smad1/5信號通路在誘導多能干細胞成釉分化中的作用研究
2020-10-26 02:05:22覃文聘張樂琪
醫學研究雜志 2020年3期
關鍵詞:實驗
劉 治 覃文聘 高 鵬 譚 蕾 張樂琪
誘導多能干細胞(induced pluripotent stem,iPS)被認為是實現牙齒再生的一種新的干細胞來源,其來源于患者自體成熟細胞,因此成功克服了細胞來源受限、倫理和免疫排斥上的障礙,具有良好的臨床應用前景。研究顯示,iPS細胞可被誘導分化為神經細胞、胰島素分泌細胞、心血管和造血細胞、肝臟、腎臟等細胞[1]。Liu等[2]近期研究表明,iPS細胞可在成釉細胞無血清條件培養基(ameloblast serum-free conditioned medium,ASF-CM)誘導下,分化為成釉細胞樣細胞,并表達成釉細胞標志物成釉蛋白(ameloblastin,AMBN)及細胞角蛋白-14(cytokeratin-14,CK14);當在ASF-CM中添加骨形態發生蛋白4 (morphogenetic protein 4,BMP4)可顯著促進iPS細胞的成釉分化,而加入了BMP抑制劑noggin則可顯著抑制iPS細胞的成釉分化,表明BMP家族分子在iPS細胞成釉分化過程中發揮關鍵調控作用。然而,iPS細胞成釉分化過程中的信號轉導通路目前仍不清楚。
骨形態發生蛋白(bone morphogenetic proteins,BMPs)屬于轉化生長因子β(transforming growth factor β,TGF-β)家族成員,是一組能廣泛參與調節多種細胞的增生、遷移、分化和凋亡的生物學過程的功能蛋白,其在早期胚胎發生和隨后的器官形成中發揮關鍵調控作用。BMP信號通過激活其受體磷酸化smad1、5、8蛋白,從而啟動細胞內信號轉導,繼而直接或間接通過 DNA 結合蛋白或重新合成蛋白質來調節靶基因的轉錄。此外,有研究顯示BMPs也可直接激活絲裂原活化蛋白激酶(mitogen-activated protein kinases,MAPKs)通路,其中,ERK1/2信號轉導通路主要調控細胞生長與分化,JNK和 p38 MAPK 信號轉導通路參與炎癥、細胞凋亡等應激反應[3]。然而,smad與MAPKs信號通路在iPS細胞被誘導向成釉細胞分化過程中的調控作用,目前仍未見文獻報道。
