miRNA參與益生菌調節腸道屏障功能的研究進展
2020-10-29 06:17:40李艾黎楊佳杰馬向陽賈新棟
食品科學 2020年19期
關鍵詞:小鼠
李艾黎,張 欣,李 穎,楊佳杰,馬向陽,賈新棟,杜 鵬
(東北農業大學食品學院,黑龍江 哈爾濱 150030)
益生菌作為一類可對宿主健康產生有益影響的微生物,可通過調節腸道微生態環境來改善精神健康狀況、降低膽固醇、改善人體胰島素水平、調節免疫因子、輔助過敏治療等[1-4]。常用的益生菌如雙歧桿菌、嗜酸乳桿菌、植物乳桿菌等憑借其安全可靠、性能優良的生理特性備受消費者關注和青睞,已被廣泛應用于食品、醫藥和飼料工業。預計2020年益生菌膳食補充劑和益生菌非處方藥的國內市場規模將達到850億 元[5]。
越來越多的研究發現益生菌與腸道屏障的生理狀態以及病理性改變密切相關,有關益生菌保護和調節宿主腸道屏障的作用機制不斷被揭示。如Jang等發現短乳桿菌G-101可顯著下調結腸炎小鼠腸道內細胞炎性因子細胞白介素(interleukin,IL)-1β、IL-6和腫瘤壞死因子(tumor necrosis factor-α,TNF-α)的水平,增強腸道黏膜免疫功能[6]。Yi Hongbo等觀察到羅伊氏乳桿菌LR1通過恢復仔豬腸道緊密連接蛋白表達,修復致病性大腸桿菌破壞的腸上皮屏障[7]。miRNA(microRNA)是一種廣泛參與細胞生理及病理活動的非編碼小RNA分子,主要通過特異性的堿基配對與靶基因mRNA結合,在轉錄后水平上對基因表達進行調控。miRNA在腸道組織中表達極為豐富,不僅是腸道生長、發育和黏膜屏障的重要調控環節,而且在腸道免疫耐受和防御病原微生物感染方面也發揮重要作用[8]。最新的體外細胞和體內動物實驗提示,益生菌保護或恢復腸黏膜屏障功能的作用亦與調節miRNA的表達密切相關。本文就miRNA參與益生菌保護腸道屏障和腸道穩態的相關研究進行文獻收集和整理,擬從miRNA的作用機制及其在腸道疾病中的差異表達、益生菌調節腸道屏障功能和miRNA參與益生菌調節腸道屏障這3 個主要方面展開綜述,旨在從miRNA角度分析益生菌對腸道上皮屏障功能的調節作用及其分子機制。
1 miRNA與腸道屏障
1.1 miRNA的生成及作用
miRNA是一類約由19~22 個核苷酸組成的內源性單鏈非編碼小分子RNA,具有高度的保守性和組織器官特異性,參與腸道上皮細胞的增殖、分化以及腸道免疫功能[9]。miRNA編碼基因在細胞核內被核酸內切酶裂解,形成含莖環結構的前體Pre-miRNA,隨后該前體從細胞核中轉移到細胞質,并在內切酶作用下形成短的雙鏈RNA,雙鏈中的前導鏈結合RNA誘導的沉默復合體產生具有生物活性的成熟miRNA。miRNA通過結合靶基因促進靶基因的mRNA降解并抑制其蛋白的表達,從而在轉錄后水平調控靶基因的表達[10]。具體機制與靶基因結合程度有關,如果miRNA與靶mRNA匹配完全,則該復合體降解mRNA;若兩者序列部分匹配,如miRNA的5’端2~8 個被稱為種子序列的核苷酸與靶mRNA完全匹配,則通過抑制靶mRNA的翻譯來沉默特定基因。此外,某些miRNA能夠特異結合于靶基因的富含腺嘌呤/尿嘧啶元件的3’端非翻譯區,從而指導Ago蛋白與鋅脂蛋白結合成沉默復合體區,進而改變相應mRNA的半衰期,加速靶mRNA的降解[11-12]。
1.2 miRNA在腸道疾病中的異常表達
