大麗輪枝菌蛋白激發子PevD1激活本生煙促分裂原活化蛋白激酶MAPK
2020-11-02 08:49:14賈豐蓮李澤梁穎博李廣悅楊秀芬
生物技術通報 2020年10期
賈豐蓮 李澤 梁穎博 李廣悅 楊秀芬
(中國農業科學院植物保護研究所 植物病蟲害生物學國家重點實驗室 北京 100081)
促分裂原活化蛋白激酶(MAPKs)是植物細胞內重要的信號傳導途徑。在植物與微生物互作過程中,病原物/微生物相關分子模式(Pathogen/microbeassociated molecular patterns,PAMP/MAMP)被植物細胞膜上的識別受體(Pattern recognition receptors,PRRs)識別后能快速激活MAPK級聯分子,并將底物蛋白上的絲氨酸和蘇氨酸殘基磷酸化,從而實現信號逐級傳遞。初始的級聯分子MAPKKK被磷酸化后,通過磷酸化下游的MAPKs分子,形成MAPKKK-MAPKK-MAPK級聯信號傳導途徑,MAPK進一步磷酸化下游的轉錄因子靶蛋白,導致大量基因轉錄重編程,形成復雜的免疫信號傳遞網絡,最終引起植物抗性基因表達、抗性物質積累,使植物產生廣譜抗性。MAPK通路并不是簡單的直線信號傳遞過程,大量與生長發育和防御相關的分子受到調控后形成錯綜復雜的植物信號調控網絡。盡管已明確MAPK參與調節植物生長發育以及逆境響應中的多種信號傳導途徑,是植物免疫的重要防御反應,但是目前僅明確少數MAPKs級聯分子的功能[1-2]。已經鑒定出完整的MAPK信號通路(MEKK1-MKK4/MKK5-MPK3/MPK6-WRKY22/WRKY29)的上游是鞭毛蛋白flag22激發子受體FSL2(Flagellin Sensing Locus 2)與BAK1(BRASSINOSTEROID INSENSITIVE 1-associated receptor kinase 1)復 合 體[3-4]。flg22激活的另一個MAPK級聯是MAP3K1-MAPKK1/MAPKK2-MAPK4-MKS1,該級聯介導茉莉酸(Jasmonate,JA)和水楊酸(Salicylate,SA)依賴的防御反應[5],其中MAPK4 通過其激酶底物MKS1和轉錄因子WRKY33 控制植保素camalexin合成酶PAD3的表達。不同植物MAPK通路的級聯分子功能也不盡相同,擬南芥MAPKs主要有MPK3、MPK4和MPK6,他們均能被非生物脅迫、病原菌侵染和氧脅迫等各種刺激而激活。大豆中MAPKKK(GmMEKK1)通過促進GmMPK6活化和抑制GmMPK3活化來響應flg22的刺激信號,從而不同程度地激活下游MPKs,因此GmMEKK1既能正調控又能負調控植物免疫防御反應。大量研究已經表明,MAPK級聯反應在R基因介導的抗病性中發揮重要作用,通過遺傳學手段已經在擬南芥、煙草、番茄和水稻中鑒定出參與R基因介導抗病性的MAPK組分[6-7]。