本研究采用ASF-CM誘導iPS細胞成釉分化,檢測分化過程中smad1/5與MAPKs信號分子磷酸化水平,并觀察smad1/5或MAPKs通路抑制劑對iPS成釉分化的影響,從而探明smad1/5與MAPKs信號通路在iPS細胞成釉分化中的作用。
材料與方法
1.實驗材料:小鼠C5系iPS細胞購自中國科學院廣州生物醫藥與健康研究院。成品上皮細胞培養液購自美國Sciencell公司,smad1/5通路抑制劑LDN-193189購自美國Selleck公司,p38 MAPK通路抑制劑SB203580及ERK1/2通路抑制劑U0126,以及phospho-smad1/5、phospho-p38、total-p38、phospho-ERK1/2、total-ERK1/2、phospho-JNK、total-JNK及GAPDH抗體均購自美國Cell Signaling Technology公司。RIPA細胞裂解液及BCA蛋白定量試劑盒購自上海碧云天生物科技有限公司,RNeasy Mini kit試劑盒購自美國Invitrogen公司,反轉錄試劑盒(DRR037A)及SYBR法實時定量PCR試劑盒購自日本TaKaRa公司,Oct-4、SSEA-4、SSEA-1、AMBN、ENAM和CK14抗體均購自美國Santa Cruz 生物技術公司,MTT檢測試劑盒(C0009)購自上海碧云天生物技術公司。
2.實驗方法:分離培養小鼠胚胎成纖維細胞:將懷孕12.5~14.5天 C56孕鼠斷頸處死,無菌條件下取出胎鼠,小心分離其軀干并置于盛有PBS的平皿內,充分洗滌棄除紅細胞。用無菌眼科剪剪碎鼠胚軀干,加適量胰酶37℃消化30min后終止消化,常規離心棄上清,加適量含10%胎牛血清的DMEM培養液,反復吹打后接種置細胞培養瓶中,37℃、5%CO2、飽和濕度培養,培養6天后傳代。
3.制備iPS細胞滋養層:將絲裂霉素(10μg/ml)加入到上述長滿胚胎成纖維細胞的培養瓶中孵育3h,吸棄廢液PBS充分洗滌后,加入0.25%胰酶消化。在顯微鏡下觀察,當貼壁細胞層出現裂隙且有少量細胞漂浮時,用移液管反復吹打瓶底,加含血清培養液終止消化,常規離心后棄上清,以3.0×104/cm2密度鋪在經0.1%明膠溶液預處理的培養瓶中,于培養箱中靜置過夜,使細胞貼壁。鋪好的飼養層在7 天內可有效支持iPS細胞的生長并維持其全能性。
4.培養小鼠iPS細胞:于37℃水浴中迅速解凍iPS細胞,將細胞懸液吸至加有少量PBS的離心管中,常規離心棄上清,加入6ml含10%胎牛血清的DMEM培養液,吹打混勻為單細胞懸液,并移至已鋪好飼養層細胞的培養瓶中,37℃、5%CO2、飽和濕度培養,每天換液,觀察克隆的大小和密度,一般在培養后的第3 天可進行傳代。