炎性腸病、腸道腫瘤等腸道疾病存在不同程度的腸道屏障損傷,宿主體內異常表達的miRNA可能是誘因之一。如炎癥性腸病(inflammatory bowel disease,IBD)患者腸道黏膜中激增的Th17細胞、內皮細胞、巨噬細胞等產生大量促炎細胞因子(如TNF-α、IL-1β、IL-6、IL-23等)可造成腸道屏障損傷[13]。Wu Feng等通過微陣列、實時熒光定量聚合酶鏈式反應和原位雜交分析發現,與健康組相比,在結腸炎患者結腸中有11 個miRNA差異表達,其中miR-192表達水平顯著降低,該miRNA可靶向下調免疫調控因子巨噬細胞炎癥蛋白2的表達,導致炎性細胞因子TNF-α大量產生破壞腸道屏障完整性,從而加重患病癥狀[14]。Xue Xiaochang等發現IBD患者腸黏膜組織中表達水平降低的miR-10a可負反饋調節相應靶基因,致使IL-23分泌激增[15]。Polytarchou等觀察到IBD患者體內過表達的miR-214激活了轉錄調控因子STAT3,進而促進IL-6產生[16]。另外,IBD患者體內上調的miR-223可通過靶向抑制緊密連接蛋白相關基因CLDN8表達,破壞腸道上皮細胞屏障[17]。
此外,結直腸癌(colorectal cancer,CRC)患者腸道癌變細胞大量快速增殖會破壞其細胞組織,進而造成嚴重腸道屏障損傷[18]。迄今已發現百余個與CRC密切相關的miRNA[19],如Slattery等對217 例CRC病例分析后發現,CRC患者腸道內的11 個異常基因與37 個異常表達的miRNA相關,特別是miR-150-5p和miR-196b-5p[20]。進一步的研究提示,miRNA可通過影響腫瘤生成通路及其相關蛋白表達進而影響腫瘤增殖、侵襲和轉移。如miRNA-106a、miRNA-92可通過激活磷脂酰肌醇-3激酶/蛋白激酶B
(phosphatidylinositol-3 kinase/protein kinase B,PI3K/AKT)信號通路促進結腸癌的轉移[21-22]。而miRNA-145則可通過擾亂結腸癌細胞內Wnt/β-連環蛋白(wingless integrated/β-catenin,Wnt/β-catenin)信號通路的轉運抑制早期細胞癌變。此外,miRNA-145還可通過破壞連環蛋白δ-1的核穿梭功能,致使β-catenin核轉運異常而發揮抑癌作用[23]。
另有實驗利用過表達和干擾工具觀察miRNA協同其靶基因的負反饋功能,評估miRNA作為生物靶點修復腸黏膜損傷的潛在作用。如Valeri等發現沉默miRNA-135b表達可有效抑制結腸腫瘤細胞增殖[24]。而給予2,4,6-三硝基苯磺酸誘導的急性結腸炎小鼠miR-301a或miR-141抑制劑灌腸治療,可顯著降低Th17細胞比例以及IL-17表達,防止炎癥浸潤損傷腸道屏障[25-26]。由此可見,加強對miRNA分子和腸道屏障功能之間關系的研究,有利于深層次挖掘和揭示影響機體腸道健康的機理。
2 益生菌調節腸道屏障功能
腸道屏障是機體抵抗外界有害物質的重要保障,與人體健康息息相關,主要由機械屏障、免疫屏障與生物屏障構成。其中機械屏障由腸上皮細胞、細胞間緊密連接蛋白與菌膜三者構成;免疫屏障由細胞因子、免疫球蛋白(immunoglobulin,Ig)與免疫活性細胞等共同組成;生物屏障由腸道菌群和腸上皮細胞結合產生的黏蛋白、活性肽等共同組成[27-29]。益生菌可通過調節腸道免疫、保護腸道上皮細胞和細胞間緊密連接、改善菌群失調等途徑增強腸道屏障功能[30],進而維護腸道健康。