水楊酸誘導的蛋白激酶(alicylic acid-induced protein kinase,SIPK)和傷誘導的蛋白激酶(Wound-induced protein kinase,WIPK)是與煙草防御反應相關的MAPKs。SIPK和WIPK的磷酸化可以激活眾多的轉錄因子,特別是WRKY家族轉錄因子,由此調控與生長發育和脅迫反應相關基因的表達。ROS能快速而強烈地激活SIPK(AtMPK6),在此過程中需要鈣離子運輸和MAPKK上游的激活。MAPKs在細菌蛋白激發子Harpin 誘導的細胞壞死中起到核心作用并能調控煙草細胞壞死反應[8]。
PevD1是本實驗室從大麗輪枝菌培養液中分離的蛋白激發子,能快速引起煙草葉片組織壞死,激發植物ROS早期防御反應,引起胼胝質,酚類化合物及木質素的積累等,提高棉花對大麗輪枝菌、煙草對煙草花葉病毒(Tobacco mosaic virus,TMV)和丁香假單胞菌的抗病性[9-10]。為了研究PevD1誘導抗病性的分子機制,前期利用轉錄組測序技術獲得了本生煙響應PevD1誘導的大量差異表達基因,其中有大量基因富集在MAPK信號傳導途徑上,本研究將進一步分析富集在MAPK信號傳導途徑中的差異表達基因的功能,研究MAPK信號途徑中重要基因的表達模式,用特異性SIPK(Salicylic acid-Induced Protein Kinase)和WIPK(Wound-Induced Protein Kinase)磷酸化位點的抗體雜交技術,明確PevD1誘導了本生煙MAPK磷酸化激活,闡述MAPK激活是PevD1誘導植物抗病性的重要機制之一。
1 材料與方法
1.1 蛋白激發子PevD1表達純化及本生煙培養
構建PevD1真核表達載體pPICZαA-PevD1,轉化畢赤酵母進行誘導表達,用Ni-NTA純化介質(ProteinIso? Ni-NTA Resin,北京全式金)對PevD1-His蛋白進行親和純化,通過SDS-PAGE和Western blot技術驗證了融合蛋白的正確性,具體方法參照文獻[11];本生煙種子(Nicotiana benthamiana)由本實驗室保存,在25℃下培養21 d的幼苗葉片用于激發子處理。
1.2 PevD1處理本生煙
將純化的激發子PevD1蛋白液稀釋成10 μmol/L后,取四周齡的本生煙植株,用無針的注射器在本生煙葉片背部滲透注射,每個葉片注射20 μL,以等量的Tris-HCl(濃度20 mmol/L,pH8.0)為對照。分別在處理后6 h、12 h、24 h葉片取樣,液氮速凍后于-80℃保存備用。
1.3 差異表達基因的篩選
