(fù)合物IL 15R的相互作用,在適應(yīng)性免疫系統(tǒng)中發(fā)揮重要生物功能。IL 15R包括3個(gè)亞基:IL 15Rα、IL 15 Rβ及IL 15 Rγc。β鏈及γc鏈為IL 15與白細(xì)胞介素2(IL 2)的共享亞基,也是兩者在結(jié)構(gòu)與功能特征上重疊的基礎(chǔ)[2-4];α鏈具有特異性,介導(dǎo)不同的信號通路,使IL 2與IL 15產(chǎn)生不同生理效應(yīng)。IL 15Rα的mRNA存在于各種免疫或非免疫細(xì)胞和組織[5]。轉(zhuǎn)錄水平的IL 15Rα存在多種可變剪接形式,并能引發(fā)不同的信號通路[6]。蛋白水平上的IL 15R除以膜蛋白形式分布在細(xì)胞膜表面外,還能自發(fā)地或經(jīng)誘導(dǎo)后從膜表面脫落形成可溶性受體,并通過拮抗作用抑制IL 15與膜受體的結(jié)合[7]。
IL 15Rα因其與IL 15的高親和力及特異結(jié)合能力而最初分離于人體[8-9]。人的IL 15Rα由7個(gè)外顯子組成,編碼263個(gè)aa的蛋白多肽[9],且其胞外區(qū)段具有特殊的sushi結(jié)構(gòu)域,在與IL 15的結(jié)合中發(fā)揮關(guān)鍵作用[10]。在哺乳動(dòng)物中IL 15Rα結(jié)構(gòu)與功能相關(guān)研究較多,在非哺乳動(dòng)物中,雞(Gallusgallus)的IL 15Rα首次被發(fā)現(xiàn),其IL 15Rα亞基類似于哺乳動(dòng)物,參與IL 15信號介導(dǎo)[11]。在2006年之前,還未見有關(guān)魚類IL 15Rα的研究報(bào)道。紅鰭東方鲀(Takifugu rubripes)IL 15Rα同源基因的克隆鑒定是魚類中的首例[12];從虹鱒(Oncorhynchusmykiss)中也克隆到了魚類的IL 15Rα基因,并發(fā)現(xiàn)3種選擇剪接方式,其后也從其他硬骨魚中鑒定或預(yù)測到了IL 15Rα[13]。
本次研究材料為松江鱸(Trachidermus fasciatusHeckel),松江鱸隸屬于鲉形目(Scorpaeniformes),杜父魚科(Cottidae),松江鱸屬,是一種近海溯河洄游的肉食性魚類,已被列為國家Ⅱ級保護(hù)動(dòng)物[14-15]。目前,有關(guān)松江鱸IL 15Rα基因的研究尚未見報(bào)道。本實(shí)驗(yàn)克隆得到松江鱸IL 15Rα基因(命名為TfIL 15Rα),對其序列結(jié)構(gòu)特征進(jìn)行了分析,利用qRT PCR技術(shù)研究了IL 15Rα的mRNA在魚體內(nèi)的分布和免疫刺激后基因應(yīng)答情況,并對其與IL 15免疫刺激后的表達(dá)情況進(jìn)行了系統(tǒng)分析比較。
1 材料與方法
1.1 材料與組織樣品制備
實(shí)驗(yàn)動(dòng)物為取自山東省文登埠口松江鱸自然保護(hù)區(qū)9~10月齡的松江鱸(體質(zhì)量約15~23 g)。實(shí)驗(yàn)前經(jīng)充氣海水(12~14℃)飼養(yǎng)一周后進(jìn)行正常組織的RNA提取:先挑選健康個(gè)體進(jìn)行麻醉,心臟取血后解剖取其心臟、肝臟、鰓、腸、皮膚、腎臟、脾臟、腦、卵等組織,立即放入Trizol中研磨。
進(jìn)行脂多糖(lipopolysaccharides,LPS)刺激實(shí)驗(yàn)時(shí),將100條生長狀況相同的松江鱸平均分成兩組,實(shí)驗(yàn)組腹腔注50μL(0.04 mg·kg-1)的LPS,對照組注射同體積無菌生理鹽水。經(jīng)LPS刺激后分別于0、2、6、12、24、48、72、96 h麻醉取樣(每次6條),各從血、皮膚、肝臟和脾臟中提取總RNA。
1.2 總RNA的提取和cDNA合成
從各組織樣品中提取總mRNA是以Trizol(Invitrogen公司)的實(shí)驗(yàn)說明為依據(jù)進(jìn)行操作,并參照CLONTECH公司SMART的指導(dǎo)說明書進(jìn)行反轉(zhuǎn)錄合成cDNA第一鏈,引物見表1。
1.3 TfIL 15RαcDNA的全長克隆