已有研究報道益生菌通過影響免疫屏障中的p38絲裂原活化蛋白激酶/核轉錄因子-kappa B(p38-mitogenactivated protein kinase/nuclear factor kappa B,p38 MAPK/NF-κB)信號通路,降低炎癥因子IL-6、TNF-α、IL-8的轉錄與表達,減少上皮細胞凋亡,改善炎癥病變。如Kim等證實雙歧桿菌KCTC 5727抑制了TNF-α誘導的NF-κB信號通路激活,顯著緩解小鼠急性結腸炎[31]。Takeda等研究表明,副干酪桿菌06TCa19通過抑制p38 MAPK信號通路,下調IL-8水平,從而有效改善幽門螺旋桿菌患者胃腸炎癥[32]。Liu Meiling等也發現乳酸鏈球菌ML2018可通過抑制p38 MAPK/NF-κB炎癥通路,下調TNF-α、IL-1β、IL-6水平,減輕硫酸葡聚糖鈉(dextran sulfate sodium,DSS)誘導的小鼠結腸炎[33]。此外,益生菌對宿主免疫屏障的調節作用還體現在刺激宿主分泌Ig、增強體液免疫方面。如雙歧桿菌OLB6378可上調血清IgG和腸道IgA表達水平,增強低出生體重嬰兒的免疫力[34]。同樣地,副干酪乳酸菌MCC1849也可誘導腸道分泌IgA[35]。Kozakova等發現小鼠灌胃含李糖乳桿菌LOCK0900、鼠李糖乳桿菌LOCK090833和干酪乳桿菌LOCK0919的混合益生菌后,其血清中特異性IgE降低,總IgA水平提高,同時明顯改善了小鼠對花粉的敏感性[3]。
益生菌還可促進緊密連接蛋白(包括閉合蛋白(Occludin)、密封蛋白(Claudin)、閉合小環蛋白(zonula occludens,ZO)-1、2、3)表達從而保護腸道機械屏障完整性。如蔣紅利等發現,雙歧桿菌Bi-07通過上調尿毒癥大鼠腸道緊密連接蛋白Occludin和Claudin-1的表達水平來促進腸上皮屏障的完整性[36]。Guo Shuangshuang等報道嬰兒雙歧桿菌和嗜酸乳桿菌可抑制Caco-2細胞中IL-1b誘導的NF-κB信號通路活化,調節Claudin和Occludin表達,降低上皮細胞通透性,保護腸道屏障[37]。Wang Jing等研究發現植物乳桿菌ZLP001通過恢復Claudin和ZO-1表達提高斷奶仔豬腸防御肽pBD2、PG1-5和pBD2的生成,下調炎性因子IL-6、IL-8、TNF-α水平,從而改善腸道屏障功能[38]。另外,益生菌可通過修復病原菌誘導的緊密連接蛋白內吞,改善腸道屏障功能。如Jariwala等研究表明,鼠李糖乳桿菌修復了大腸桿菌O26:H11誘導的緊密連接蛋白內吞,并將緊密連接蛋白重新分布到細胞邊界,改善致病菌造成的腸道上皮功能障礙[39]。
益生菌調節腸道屏障功能的作用還體現在增加有益菌豐度、減少有害菌比列、維護菌群平衡、保護生物屏障等方面。多項給予腸易激綜合征患者或DSS誘導的潰瘍性結腸炎小鼠益生菌治療的研究表明,益生菌可明顯增加腸道菌群中有益菌(如雙歧桿菌和乳酸桿菌屬)豐度,降低有害菌(如芽孢桿菌和嗜膽菌)比例,修復失衡菌群并減少患者體內炎癥標志物,促進腸道黏膜功能恢復,使腸道炎癥得到顯著改善[40-42]。這些研究表明,應用益生菌在防治屏障功能受損相關的腸道疾病中取得良好效果。
3 miRNA參與益生菌調節腸道屏障
3.1 腸道菌群影響宿主miRNA的表達