根據華大BGISEQ-500平臺提供的數據,使用DEG-seq 進行差異表達基因的篩選。將PevD1處理與Tris-HCl對照之間的基因表達量差異倍數為兩倍以上(Fold Change >= 2)并且Q-value≤0.001的基因篩選為顯著差異表達基因。在PevD1分別處理6 h、12 h、和24 h的3個時間段中,只要有一個時間點的基因達到篩選標準即視為差異表達基因。
1.4 差異表達基因qRT-PCR檢測
參考茄科數據庫(https://solgenomics.net/)的目的基因序列,使用Beacon Designer 8.0軟件設計特異性引物(表1)。以上述樣品提取的RNA反轉錄后的cDNA為模板,以Actin為內參基因,采用熒光定量試劑盒(北京全式金)特異性擴增每個目的基因。參照2-ΔΔCt計算方法,分別對mRNA進行相對定量分析。

表1 差異表達基因的引物
1.5 PevD1誘導MAPK級聯反應的激活檢測
PevD1處理本生煙后,分別在處理后的5 min、15 min、30 min、60 min剪取處理部位的本生煙葉片約200 mg,液氮速凍后,提取總蛋白。總蛋白在10%SDS-PAGE電泳后取下凝膠,少量蒸餾水沖洗,凝膠放入1×Transfer buffer中,置搖床輕搖20 min;取兩張轉膜專用濾紙和1張PVDF 膜,用100% 的甲醇浸泡PVDF膜,置搖床輕搖 1-2 min;將PVDF膜轉入1×Transfer buffer,置搖床輕搖5 min,用1×Tranfer buffer 將濾紙浸透;轉膜采用濕轉法,按照以下順序組裝:負極-濾紙-膠-膜-濾紙-正極(在緩沖液中組裝,可避免產生氣泡),然后在200 mA,1 h條件下進行電轉。取出PVDF膜置于潔凈培養皿中,TBST 洗膜,搖床上輕搖 5 min,重復3次。加入封閉液(5%BSA溶液),搖床上室溫封閉3 h。TBST 洗膜,搖床上輕搖5 min;重復3次。按體積比1∶2 000以封閉液稀釋一抗(Phospho-p44/42 MAPK),加入一抗后,室溫孵育4 h;TBST 洗膜,搖床上輕搖5 min,重復3次;按1∶2 000的比例加入用TBST稀釋的二抗(Anti-rabbit IgG,HRPlinked Antibody),置于搖床上室溫孵育1 h;TBST洗膜,搖床上輕搖5 min,重復3次;將PVDF膜轉移到塑料封口膜中,加入1 mL顯色液(EasySee Western Blot,北京全式金),使顯色液與膜充分接觸1min后,用化學發光儀曝光觀察并保存圖片。進一步對采集的圖片用Image軟件進行灰度分析,定量比較對照與處理間不同時間點磷酸化差異。
2 結果
2.1 富集在MAPK通路上基因參與植物免疫多種防御反應