從構(gòu)建的cDNA文庫中獲得TfIL 15Rα基因的EST序列,利用primer設(shè)計(jì)特異性引物,分別進(jìn)行3′ 和5′ RACE(表1)。PCR反應(yīng)條件為94℃預(yù)變性5 min,94℃變性30 s,55℃復(fù)性30 s,72℃延伸40 s,進(jìn)行30個(gè)循環(huán)后,72℃延伸7 min。然后將PCR純化產(chǎn)物與pMD 18T載體進(jìn)行連接、克隆測序,獲得5′端和3′端序列片段。
1.4 TfIL 15Rα基因的序列分析
TfIL 15RαcDNA的全長通過軟件BioEdit對EST序列及5′ 和3′ 端序列片段進(jìn)行拼接獲得;基因的蛋白翻譯、等電點(diǎn)及分子量預(yù)測通過在線軟件Expasy(http://www.au.expasy.org/)進(jìn)行;使用http://www.cbs.dtu.dk/services/NetNGlyc/和PROSITE服務(wù)器(http://www.Expasy.org/prosite)對蛋白N 糖基化位點(diǎn)及可能的基序(motif)進(jìn)行預(yù)測。通過SMART(http://smart.embl heidelberg.de/)對蛋白的信號肽、跨膜區(qū)段和sushi結(jié)構(gòu)域進(jìn)行預(yù)測;同源序列從NCBI下載后由Clustal W進(jìn)行比對分析;并利用MEGA7.0軟件進(jìn)行TfIL 15Rα序列的系統(tǒng)發(fā)育分析。

表1 PCR引物名稱及序列Tab.1 PCR primers and sequences
1.5 組織表達(dá)分析
應(yīng)用qRT PCR來分析TfIL 15的組成型表達(dá)及刺激后表達(dá)模式的變化。設(shè)計(jì)特異性qRT PCR引物,模板為正常組織及不同時(shí)間點(diǎn)試驗(yàn)組和對照組的cDNA,內(nèi)參基因選擇β actin,按此進(jìn)行反應(yīng)。反應(yīng)和分析使用7300實(shí)時(shí)熒光定量PCR儀(Applied Biosystems ABI,美國)進(jìn)行。反應(yīng)條件為:94℃3 min;94℃15 s,60℃60 s,40個(gè)循環(huán)[18]。反應(yīng)結(jié)束后進(jìn)行熔解曲線分析。每個(gè)樣品cDNA進(jìn)行3個(gè)平行實(shí)驗(yàn)。基因的相對表達(dá)量根據(jù)2 ΔΔCt法計(jì)算,使用T檢驗(yàn)的方法進(jìn)行差異顯著性分析,P<0.05時(shí)認(rèn)為是可接受的顯著性差異標(biāo)準(zhǔn)。
2 結(jié)果與分析
2.1 TfIL 15Rα基因克隆及序列分析
松江鱸TfIL 15Rα的cDNA全長為998 bp(GenBank登錄號:MT293626),包括5′ 非編碼區(qū)(5′ UTR)85 bp,開放閱讀框(ORF)663 bp和3′UTR 250 bp(圖1)。基因ORF編碼氨基酸220aa,其中前27aa為信號肽序列,成熟肽全長193aa。松 江 鱸TfIL 15Rα為 膜 受 體,經(jīng)TMHMM2.0軟件預(yù)測TfIL 15Rα蛋白包含145aa的胞外區(qū),23aa的跨膜區(qū)(173~195 aa)及25aa的胞內(nèi)區(qū),且其胞外區(qū)只有一個(gè)sushi結(jié)構(gòu)域(31~95 aa)。成熟肽含有1個(gè)可能的N糖基化位點(diǎn):NDTQ,40~43;4個(gè)casein kinase II phosphorylation位點(diǎn)(5~8 aa,159~162 aa,199~202 aa,207~210 aa);兩個(gè)N myristoylation位點(diǎn)(21~22 aa,117~122 aa);1個(gè)Protein kinase C phosphorylation位點(diǎn)(55~57 aa);各有1個(gè)c AMP and cGMP dependent protein kinase phosphorylation位點(diǎn)(100~103 aa及196~199 aa),這提示松江鱸IL 15Rα可經(jīng)過不同修飾而產(chǎn)生結(jié)構(gòu)的變化。