腸道微生物主要由共生菌、條件致病菌以及病原菌組成,在調節機體營養代謝、拮抗病原菌定植、維持腸道屏障功能等方面發揮重要作用,被認為是連接基因、環境和免疫系統的重要紐帶[43-44]。最近研究表明,腸道微生物群可通過調控宿主miRNA表達來影響宿主腸道健康。如Peck等研究發現,在共生菌的影響下,腸道上皮細胞存在19 個miRNA差異表達,其中miR-375與上皮細胞增殖密切相關[45]。Nakata等比較了普通小鼠和無菌小鼠腸上皮細胞的miRNA表達譜,發現共生菌誘導的miR-21-5p在腸道上皮細胞中顯著過表達。進一步研究發現,該miRNA可靶向作用于核糖基化因子4調節緊密連接蛋白(Claudin和Occludin)表達繼而影響腸上皮通透性[46]。與此類似,Natasha等研究發現,與正常對照組小鼠相比,在無菌小鼠的盲腸中有16 種miRNA差異表達,這些miRNA的靶點均參與調節腸道屏障功能[47]。Liang Guanxiang等研究發現定植于牛腸道內的雙歧桿菌或乳酸菌的數量與miR-15/16、miR-29和miR-196的表達水平呈正相關,對淋巴組織和免疫細胞的發育有重要意義[48]。此外,一些病原菌降低機體免疫力使宿主患病也與影響miRNA表達有關。如Xue Xiaochang等發現,與空白對照組小鼠相比,大腸桿菌感染使小鼠腸道內miR-107表達降低,導致該miRNA靶向抑制的炎性因子IL-23 p19大量生成,破壞腸道穩態[49]。Archambaud等報道李斯特菌通過下調小鼠腸道miRNA(miR-192、miR-200b和miR-215)和抗炎因子IL-2、IL-10表達,上調促炎因子IL-22水平,降低小鼠免疫力從而導致小鼠感染[50]。與此相似,幽門螺桿菌和土拉弗菌可通過干擾正常miRNA表達引發宿主感染[17,51]。
此外,共生菌代謝低聚糖產生的短鏈脂肪酸(如乙酸、丙酸、丁酸等)也可通過調控腸道組織miRNA表達影響腸道屏障穩態。丁酸是重要短鏈脂肪酸之一,不僅為上皮細胞提供能量,還有抗癌功效[52]。以往研究表明丁酸鹽能抑制組蛋白脫羧酶誘導的蛋白乙酰化,增加細胞周期阻滯的關鍵調控因子p21的轉錄,從而限制癌細胞增殖[53]。新近研究發現miRNA也參與丁酸鹽的抗癌作用,如丁酸能抑制miR-106b表達、促進細胞周期抑制蛋白p21轉錄、減少腫瘤細胞增殖[54]。另一研究發現丁酸鈉通過上調HT-29細胞和Caco-2細胞中miR-203表達來誘導細胞凋亡,進一步研究發現,miR-203還可靶向腫瘤細胞轉移相關蛋白NEDD9的3’端非翻譯區,下調NEDD9表達,抑制癌癥惡化[55]。Nielsen等也發現給大鼠喂食抗性淀粉(低聚糖)后,促癌基因miR-17的表達減少,糞便中的丁酸鹽含量增加[56]。上述證據提示,腸道菌群種類及其代謝產物可通過影響miRNA表達來調節宿主腸道屏障功能。事實上,益生菌作為腸道微生物中的一部分,其對腸道屏障的調節作用也是通過介導miRNA表達實現的(表1)。

表1 益生菌調節miRNA保護腸道屏障Table 1 Probiotics regulate miRNA expression to protect intestinal barrier
3.2 miRNA參與益生菌調節腸道免疫屏障
腸道依賴于天然免疫和適應性免疫來實現免疫調節作用。天然免疫是通過抗原呈遞細胞識別病原體并呈遞給T細胞來啟動獲得性免疫應答,同時通過合成炎性因子和抗炎因子引發免疫反應[70]。特異性免疫分為細胞免疫和體液免疫。其中體液免疫是以Ig起主要作用的免疫應答反應[71]。益生菌對腸道天然免疫和適應性免疫均可產生有益影響。
3.2.1 miRNA參與調節TLR4信號通路