分析富集在MAPK通路上的差異表達基因發現,這些基因參與調控的功能十分廣泛,除了級聯分子MAPKKK和MAPKK差異表達外,還有調控基因表達的轉錄因子家族WRKY和ERF,參與識別的蛋白激酶(LRR-RLK),參與抗病的幾丁質酶(Chinase)基因。此外,還有大量參與鈣離子信號傳遞的鈣調蛋白(Calmodulin)、參與活性氧(Reactive oxygen species,ROS)產生和清除的呼吸爆發氧化酶(Rboh)和過氧化氫酶(Catalase)等(圖1,表2),這些差異表達基因的功能涉及植物免疫過程中識別-信號傳遞-防御物質積累-抗病性等各個環節。
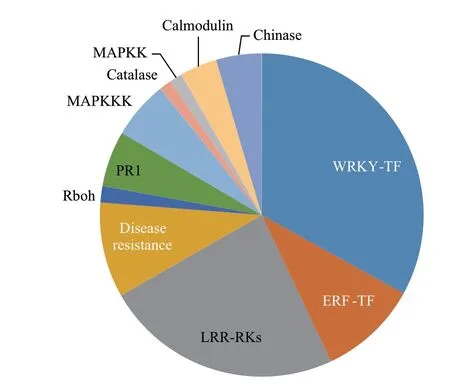
圖1 蛋白激發子PevD1誘導的MAPK通路差異表達基因的功能分布
2.1.1 PevD1誘導LRR-RLK基因上調表達 植物類受體激酶(Receptor-like kinases,RLKs)是植物免疫識別受體的重要成員,其中LRR-RLKs(Leucinerich repeat receptor-like protein kinases)是RLK中研究較多的一類家族,其結構含有識別胞外信號的受體結構域,錨定在細胞膜上的跨膜結構域和胞內傳遞信號的激酶結構域。目前已經鑒定的PAMPs受體均屬于這一類激酶,如細菌flag22受體FLS2[12],細 菌 延 伸 因 子(EF-Tu)受 體EFR[13]以 及 真菌PAMP幾丁質擬南芥受體CERK1和水稻受體CEBiP[14-16],這些受體與蛋白激酶如BAK1、BIK1、SERK或SOBIR1形成受體復合體共同識別PAMP而激活下游傳導信號,最終使植物產生廣譜抗性[17-18]。擬南芥SIF2屬于LRR-RLK,可以感知病原菌的存在并與BAK1結合,通過MAPK途徑激活下游防御相關基因的表達,從而調控植物對病原菌侵染的抗性。幾丁質作為真菌PAMP處理水稻后,OsRLCK和PBL27 參與幾丁質受體CERK1介導的信號傳導并 控 制MAPK的 激 活[10-20]。總之,LRR-RLKs在啟動植物免疫反應、產生系統抗病性中具有重要作用,但是目前僅闡述了少數LRR-RLKs的生物學功能。PevD1誘導后富集在MAPK途徑上的差異表達基因中有62個LRR-RKs顯著上調表達(圖1),說明PevD1誘導后植物啟動了大量參與識別的類受體激酶,鑒定這些LRR-RLKs將有助于闡述植物識別PAMPs后激活MAPK途徑的信號傳導機制。另外,大量研究表明LRR-RLKs轉化植物能提高植物抗病性,如花生AhRLK1轉化煙草能顯著提高對病原細菌青枯菌(Ralstonia solanacearum)抗性[21],進一步研究本實驗鑒定出的LRR-RLKs的功能,有可能為未來分子抗病育種提供有用的基因資源。
2.1.2 PevD1誘導WRKY轉錄因子基因上調表達