5.成釉細胞無血清條件培養基(ASF-CM)的制備:斷頸處死出生7天的C57 仔鼠20只,無菌分離仔鼠下頜骨, PBS沖洗干凈后分離其下頜切牙牙胚,在體視學顯微鏡下用顯微鑷剝離成釉上皮,浸泡于含 10%胎牛血清的DMEM培養液中。剪碎組織后加入2ml Ⅰ型膠原酶,于37℃消化1h,終止消化后常規離心,棄上清,加入無血清的上皮細胞培養液,以1×105/ml密度接種于培養瓶內,37℃、5%CO2、飽和濕度培養。待細胞貼壁后,采用差別消化法純化上皮細胞,首先加入0.25%胰酶消化細胞5min,倒置顯微鏡下觀察到成纖維樣細胞完全脫壁,此時加入含血清培養基終止消化,PBS反復沖洗后加入無血清的上皮細胞培養液繼續培養。可重復此法,直至貼壁的完全是上皮細胞而沒有成纖維細胞。當成纖維樣細胞全部去除后,開始收集成釉細胞無血清條件培養液(ASF-CM),避光條件下用無血清上皮細胞培養液每天換液1次,置換出來的ASF-CM用小濾器過濾后-80℃冷藏備用。為確保實驗的可重復性,將所有ASF-CM混勻后分裝,用于下述實驗。
6.iPS細胞誘導分化及通路阻斷:以1∶1體積比配制ASF-CM與iPS常規培養液混合液(ASF-CM),分別在上述混合液中加入200nmol/L LDN-193189(smad1/5通路抑制劑),或10μmol/L SB203580 (p38 通路抑制劑) 或 10μmol/L U0126 (ERK1/2通路抑制劑),采用ASF-CM或加入上述通路抑制劑的ASF-CM培養iPS細胞,2天換液1次,以常規iPS培養液培養作為陰性對照,培養14天后收集細胞,進行下述蛋白及基因檢測。
7.Western blot法檢測蛋白表達量:RIPA裂解細胞30min,4℃高速離心取上清,BCA蛋白定量法測定樣品總蛋白濃度;取各組蛋白樣品40μg用體積分數為10%的十二烷基硫酸鈉-聚丙烯酰胺凝膠進行凝膠電泳后,再以半干法轉印到PVDF膜上,5.0%脫脂牛奶封閉 1h,然后分別加兔抗小鼠phospho-smad1/5、GAPDH、phospho-p38、total-p38、phospho-ERK1/2、total-ERK1/2、phospho-JNK及total-JNK一抗(1∶1000,美國Cell Signaling Technology公司),4℃孵育過夜;TBST洗膜后,滴加辣根過氧化物酶標記的羊抗兔二抗(1∶5000,北京中杉金橋生物技術公司)室溫孵育1h;TBST再次洗膜后,ECL化學發光試劑顯影,化學發光凝膠成像系統檢測蛋白條帶,并計算蛋白的相對表達水平。