宿主腸道上皮細胞、樹突狀細胞和巨噬細胞等抗原呈遞細胞上存在的一系列模式識別受體(pattern recognition receptors,PRRs),可識別分布于致病菌表面的致病相關分子模式。Toll樣受體4(Toll-like receptor 4,TLR4)是一種重要的PRRs。當TLR4的胞外區域與刺激物結合后,其胞內區與髓樣分化因子88(myeloid differentiation factor 88,MyD88)結合,從而使IL-1受體相關激酶1(IL-1 receptor-associated kinase 1,IRAK1)磷酸化,進而結合TNF受體相關因子6(TNF receptor associated factor 6,TRAF6),導致轉化生長因子激酶1(transforming growth factor kinase 1,TAK-1)及TAK-1結合蛋白(TAK binding proteins,TAB)募集并形成復合物,并啟動下游信號通路:1)TRAF6通過磷酸化激活MAPK,繼而活化P38 MAPK信號通路;2)TAK-1誘導核轉錄因子抑制因子(inhibitor of nuclear factor kappa B,IκB)蛋白抑制劑磷酸化,使IκB與NF-κB復合物解離,導致NF-κB向細胞核移位,激活NF-κB信號通路[72]。上述信號轉導過程被稱為TLR4/p38 MAPK/NF-κB信號通路,該通路是機體發揮先天免疫作用的重要途徑,過度活躍的TLR4/p38 MAPK/NF-κB信號通路將常導致炎性因子大量產生,進而誘發炎癥反應[73-74]。
炎癥反應激活TLR4/p38 MAPK/NF-κB信號通路,同時誘導特異性miRNA表達;而上調的miRNA可通過靶向沉默TLR4/MyD88下游關鍵蛋白,如IRAKl和TRAF6表達,減弱NF-κB信號活化,抑制IL-6、TNF-α等炎癥因子產生[74-75]。如Zhang Quanbo等發現miR-146a缺失使得痛風關節炎模型小鼠的TRAF6、IRAK1上調,并增加IL-1、TNF-α水平[76]。此外,miR-146、miR-155、miR-21、miR-9、miR-148、miR-27a也可靶向作用于TLR4/p38 MAPK/NF-κB信號通路,參與免疫調節反應(表2)。上述成果提示,miRNA仿佛強大的“剎車片”,協同其靶基因參與負向調節TLR4信號通路中的關鍵分子,從而可能成為緩解和終止炎癥反應的治療靶點。

表2 參與調節TLR4/p38 MAPK/NF-κB信號通路的miRNATable 2 MiRNA involved in TLR4/p38 MAPK/NF-κB signaling pathway
近年來,研究者對益生菌免疫調節機制的解析已深入至分子水平,有關miRNA參與益生菌調控腸道屏障和腸道穩態的作用被不斷揭示。多數研究認為,益生菌可通過調控miRNA表達來影響TLR4/p38 MAPK/NF-κB信號通路,抑制促炎因子(TNF-α、IL-8、IL-6)合成,促進抗炎因子IL-10分泌,從而增強腸道免疫屏障功能(圖1)。如G i a h i 等報道,熱滅活的鼠李糖乳桿菌G G、德氏乳桿菌保加利亞亞種分別與脂多糖(lipopolysaccharide,LPS)處理的樹突狀細胞共培養時,均可抑制TLR4 mRNA表達,其中鼠李糖乳桿菌GG可影響miR-146表達并抑制NF-κB信號通路,德氏乳桿菌保加利亞亞種可通過調控miR-155表達抑制p38 MAPK信號通路,從而緩解炎癥反應[57]。