轉錄組數據分析顯示,PevD1誘導大量MAPK途徑上的WRKY轉錄因子上調表達。WRKY轉錄因子是植物特有的鋅指型轉錄因子蛋白家族,參與調控植物各種生理過程,包括對各種生物和非生物脅迫的防御反應。WRKY轉錄因子通過與下游靶基因啟動子上的W-box元素結合來調控靶基因的表達[22],許多WRKY轉錄因子的啟動子區域也有W-box元素,因此它們可以與自身啟動子區結合或者與其他WRKY的W-box結合,實現自我調控或者交叉調控[23-24]。WRKY33被MAPK3/6激活后,能結合在自身啟動子上,通過正向反饋環實現自動調控[25]。WRKY18、WRKY40和WRKY60 作為ABA信號的負調控子能夠直接結合到其它WRKY基因啟動子區的W-box上,從而抑制被結合的WRKY基因的表達[25-27]。WRKYs 位于各種MAPKs 的下游,通過自調控或者交叉調控實現細胞中各種WRKY蛋白家族成員間轉錄表達的平衡,并通過植物多種信號傳導途徑調整植物生長和防御之間的平衡,因此同一個WRKY在植物不同發育階段和不同生長環境下所發揮的作用也不盡相同。WIPK和SIPK共享MAPK激酶MEK2的上游[28],其下游底物是WRKY轉錄因子,病原菌侵染誘導的MEK2-SIPK/WIPK-WRKY級聯反應是植物免疫重要的信號途徑,參與類HR的細胞死亡、呼吸爆發氧化酶(Rboh)介導的ROS的產生以及對致病疫霉(Phytophthora infestans)的抗病性[29]。煙草WRKY7、WRKY8、WRKY9和WRKY11參與MEK2-SIPK/WIPK級聯下游的類HR細胞死亡,是細胞死亡依賴和非依賴型植物免疫所必需的[30]。PevD1是誘導HR的蛋白激發子,通過激發H2O2的產生和積累,提高氧化酶等防御酶活性,使細胞壁木質素含量提高、細胞壁加厚,最終提高棉花對大麗輪枝菌的抗性[10]。PevD1誘導后大量WRKY轉錄因子上調表達(圖1),預示著這些WRKY轉錄因子參與了PevD1誘導HR和H2O2的調控過程。
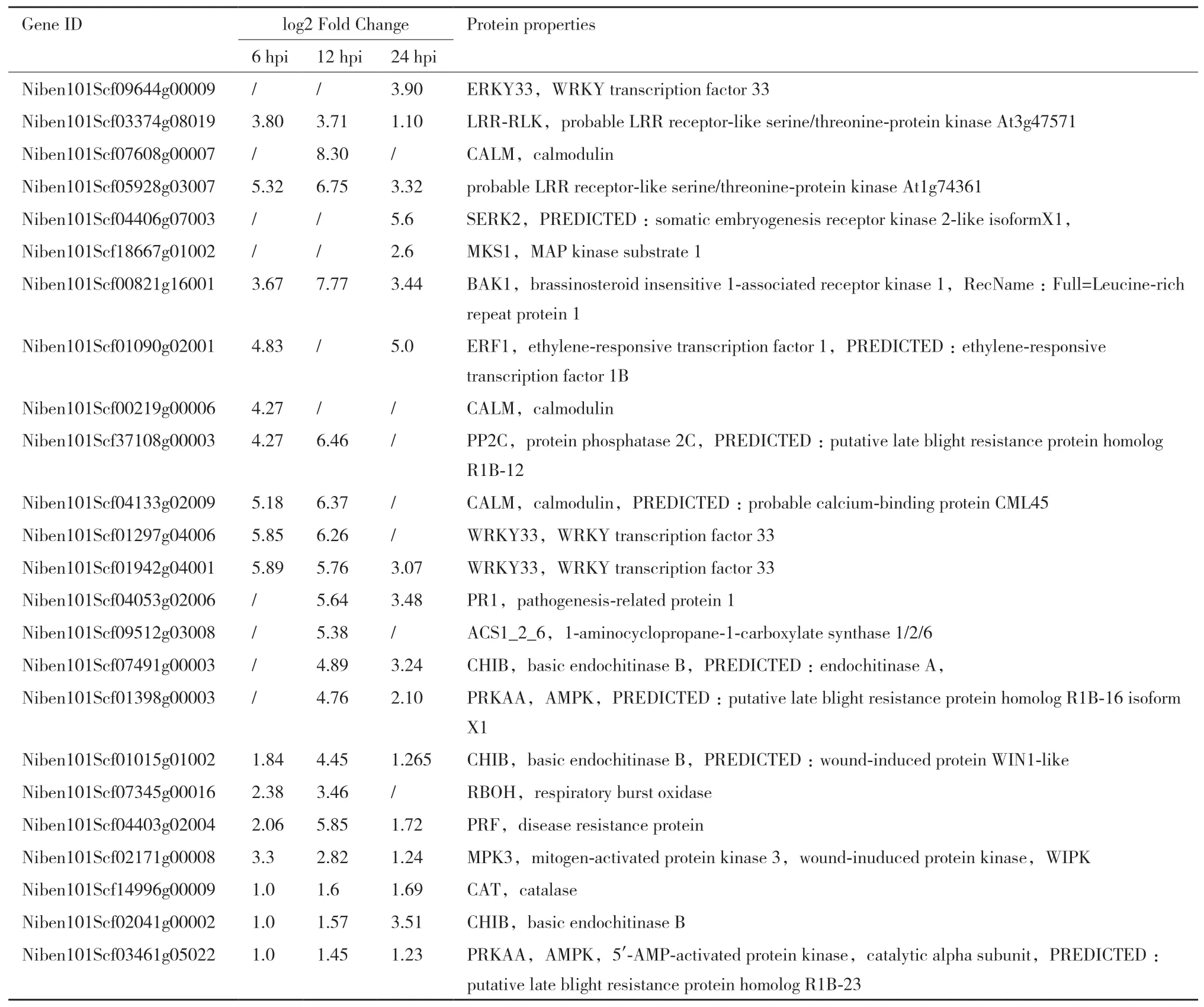
表2 富集在MAPK通路上部分差異表達基因
2.1.3 PevD1誘導ERF轉錄因子基因上調表達 ERF(Ethylene Responsive Factor)是另一個非常重要的轉錄因子家族,其家族成員參與植物生長發育和各種生物和非生物脅迫反應包括干旱、高鹽、高溫和病害等。ERF一般與靶基因啟動子區的GCCGCC序列結合調控基因的表達。煙草中NtERF3具有誘導煙草葉片細胞死亡的功能,在細胞壞死信號途徑中位于NtSIPK/NtWIPK和NtWRKY1的下游,通過反饋調節發揮其調控功能[31]。一些ERF基因在植物中表達可誘導PR基因的表達,使植物產生對細菌、真菌或病毒的廣譜抗病性,如在煙草中過表達GbERF2能提高對褐斑病抗性[32],Tsi1能提高辣椒對病毒、細菌和卵菌的侵染[33],大豆的GmERF5能提高對大豆疫霉的抗性[34]。在煙草中過表達GmERF3能提高對鹽、干旱和植物病害的抗性[35]。PevD1誘導了大量的ERF轉錄因子基因上調表達,說明這些ERF轉錄因子參與了PevD1誘導的細胞壞死和抗病性,深入研究這些轉錄因子的調控功能,不僅對揭示PevD1與植物互作分子機制具有重要意義,而且有望成為植物抗病育種新的基因資源。