8.實時定量PCR:RNeasy Mini kit試劑盒提取樣品總RNA,反轉錄試劑盒反轉錄合成cDNA,再以cDNA為模板,以GAPDH為內參照,采用SYBRⅡ試劑盒檢測各組細胞Ambn、ENAM及Ck14 mRNA的表達水平,反應條件嚴格按產品說明書進行;所用引物均由日本TaKaRa公司合成,具體引物序列見表1。最后以目的基因與 GAPDH 的起始拷貝數的比值表示目的基因的相對表達量。實驗重復3次。
9.免疫熒光染色:4%多聚甲醛固定細胞15min,PBS洗后滴加0.25% Triton X-100,37℃孵育15min,PBS洗后加入4%山羊血清孵育30min,不沖洗,甩干后直接加入相應一抗 (1∶100),4℃孵育過夜;PBS洗3次, 滴加FITC或Rhodamine標記的二抗(1∶200),37℃避光孵育30min;5μg/ml DAPI襯染細胞核2min,Olympus 熒光顯微鏡觀察及DP Manager軟件處理圖。

表1 PCR引物序列
10.MTT實驗:分別在上述阻斷實驗進行1、7、14天時,在細胞培養孔中加入MTT 溶液(5mg/ml)20μl,繼續孵育4h,每孔加入100μl Formazan溶解液并混勻,繼續孵育使結晶物充分融解,選擇 570nm 波長,在酶標儀上測定各孔吸光度值,記錄結果。
11.統計學方法:采用SPSS 11.0統計學軟件對數據進行方差分析,采取盲法獲取及計算以上數據,即由兩個對實驗設計不了解的實驗員各計數2次,并間隔1周。經標準一致性檢驗顯示,兩人所得數據高度吻合(r>0.9),因此兩人計數的平均值被用于統計分析。采用SNK-q檢驗進行兩兩亞組間比較,以P<0.05為差異有統計學意義。
結 果
1.Western blot法檢測:用于實驗的ips細胞表達胚胎干細胞特異性標記分子SSEA-4及Oct-4,陽性細胞百分比均高于95%,但不表達分化的胚胎干細胞標志物SSEA-1(圖1),表明其具有胚胎干細胞樣特性。本實驗采用Western blot法檢測磷酸化的smad1/5及MAPKs通路分子的表達。結果顯示,經ASF-CM處理的iPS細胞p-smad1/5 (6.1±1.3)、p-P38 (3.2±0.6)以及p-ERK1/2 (4.1±0.8)的表達較對照組顯著升高(圖2、圖3),但其p-JNK表達水平與對照組比較,差異無統計學意義(t=0.67,P=0.61)。