Sabharwal等也觀察到大腸桿菌Nissle 1917與腸上皮細胞T84共培養時可調節miR-146表達,并抑制炎性因子IL-8和趨化因子CXCLI的產生[58]。Chen Qiaoling等研究發現,經植物乳桿菌ZO-1喂養后,沙門氏菌感染的雞腸道內miR-193、miR-375表達發生變化,這兩個miRNA的靶基因可靶向NF-κB信號通路的關鍵分子,進而降低炎性因子TNF-α分泌,減輕感染癥狀[60]。與此類似,熱致死副干酪乳桿菌NCC2461與外周血單核細胞共培養可抑制miR-27a表達,上調抗炎因子IL-10水平[59]。值得注意的是,Taibi等提出益生菌調節miRNA表達具有時間依賴性[61],如雙歧桿菌MIMBb75干預結腸炎小鼠的實驗中發現,表達的miR-148可通過靶向TLR4/p38 MAPK/NF-κB信號通路減少內皮PAS1蛋白基因表達及TNF-α的合成。并且,miR-148表達在益生菌干預宿主2~13 d內顯著上調,14 d后miR-148的表達水平不再變化。但這種調節作用的具體機制有待進一步闡釋。
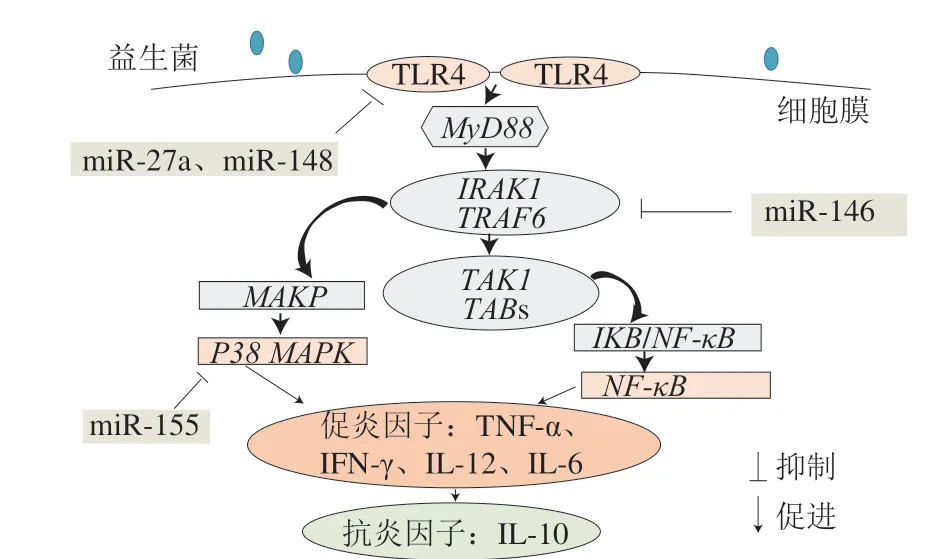
圖1 益生菌調節miRNA表達影響TLR4/p38 MAPK/NF-κB信號通路[73]Fig. 1 Regulation of miRNA expression by probiotics affects TLR4/p38 MAPK/NF-κB signaling pathway[73]
3.2.2 miRNA參與調節Ig分泌
Ig能特異性識別并結合外來分子,阻斷有害物質入侵,在體液免疫過程中發揮著主要作用。Ig由Ig重鏈(Ig heavy chain,IGH)和Ig輕鏈(Ig light chain,IGL)構成,根據恒定區的不同,IGH可分為IgG、IgA、IgM、IgD和IgE等5 類,而IGL有κ鏈和λ鏈等2 種類型[82]。新近研究顯示,miRNA可參與特異性免疫反應影響Ig分泌。如Chen Zhe等報道在變應性鼻炎小鼠體內低表達的miR-466a-3p可靶向轉錄因子GATA結合蛋白3從而抑制Th2細胞轉錄影響IgE分泌[83]。Fang Lei等發現過敏性哮喘患者體內IgE含量與miRNA-21-5p表達水平呈正相關,進一步研究發現miRNA-21-5p通過下調磷酸酶表達水平及張力蛋白同源物PTEN轉錄,刺激氣道平滑肌細胞重塑,造成病情加重[84]。Younger等研究表明高表達的miR-423可以降低Ig超家族成員1的水平,影響免疫細胞間的相互作用[85]。有關益生菌調節miRNA與體液免疫的作用機制仍然不清楚,但已有研究者從分子水平探索,并且取得了一定進展。如Kreuzer-Redmer等發現,給斷奶后給仔豬喂食富含糞腸球菌NCIM 10415的食物后,可上調仔豬腸道內miRNA-423表達,抑制IGL合成增強體液免疫,從而減輕斷奶造成的腹瀉[63]。本課題組前期研究也發現,乳球蛋白過敏小鼠灌胃嗜酸乳桿菌后,腸道miR-155、miR-21表達量下降,同時抑制了特異性IgE和炎性因子IL-6、TNF-α分泌,牛乳過敏癥狀得到緩解[62]。
3.3 miRNA參與益生菌調節腸道機械屏障