2.1.4 PevD1誘導呼吸爆發氧化酶基因Rboh上調表達 植物呼吸爆發氧化酶(Respiratory burst oxidase homologue,Rboh)又稱NADPH氧化酶,在植物生長發育及脅迫反應中起重要作用,特別是在植物抗脅迫反應中的功能備受關注。在植物識別病原菌或PAMP后產生的各種防御反應中,ROS是最早發生的防御反應。ROS不僅具有直接抗菌活性,還能作為信號分子進一步激活免疫防御途徑和各種生物過程。植物Rboh氧化酶的主要功能是催化產生ROS,采用病毒誘導基因沉默(VIGS)的方法沉默NbRbohA或NbRbohB基因的植株ROS 生成量降低,同時對致病疫霉菌的抗性降低,且HR受到抑制[36]。Rboh基因目前已在擬南芥、水 稻、煙草和番茄等約10個物種中克隆并進行相關功能研究,其活性主要受Ca2+、蛋白磷酸化、Rac蛋白及ABA所調控[37]。研究發現Rboh 蛋白的活性還受Ca2+和磷酸化的協調調控,擬南芥的Ca2+依賴蛋白激酶CPK4、CPK5、CPK6和CPK11是PAMP誘導ROS的正調控子[38],CPK5通過磷酸化來調控RbohD的活性。大量研究已經證實,MAPKs(SIPK/ AtMPK6)是被ROS快速而強烈激活并依賴于RbohD[39]。本研究中PevD1誘導大量Rboh基因上調表達,而且大量編碼鈣調蛋白(Calmodulin)的基因也大幅度轉錄上調(圖1),預示PevD1誘導產生大量的Ca2+,從而激活RbohD產生ROS。誘導產生的ROS可能通過調控氣孔關閉、細胞壁加厚、胼胝質積累等阻止病原菌的侵入,最終使植物產生抗病性。深入研究調控Rboh的分子機制,將為進一步闡述激發子誘導的ROS及其傳遞網絡提供重要理論依據。
2.2 MAPK通路差異表達基因的表達模式
為了進一步對RNA-Seq的結果進行驗證,從富集在MAPK通路中的差異基因里選出10個基因進行了qPCR驗證。結果顯示,大部分基因在誘導后12 h轉錄水平達到高峰,隨著時間點推移轉錄水平逐漸下降。不同基因表達模式可能與其行使的調控功能有關,如ACS6是1-氨基環丙烷-1羧酸合成酶6,在MAPK途徑中被SIPK/WIPK6磷酸化,參與植物次級代謝產物乙烯的合成,誘導后36 h出現轉錄高峰。ERF1B是響應乙烯的轉錄因子也是在MAPK途徑的下游,調控次級代謝產物的合成,在誘導后24 h出現轉錄水平高峰。所選擇的10個基因的表達模式在表達趨勢上與RNA-Seq結果一致(圖2),說明PevD1誘導轉錄組測序結果是準確的。