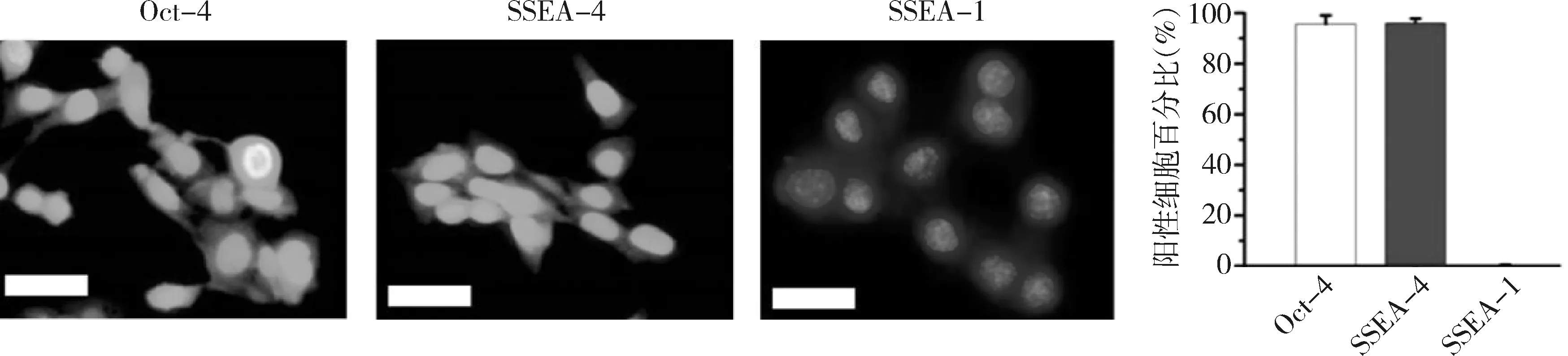
圖1 iPS細胞免疫熒光染色及陽性細胞定量(×40)Oct-4與SSEA-4為胚胎干細胞特異標記分子,SSEA-1為分化的胚胎干細胞標志物
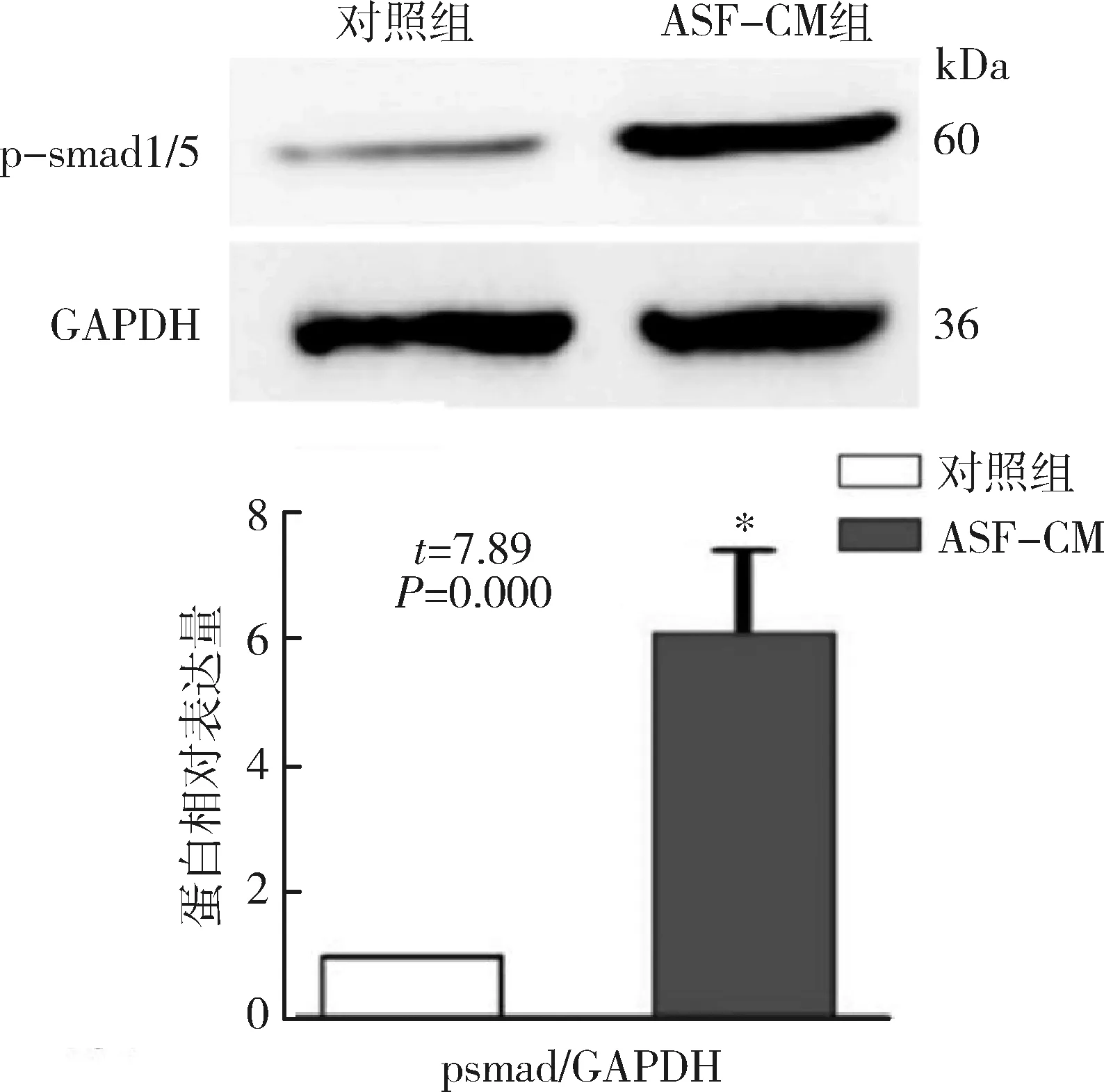
圖2 Western blot法檢測及定量對照培養基或ASF-CM培養iPS細胞后p-smad1/5蛋白表達水平與對照組比較,*P<0.01
2.實時定量PCR結果:分別將smad1/5、P38以及ERK1/2信號通路抑制劑加入到ASF-CM中培養iPS細胞14天,并以對照培養基或ASF-CM培養的iPS細胞作對照,檢測iPS細胞成釉細胞特異性標志物(Ambn、ENAM和Ck14)基因的表達。對照組、ASF-CM組、ASF-CM+LDN-193189組、ASF-CM+SB203580組及ASF-CM+U0126組iPS細胞Ambn(F=4.290,P<0.05)、ENAM(F=3.620,P<0.05)和Ck14(F=3.110,P<0.05)的表達比較,差異有統計學意義,其中ASF-CM組iPS細胞Ambn(6.6±1.3)、ENAM(5.4±0.9)和Ck14 mRNA(5.9±1.1)的表達較對照組顯著增高(P<0.05),smad1/5抑制劑LDN-193189可顯著反轉ASF-CM的上述促iPS細胞成釉分化的效應,表現為ASF-CM+LDN-193189組iPS細胞Ambn(2.4±0.5)、ENAM(1.9±0.6)和Ck14 mRNA(1.4±0.4)的表達較單純ASF-CM組顯著降低(P<0.05,圖4),但仍舊高于對照組(P<0.05)。然而,p38 MAPK通路抑制劑SB203580及ERK1/2通路抑制劑U0126則對ASF-CM促iPS細胞成釉分化的效應無顯著影響,表現為ASF-CM+SB203580組及ASF-CM+U0126組iPS細胞Ambn(SB203580組:6.1±1.1;U0126組:5.9±0.8)、ENAM(SB203580組:4.8±0.9;U0126組:5.1±0.5)和Ck14 mRNA(SB203580組:5.3±1.2;U0126組:4.9±0.6)的表達與單純ASF-CM組比較,差異無統計學意義(P>0.05,表2)。