機械屏障主要由單層柱狀上皮細胞及細胞間連接復合物構成,柱狀上皮細胞使腸腔與固有層分離,在對抗毒素和腸道病原體方面發揮積極的作用。細胞間連接復合物將上皮細胞緊密結合在一起,包括緊密連接、黏著連接、橋粒和縫隙連接[86]。以下就益生菌通過介導miRNA表達調控腸道上皮細胞凋亡和緊密連接蛋白兩個方面論述益生菌對腸道機械屏障的保護作用。
3.3.1 miRNA參與調控上皮細胞凋亡
上皮細胞的增殖、分化和凋亡之間保持著良好的平衡,是一個不斷自我替代的細胞屏障,保護自身不受病原體侵襲。當大量病原菌在機體腸道內增殖時,不僅引發腸上皮細胞的異常凋亡并造成嚴重腸道屏障損傷,還會加速病原菌進入腸組織引發炎癥、導致癌變,可見控制腸上皮細胞的凋亡對維持腸道屏障的穩態和功能至關重要。目前研究人員普遍認為,線粒體途徑是主要的細胞凋亡調控通路,B淋巴細胞瘤-2(B-cell lymphoma-2,Bcl-2)家族蛋白是典型的線粒體途徑凋亡調控基因,由抑制凋亡的Bcl-2、Bcl-xl基因和促進凋亡的Bcl-2相關X蛋白(Bcl-2 associated X protein,Bax)兩大類組成。線粒體在接受到凋亡信號后,刺激Bax轉錄,釋放細胞色素c,募集下游含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,Caspase)9基因,進而活化Caspase3,導致凋亡信號轉導途徑放大最終誘導細胞凋亡[87]。
miRNA作為一種轉錄后調控因子,可通過調控凋亡基因表達影響上皮細胞凋亡。如Bian Zhen[88]和Ramsay[89]等發現,在小鼠DSS結腸炎模型中高表達的miR-150可以靶向上調轉錄因子基因c-Myb(Bcl-xl、Bcl-2的前導因子)合成,進而促進細胞凋亡。XU Haixiang等在探究miR-21參與葛根素對心肌損傷(低氧再灌注導致)的修復機制時發現,上調miR-21表達,可下調Bax和Caspase3轉錄水平,從而減少心肌細胞死亡,改善心肌缺血損傷[90]。此外,某些miRNA能促進細胞進入增殖期抑制細胞凋亡,如Chen等研究表明miR-200b過表達能夠增加細胞周期蛋白D1表達,促進細胞進入增殖期,誘導腸上皮細胞增殖[91]。
新的研究發現益生菌可通過介導miRNA表達,抑制c-Myb、Bax、Caspase3轉錄,誘導Bcl-2合成,從而發揮調控細胞凋亡的作用(圖2)。如Rodriguez-Nogales等研究發現大腸桿菌Nissle 1917通過低表達miR-150,下調c-Myb轉錄水平,抑制結腸上皮細胞凋亡,改善DSS小鼠炎癥[42]。Zununi等研究發現腸膜明串珠菌可下調miR-21、miR-200和Bcl-xl表達,增加Caspase3含量,并誘導結腸癌細胞HT29凋亡[66]。值得注意的是,益生菌影響miRNA表達具有菌株特異性。如研究人員在比較發酵乳桿菌和唾液鏈球菌對DSS小鼠治療差異時,發現只有發酵乳桿菌具有影響miR-150表達、減少DSS小鼠腸道上皮細胞凋亡的能力[64]。Kalani等報道,LPS誘導的內皮細胞炎癥,經嗜酸乳桿菌處理后,細胞內miR-21合成增加,而細胞凋亡減少[65]。以上結果提示益生菌調控細胞凋亡的機制可能是通過調節miRNA表達實現的。
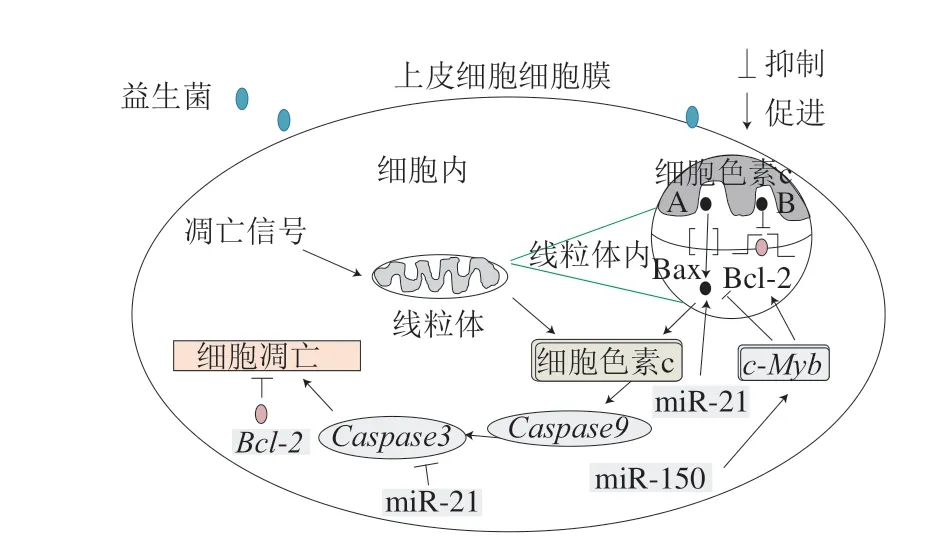
圖2 益生菌調節miRNA表達抑制腸上皮細胞凋亡[87]Fig. 2 Probiotics regulate miRNA expression and inhibit apoptosis of intestinal epithelial cells[87]