將TfIL 15Rα與其他物種同源序列進(jìn)行比對(表2),發(fā)現(xiàn)TfIL 15Rα與三刺魚(Gasterosteus aculeatus)的同源性最高為46%,其次為牙鲆(Paralichthysolivaceus)40%,與斑馬魚(Danio rerio)同源性為19%,與魚類序列同源性為19%~46%,與哺乳類的同源性為23%~34%,相似性較低。多序列比對發(fā)現(xiàn),同源區(qū)主要集中在sushi結(jié)構(gòu)域部分,該區(qū)域所有物種的4個(gè)半胱氨酸(Cys)高度保守。此外,魚類還有另外兩個(gè)保守的Cys(圖2)。

圖1 TfIL 15Rα的cDNA序列及其編碼序列Fig.1 Nucleotide and deduced am ino acid sequences of TfIL 15Rα

表2 脊椎動(dòng)物IL 15同源性比對分析Tab.2 Am ino acid identity of TfIL 15Rαw ith other vertebrate IL 15 Rα
以NJ法構(gòu)建系統(tǒng)發(fā)育樹(圖3),其結(jié)果顯示松江鱸IL 15Rα與三刺魚同源性較高,總體上與其他魚類共聚為一支,而鳥類與哺乳動(dòng)物聚為另一大支。
2.2 TfIL 15Rα組織表達(dá)模式分析
qRT PCR結(jié)果表明,TfIL 15 mRNA于松江鱸血、心、肝、鰓、腸、腎和卵等組織器官中均廣泛表達(dá)。其中,鰓中的表達(dá)量最高,其次為腸和卵(圖4)。
2.3 LPS刺激后各組織中TfIL 15基因的表達(dá)變化
腹腔注射LPS后,血液、皮膚、肝臟和脾臟中的TfIL 15RαmRNA表達(dá)量均上調(diào)(圖5)。在皮膚中,TfIL 15RαmRNA的表達(dá)量于刺激后2 h明顯上調(diào),達(dá)到對照組的14倍,其后顯著下調(diào),并一直維持在較低表達(dá)水平;在血液中,TfIL 15Rα mRNA表達(dá)量同樣于刺激后2 h迅速達(dá)到最高峰,為對照組的52倍,刺激后6 h與刺激后2 h相比,TfIL 15RαmRNA表達(dá)水平呈現(xiàn)較為顯著的下降;在肝中,刺激后2 h,TfIL 15RαmRNA表達(dá)量上調(diào)至對照組的21倍,其后下降,并維持在較低水平,直到96 h再次上調(diào)表達(dá),為對照組的25倍;在脾臟中,刺激后2 h TfIL 15RαmRNA表達(dá)量小幅度上調(diào),至12 h達(dá)到最大表達(dá)量,為對照組的2倍,此后又下調(diào)表達(dá),但變化幅度較小。

圖3 基于不同物種IL 15 Rα的氨基酸序列構(gòu)建的系統(tǒng)發(fā)育樹Fig.3 Phylogenetic tree constructed w ith am ino acid sequences of IL 15 Rαfrom various species
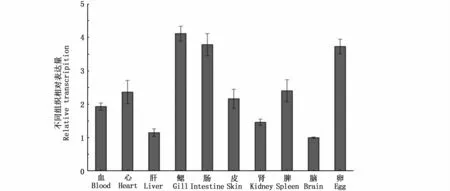
圖4 TfIL 15Rα基因在正常組織中的表達(dá)模式Fig.4 Constitutive expression of TfIL 15RαmRNA in different tissues

圖5 LPS刺激后TfIL 15Rα在不同組織的表達(dá)模式變化Fig.5 Temporary expression of TfIL 15RαmRNA in different tissues post LPS challenge
3 討論
本研究成功克隆了松江鱸IL 15Rα基因的cDNA全長,并命名為TfIL 15Rα。TfIL 15Rα編碼220aa的多肽,其中信號肽長27aa,成熟肽全長193aa。與松江鱸IL 15Rα不同,哺乳類的IL 15 Rα成 熟 肽 長 度 為231~237aa[8],雞 的 為203aa[9]。成熟肽包括145aa的胞外區(qū)段(28~147),23aa的跨膜區(qū)段(173~195)和25aa的胞內(nèi)區(qū)段(196~220),說明TfIL 15Rα編碼的蛋白與哺乳動(dòng)物IL 15Rα類似,是一種膜型受體蛋白[7]。在胞外區(qū)段存在一個(gè)特殊的sushi結(jié)構(gòu)域(31~95),與IL 15具有高親和力,在與細(xì)胞因子的結(jié)合中發(fā)揮重要作用。