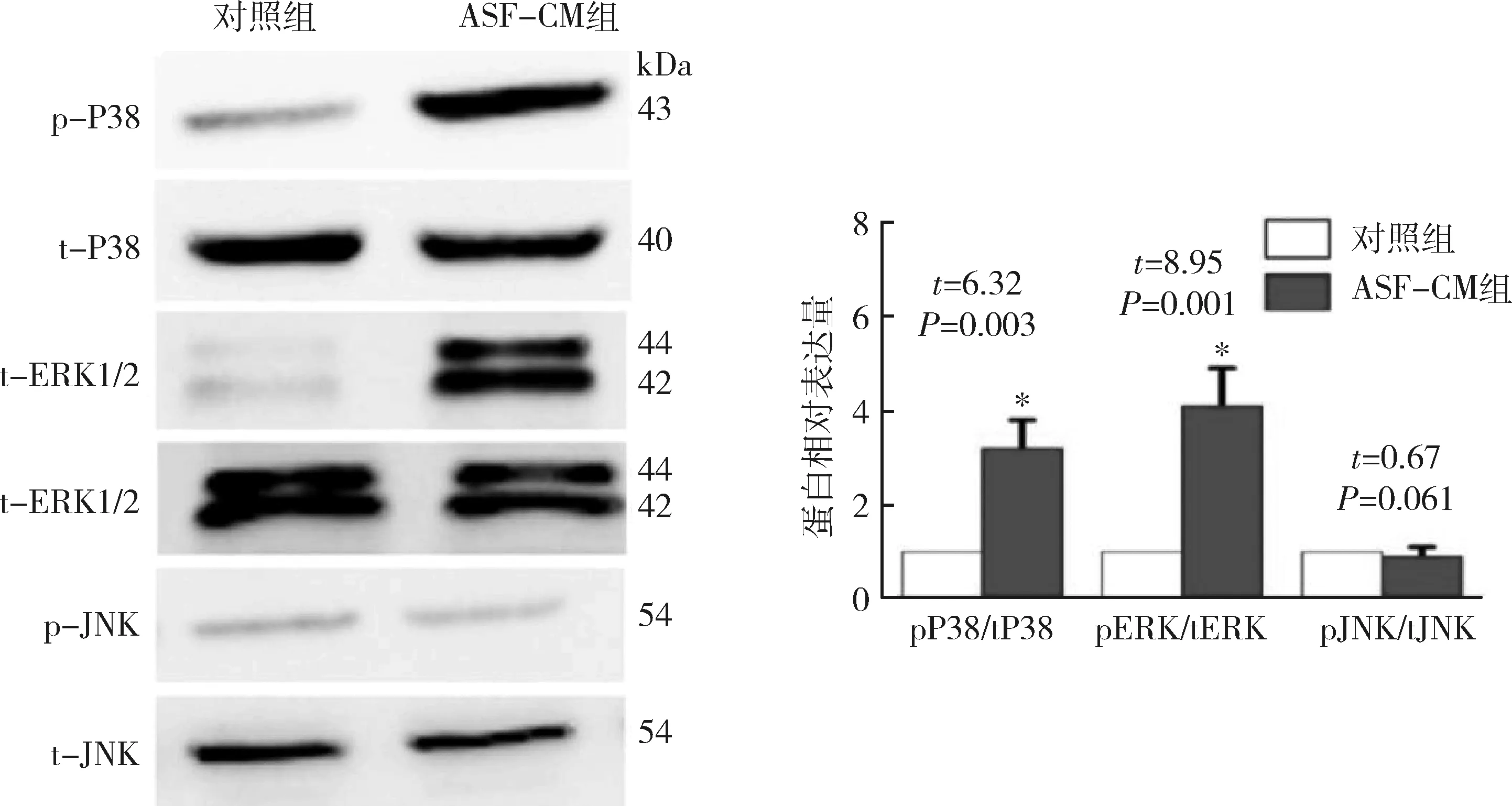
圖3 Western blot法檢測及定量對照培養基或ASF-CM培養iPS細胞后MAPKs通路分子蛋白表達水平與對照組比較,*P<0.01
3.免疫熒光染色結果:采用對照培養基培養的iPS細胞基本不表達成釉細胞特異性標志物Ambn、ENAM及Ck14,ASF-CM組及ASF-CM+抑制劑組iPS細胞均可表達Ambn、ENAM及Ck14蛋白,主要表達在胞核及胞質(圖4)。定量研究結果顯示對照組、ASF-CM組、ASF-CM+LDN-193189組、ASF-CM+SB203580組及ASF-CM+U0126組iPS細胞Ambn(F=44.190,P<0.05)、ENAM(F=52.160,P<0.05)和Ck14(F=40.680,P<0.05)陽性細胞百分比組間比較,差異有統計學意義,其中ASF-CM組iPS細胞Ambn(46.6±11.3)、ENAM(51.4±10.9)及Ck14(41.9±8.1)陽性細胞百分比均較對照組顯著增高(P<0.05),smad1/5抑制劑LDN-193189可顯著逆轉ASF-CM誘導iPS細胞成釉分化的作用,表現為ASF-CM+LDN-193189組iPS細胞Ambn(8.4±0.5)、ENAM(6.9±0.6)和Ck14 (4.4±0.4)陽性細胞百分比均較單純ASF-CM組顯著降低(P<0.05),但仍高于對照組(P<0.05)。ASF-CM+SB203580組及ASF-CM+U0126組iPS細胞Ambn(SB203580組:44.1±8.1;U0126組:45.9±8.8)、ENAM(SB203580組:49.8±10.9;U0126組:50.1±7.5)和Ck14(SB203580組:38.3±9.2;U0126組:40.9±10.6)陽性細胞百分數與單純ASF-CM組比較,差異無統計學意義(P>0.05,表3)。
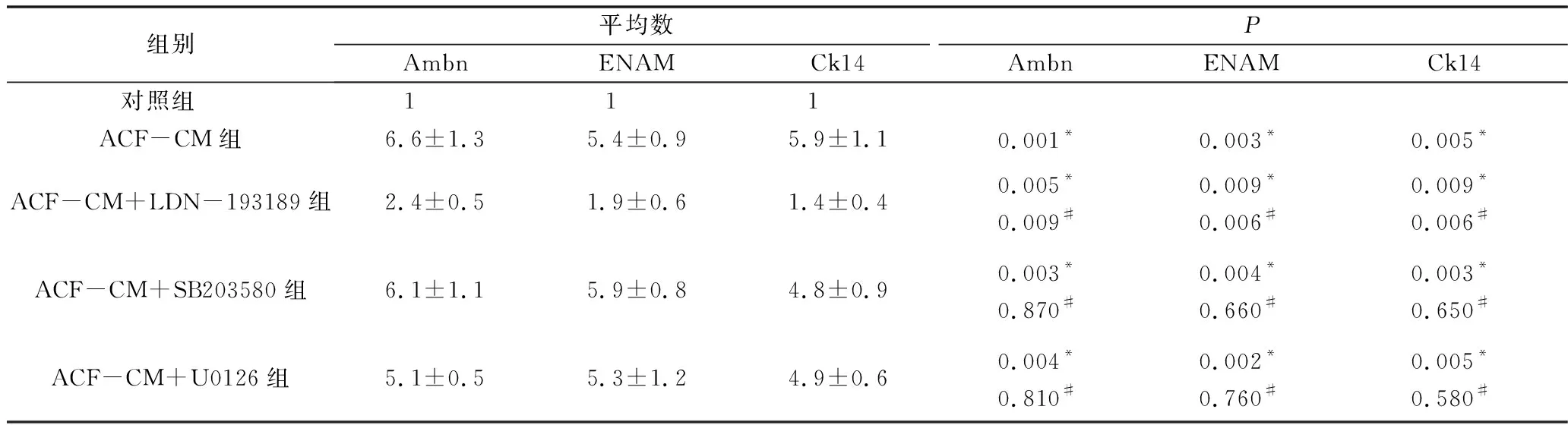
表2 實時定量PCR檢測
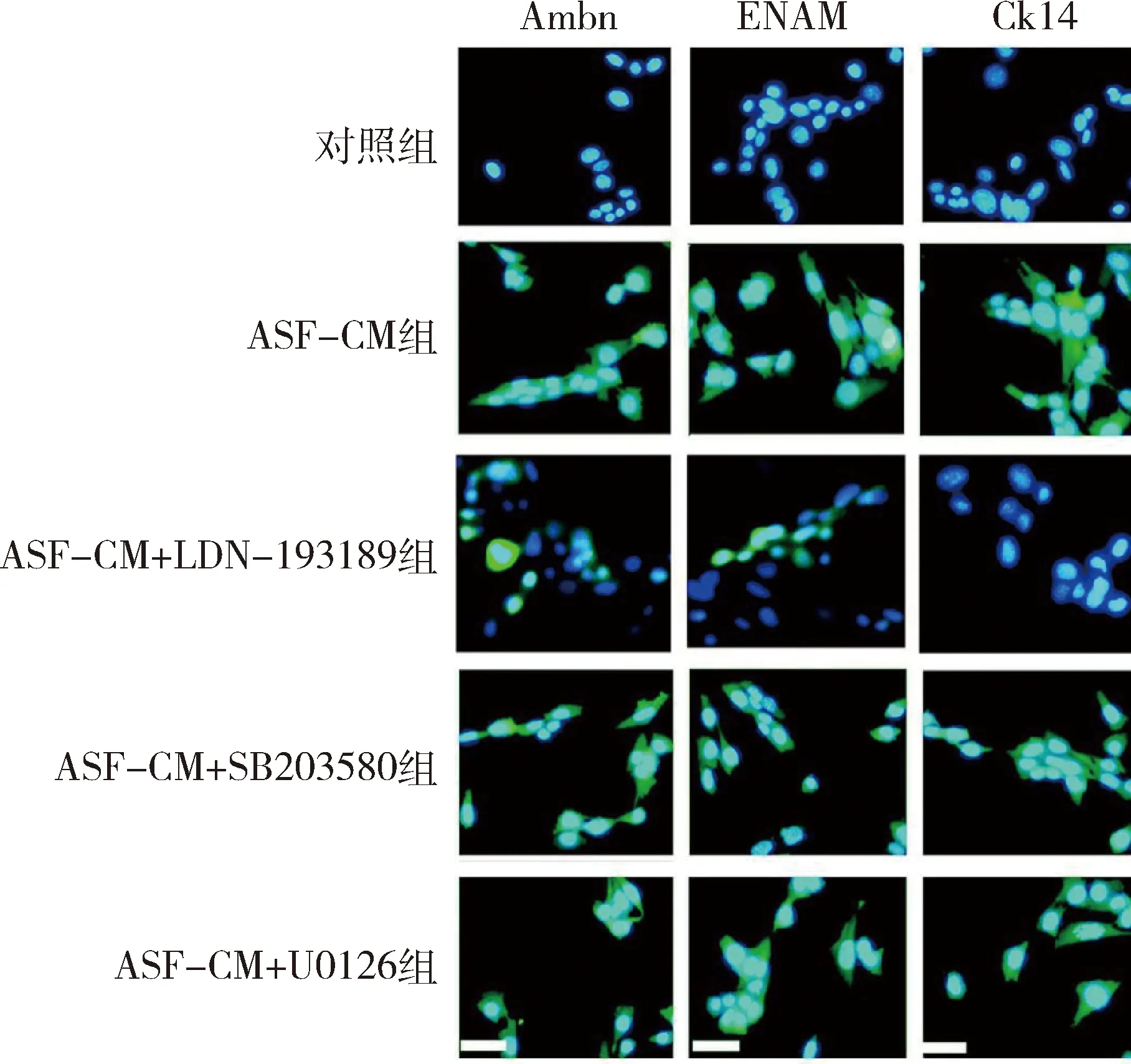
圖4 免疫熒光染色觀察(×40)
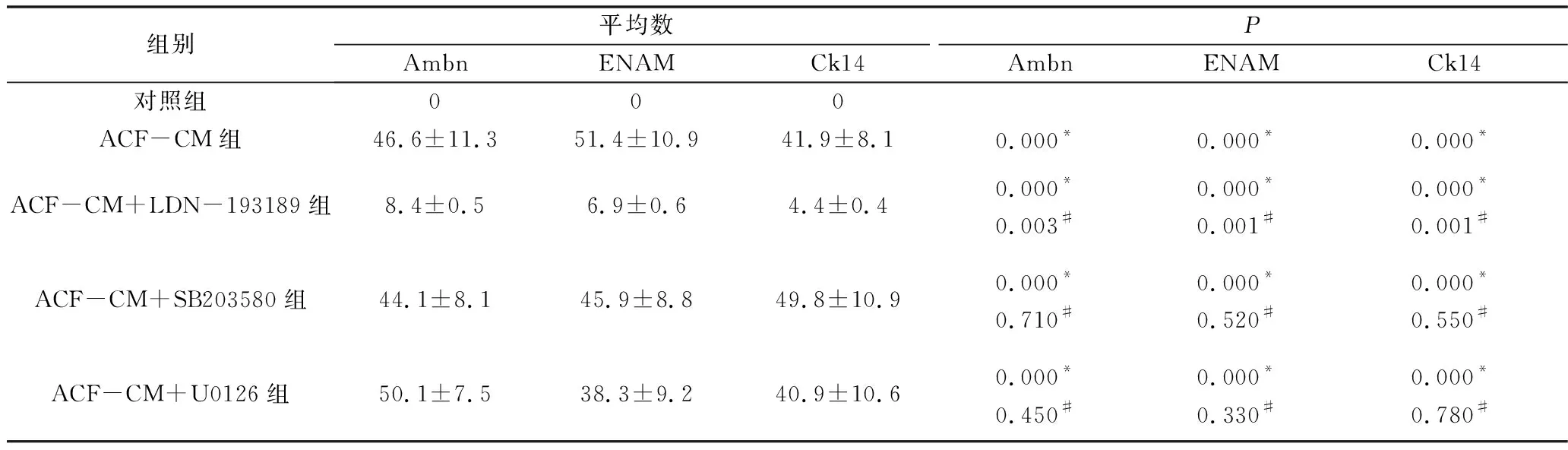
表3 免疫組化半定量比較