3.3.2 miRNA參與調控緊密連接蛋白
miRNA已被證實能夠調節緊密連接蛋白的表達進而影響腸道機械屏障。Ye Dongmei[92]和Zhang Bin[93]等報道,在腸道上皮細胞中,過表達的miR-122與Occludin mRNA的3’端非翻譯區結合,誘導其降解,損害腸道屏障功能導致腸通透性增加,進一步研究發現miR-122通過降低上皮生長因子受體轉錄來下調Occludin表達水平。Caballero-Garrido等發現大腦動脈閉塞小鼠經注射miR-155抑制劑后,其靶蛋白Rheb被上調,ZO-1的表達增加,血流梗死區域和微血管緊密連接的完整性得到改善[94]。極性蛋白復合體(polarity protein complex,PAR)由PAR-3、PAR-6與非典型蛋白激酶C(atypical protein kinase C,aPKC)等組成,該極性蛋白復合體與緊密連接蛋白的形成和功能維持密切相關。近年來的研究發現,miRNA可通過靶向PAR進而調節緊密連接蛋白表達。如Krissansen等發現IBD患者體內高表達的miR-595通過靶向抑制神經細胞黏附分子1和成纖維細胞生長因子受體2的表達,減少PAR-6合成,造成緊密連接成分缺失,提示miR-595可破壞上皮細胞間緊密連接蛋白完整性,加重IBD癥狀[95]。
研究表明益生菌可通過調節miRNA,促進緊密連接蛋白表達,保護腸道屏障完整性(圖3)。Veltman等報道大腸桿菌Nissle 1917與上皮細胞T84共培養時,可上調miR-203、miR-595、miR-483表達,這些miRNA分別靶向緊密連接蛋白ZO-2、PAR復合體PAR-3、PAR復合體PAR-6,從而促進緊密連接蛋白生成[67]。Zhao Haiyang等發現鼠李糖乳桿菌可上調酒精暴露下小鼠肝臟和Caco-2細胞中的miR-122a,促進Occludin表達[68]。Rodriguez-Nogales等研究表明布拉酵母菌可上調miR-155表達,促進Claudin和ZO-1合成,改善DSS小鼠腸道通透性[69]。以上結果提示益生菌可能通過調節miRNA表達維護腸道上皮屏障功能。
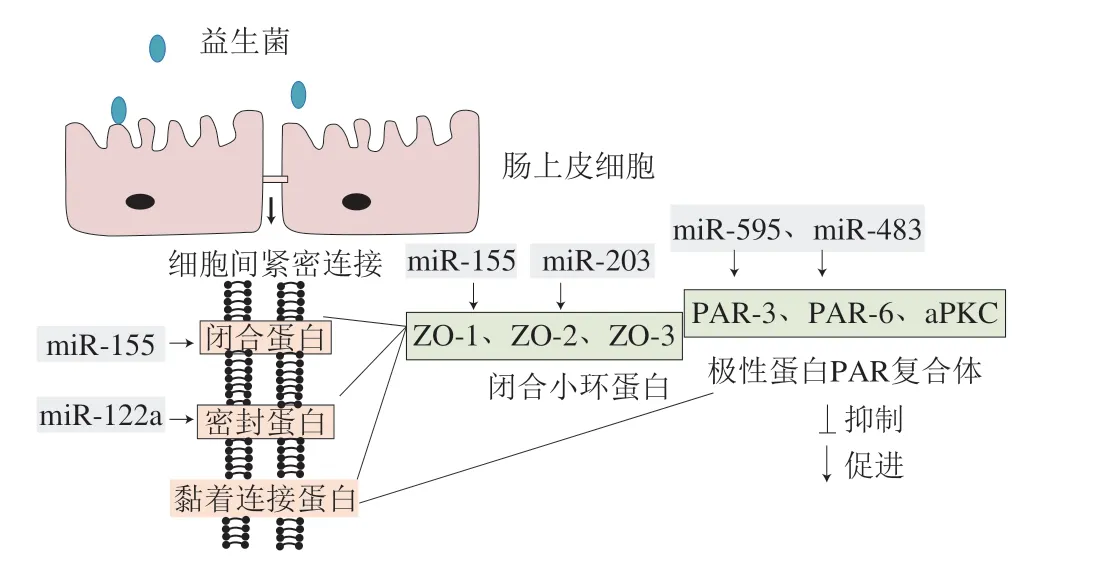
圖3 益生菌影響miRNA表達促進緊密連接蛋白轉錄[67]Fig. 3 Probiotics regulate miRNA expression and promote tight junction protein transcription[67]
4 結 語
腸道屏障是機體抵抗外界有害物質的重要防線之一,屏障功能受損將嚴重影響宿主腸道健康,并誘發各種腸道疾病。益生菌能夠保護腸道屏障的結構和功能,目前,在分子水平解析益生菌對腸道屏障的調節作用,是了解益生菌對腸道益生機制的新途徑。miRNA是一類非編碼RNA,在腸道組織中表達豐富,可通過與mRNA靶標結合發揮抑制靶基因表達的功能。近年來國內外越來越多的研究發現益生菌可通過影響miRNA表達進而調節腸道免疫,減少腸道上皮細胞凋亡,調節緊密連接蛋白表達保護腸道屏障。但miRNA受益生菌調控的機制、特定功能的miRNA對應具體哪個或哪些菌株,這些問題還需要進一步的深入研究和探討。研究益生菌調控miRNA的分子機制,加深益生菌促進健康作用的認識,將會為保護腸道健康及預防各種腸道疾病提供新的策略。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