經(jīng)氨基酸序列比對發(fā)現(xiàn),不同物種的同源相似區(qū)主要集中在sushi結(jié)構(gòu)域,這與其承擔(dān)著最重要的配體結(jié)合能力相關(guān)[16]。sushui結(jié)構(gòu)域內(nèi)的4個(gè)半胱氨酸具有高度保守性,在哺乳動(dòng)物及魚類中都有存在,并形成兩對結(jié)構(gòu)域內(nèi)二硫鍵,使sushui區(qū)結(jié)構(gòu)相似,這是其發(fā)揮配體結(jié)合功能的重要位點(diǎn)。另外,在松江鱸、暗綠鲀、紅鰭東方鲀等魚類的IL 15Rα中,還存在兩個(gè)空間位置鄰近的半胱氨酸,可能還能夠參與形成魚類特異的二硫鍵,從而使結(jié)構(gòu)域穩(wěn)定性得以增強(qiáng)[17]。總體上看,TfIL 15Rα與三刺魚的同源性最高,但也只有46%。與魚類序列同源性為19%~46%,變異程度比較大,這也解釋了目前魚類IL 15R研究較少的原因。
qRT PCR結(jié)果顯示,TfIL 15Rα廣泛表達(dá)于松江鱸各個(gè)組織器官中,包括血、心、肝、鰓、腸、皮、腎、腦、卵等,與哺乳動(dòng)物及紅鰭東方鲀、虹鱒等魚類中IL 15Rα的表達(dá)具有相似性。TfIL 15Rα的廣泛性表達(dá)模式類似實(shí)驗(yàn)前期研究的松江鱸TfIL 15的表達(dá)[21],表明IL 15Rα可能在機(jī)體正常生理過程中發(fā)揮重要的作用。值得注意的是,在卵中,TfIL 15Rα也表現(xiàn)出了與TfIL 15相似的較高的轉(zhuǎn)錄水平表達(dá)。IL 15/IL 15Rα表達(dá)模式的相似性,與IL 15/IL 15Rα信號轉(zhuǎn)導(dǎo)系統(tǒng)能在這些場所發(fā)揮作用是相關(guān)的。
在分析TfIL 15Rα正常組織分布情況的基礎(chǔ)上,我們以LPS為刺激物進(jìn)行刺激實(shí)驗(yàn),進(jìn)一步分析了TfIL 15Rα在機(jī)體免疫中的作用。結(jié)果顯示,在血液、皮膚和肝臟中,LPS刺激后2 h,TfIL 15Rα迅速上調(diào)至最高峰,在脾臟中12 h達(dá)到表達(dá)高峰,比血液、皮膚和肝臟稍有延遲。血液系統(tǒng)作為全身循環(huán)系統(tǒng),承載各種免疫細(xì)胞和細(xì)胞因子;皮膚作為機(jī)體首先接觸外界復(fù)雜水體環(huán)境的免疫組織,是最直接和快速的LPS應(yīng)答和反應(yīng)器官;而肝臟是魚類多種重要免疫相關(guān)蛋白的合成場所,故經(jīng)LPS刺激后短時(shí)間內(nèi)能檢測到TfIL 15Rα的上調(diào)表達(dá)。值得注意的是,在肝臟中,96 h再次出現(xiàn)一個(gè)新的表達(dá)高峰,這與松江鱸IL 15在肝中的表達(dá)情況相似[18],這是因?yàn)門fIL 15Rα作為TfIL 15特異受體,參與免疫相關(guān)信號通路激活與傳導(dǎo),故兩者表達(dá)變化模式類似。96 h再次達(dá)到高峰推測是因?yàn)長PS刺激后魚類先天免疫應(yīng)答系統(tǒng)快速做出反應(yīng),激活一群免疫細(xì)胞,激活的免疫細(xì)胞分泌細(xì)胞因子,再次激活另一群免疫細(xì)胞,進(jìn)行二次免疫應(yīng)答。脾臟是各種淋巴細(xì)胞和白細(xì)胞等免疫細(xì)胞產(chǎn)生、儲(chǔ)存和成熟的主要場所,因而上調(diào)時(shí)間有所滯后。小鼠感染肺炎后肺泡巨噬細(xì)胞和氣道上皮細(xì)胞也同時(shí)上調(diào)表達(dá)IL 15和IL 15Rα[21]。IL 15的產(chǎn)生細(xì)胞,尤其是單核細(xì)胞和樹突狀細(xì)胞,可同時(shí)表達(dá)IL 15和IL 15Rα[22]。因此,松江鱸IL 15和IL 15Rα相似的上調(diào)表達(dá)模式提示,這兩個(gè)基因可能通過其結(jié)合,從而在抗微生物感染的免疫反應(yīng)中發(fā)揮作用。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會(huì)科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