4.MTT結果:不同培養基組在不同時間下A值差異有統計學意義(不同培養基因素F=2.12,P<0.05,時間因素F=5.33,P<0.05,交互效應F=3.01,P<0.05)。采用對照培養基或ASF-CM培養iPS細胞1天(對照組:0.20±0.03;ASF-CM組:0.18±0.02)及7天(對照組:0.50±0.10;ASF-CM組:0.46±0.14),兩組間iPS細胞增生活動在不同時間點比較差異均無統計學意義(P>0.05),當培養14天時,ASF-CM組iPS細胞增殖活動較對照組顯著降低(對照組:0.78±0.06;ASF-CM組:0.56±0.11;P<0.05)。然而,在ASF-CM組中加入smad1/5、P38或ERK1/2信號通路抑制劑組iPS細胞增殖活動與單純ASF-CM組細胞在1、4、7天比較,差異無統計學意義(P>0.05,圖5)。
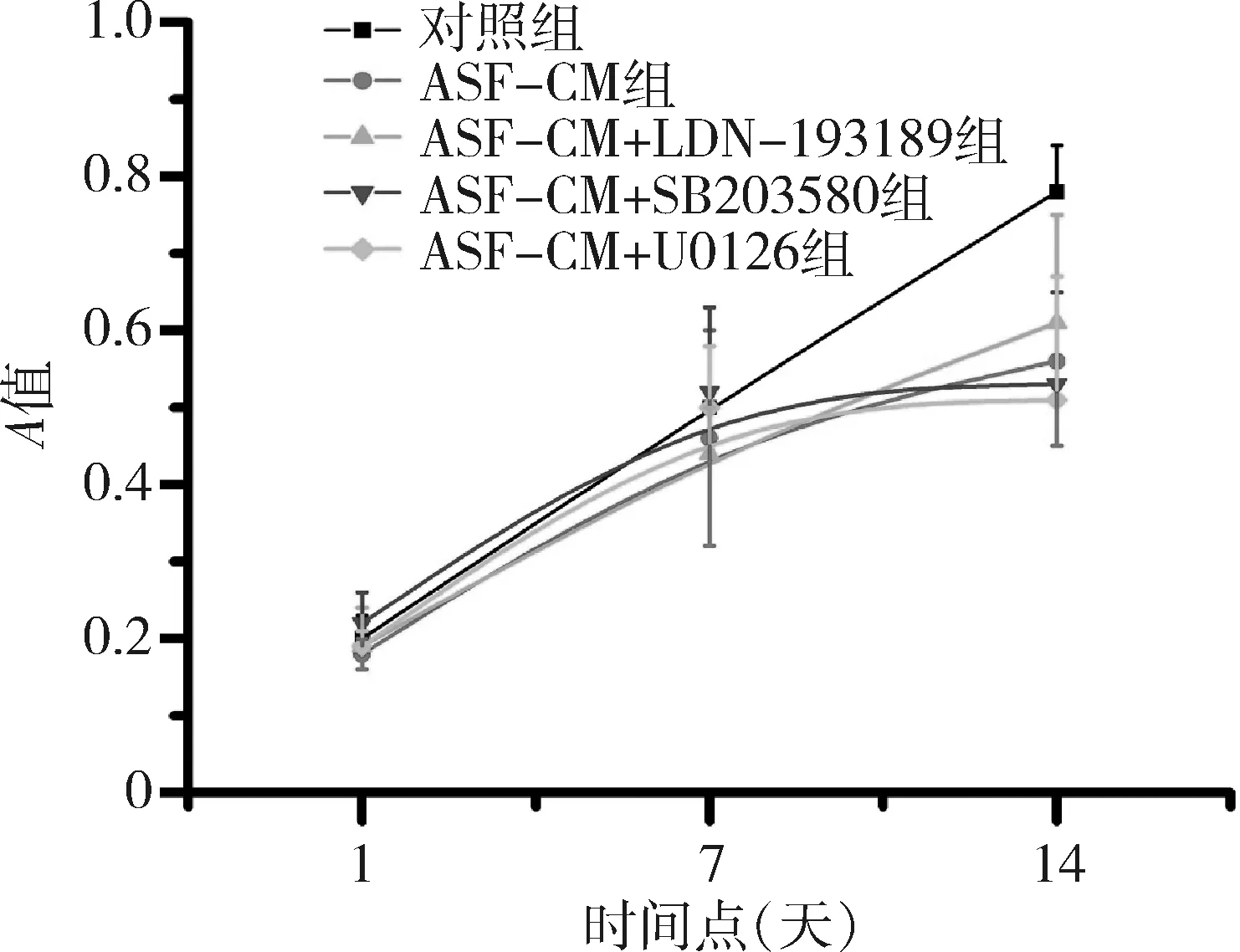
圖5 MTT實驗檢測
討 論
用于牙齒再生的干細胞主要包括牙源性干細胞和非牙源性干細胞,前者包括牙髓干細胞、牙周膜干細胞及脫落乳牙干細胞等,但由于患者特異性的牙源性干細胞的來源及細胞數量有限,限制了其在臨床上的廣泛應用[4~6]。非牙源性干細胞包括骨髓間充質干細胞、胚胎干細胞及iPS細胞等,其中骨髓間充質干細胞仍存在來源有限的問題[7];而胚胎干細胞長期受著倫理上的爭議以及免疫排斥的缺陷,也不適合應用于臨床治療[8,9]。
本實驗所采用的iPS細胞來源于成熟體細胞,因此克服了細胞來源受限、倫理和免疫排斥上的障礙,具有良好的臨床應用前景[1]。Arakaki等[10]通過分離小鼠下切牙根尖端頸環組織構建了牙源性上皮細胞系,并將該上皮細胞與鼠源性iPS細胞共培養,結果顯示上皮細胞可顯著誘導iPS細胞分化為成釉細胞樣形態,并表達Ambn、ENAM及Ck14等多種成釉細胞標志物;然而,Yoshida等[11]研究發現,相較于將Malassez上皮剩余細胞與iPS細胞共培養,采用Malassez上皮剩余細胞的條件培養基培養iPS細胞能夠更有效地誘導其形成成釉細胞樣細胞。Liu等[2]證明了成釉細胞無血清條件培養液(ASF-CM)可高效誘導iPS細胞向成釉細胞方向分化,表達成釉細胞標志物Ambn及Ck14。本實驗結果顯示ASF-CM 培養iPS細胞14天后,其成釉細胞標志物Ambn、ENAM和Ck14基因和蛋白表達水平均較對照組增高,這與Liu等[2]之前的結果一致。這些結果表明,利用成釉細胞條件培養基培養iPS細胞可誘導其分化為更接近成釉細胞的成熟細胞。牙齒的發育和形成是非常復雜生理過程,受到多種生長因子的調控,而這種動態過程不太可能僅由某種因子進行控制[12~15]。因此,本研究采用ASF-CM誘導iPS細胞成釉分化,已有實驗證實ASF-CM包含BMPs、TGF-βs、Notch 1、纖維細胞生長因子等多種細胞因子[2,16~18]。然而,本研究并未鑒定本實驗所采用的ASF-CM中的活性細胞因子成分,為了保證實驗結果的重復性,實驗中將所獲得的ASF-CM混合在一起用于后續實驗,從而確保實驗的可重復性,后續實驗可采用ELISA檢測法鑒定ASF-CM中的活性細胞因子成分。
研究顯示,BMPs是有效誘導iPS細胞分化的必要生長因子[19]。盡管smad1/5的激活被認為是BMPs經典信號轉導通路,但研究顯示BMPs亦可激活MAPKs信號通路,包括p38 MAPK、ERK1/2及JNK[20]。本實驗結果顯示,成釉細胞條件培養基培養iPS細胞14天后,可顯著促進磷酸化smad1/5、P38和ERK1/2分子的表達,表明上述通路被激活。進一步信號通路抑制劑阻斷實驗結果顯示,僅smad1/5抑制劑LDN-193189可顯著反轉ASF-CM促iPS細胞成釉分化的效應,而p38 MAPK及ERK1/2通路抑制劑對ASF-CM促iPS細胞成釉分化的效應均無顯著影響。上述結果表明smad1/5信號通路在ASF-CM誘導的iPS細胞的成釉分化過程中發揮主要調控作用。后期實驗中應采用體內功能驗證實驗進一步證實smad1/5信號通路在iPS細胞成釉分化過程中的作用。
P38 MAPK通路已被證實在BMP2及BMP7介導的牙齒發育中發揮重要調控作用[21,22]。例如調控胚胎期釉質結節p21的表達,以及成釉細胞成釉蛋白、釉質素及整合素-4的表達;外胚層P38敲除小鼠則表現出牙釉質缺陷和牙尖缺失[23]。ERK 1/2通路被證實在TGF-β1介導的組織增長,膠原蛋白改建以及牙源性干細胞分化發揮關鍵調控作用。由于ASF-CM中含有BMP2、BMP7、TGF-β1等細胞因子,本實驗在后續研究中將進一步探明P38及ERK 1/2通路在ASF-CM誘導iPS細胞成釉分化中的作用。
綜上所述,本研究結果顯示smad1/5信號通路在ASF-CM誘導的iPS細胞成釉分化過程中發揮關鍵調控作用。后期需要體內實驗進一步證實smad1/5信號通路對iPS細胞成釉分化的影響及相關機制。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