2.3 PevD1快速激活MAPKs
植物中的MAPK基因在數目上比較保守,在煙草中已鑒定了17個MAPK基因家族成員,研究較普遍的是SIPK和WIPK。最早發現SIPK能夠在外源水楊酸存在的煙草懸浮細胞中被誘導表達,WIPK則主要被病原激發子誘導表達[40]。后來的研究發現,真菌蛋白激發子隱地蛋白cryptogein能誘導激活SIPK和WIPK,并伴隨著產生HR和防御基因的表達,說明煙草SIPK和WIPK參與植物免疫信號的傳導。本文用特異性檢測SIPK和WIPK磷酸化位點的抗體來檢測PevD1誘導前后SIPK和WIPK磷酸化反應的差異。從圖3可以看出,PevD1誘導5 min后即觀察到了SIPK和WIPK的磷酸化現象,隨著時間的延長SIPK和WIPK的磷酸化程度逐步增強,并在15 min左右達到最高峰,說明PevD1激活了煙草MAPK,推測可能通過激活MAPK途徑進行免疫信號的傳導,最終使植物產生系統抗病性。
3 討論
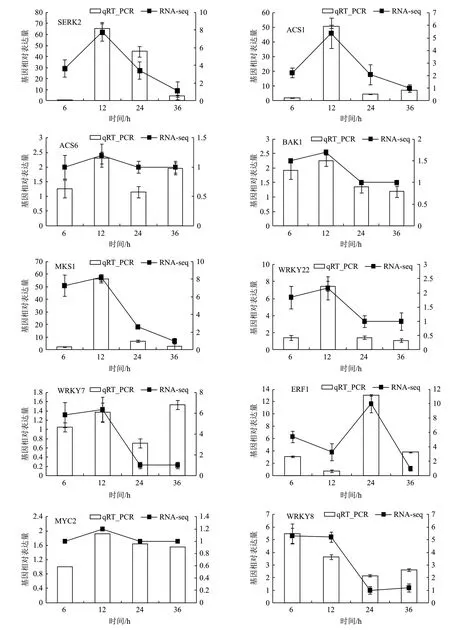
圖2 MAPK通路相關差異表達基因的qPCR驗證
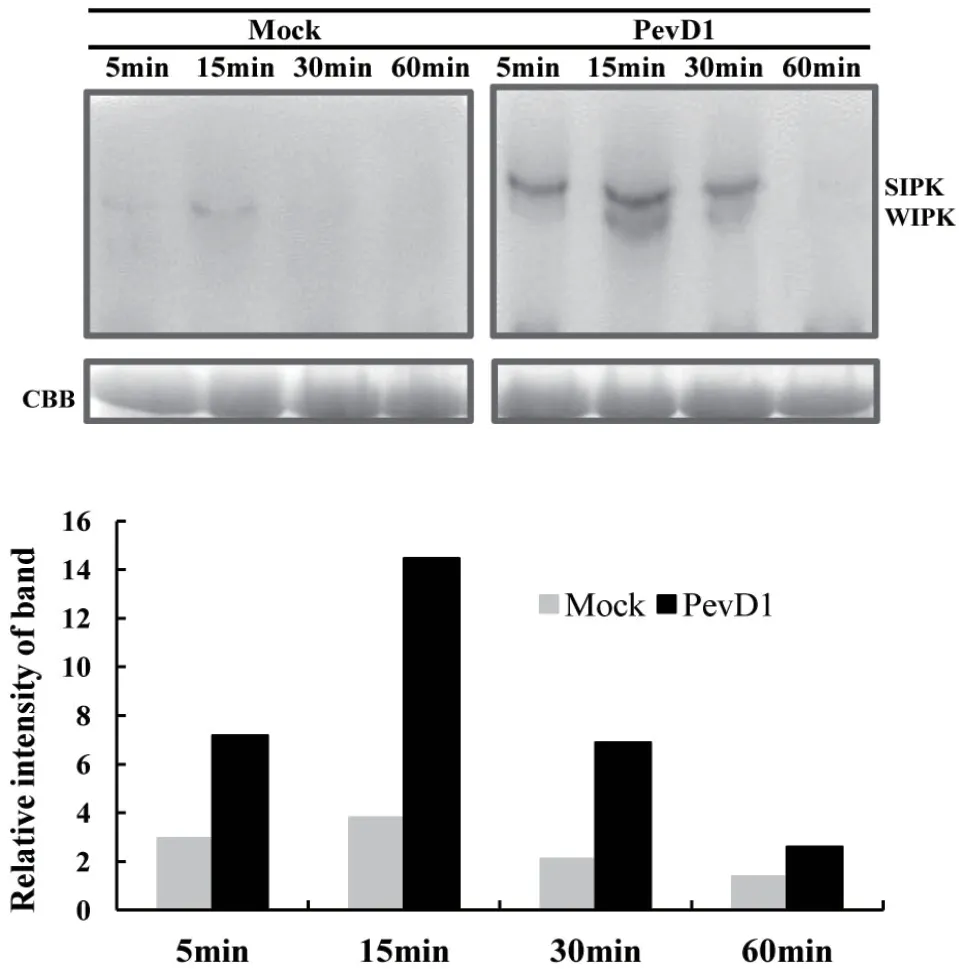
圖3 PevD1處理本生煙后不同時間MAPK激活檢測
PRRs識別外界脅迫信號是植物啟動免疫防御系統的開關,根據PRRs的細胞定位,植物識別受體包括兩類,一類是位于細胞質中的受體,通常是R蛋白,能特異性識別病原微生物產生的效應蛋白(Effectors),由此啟動ETI植物免疫反應,使植物產生較強的抗性;另一類是位于植物細胞表面,能識別胞外的MAMP/DAMP(Damage-associated molecular patterns),從而啟動PTI植物免疫反應[41],使植物產生較持久的抗性。PTI和ETI有類似的防御反應,如HR、ROS、離子流變化、原生質體滲漏、MAPK激活、激素和植保素積累、細胞壁加厚等。盡管已經明確了植物PRRs識別PAMP/DAMP是激發植物免疫防御反應的開關,但是識別后如何進行下游信號傳遞一直沒有詳細闡述,普遍認為,RLK/RLP是連接PRR與MAPK的紐帶[42]。本研究鑒定了眾多的RLK/RLP和各種轉錄因子,進一步研究它們的功能將有助于闡述病原菌/激發子被植物PRR識別后的信號傳遞以及MAPK下游轉錄因子之間的調控網絡。煙草MAPK途徑中的SIPK和WIPK不僅能被病原菌/激發子磷酸化激活,還能誘導煙草產生HR,其下游靶標NtWRKY1通過與啟動子區W-boxes結合來調控轉錄因子NsERF3[43],可見MAPK信號傳導途徑不僅與植物識別產生HR有關,還通過轉錄因子以及轉錄因子之間相互作用來調控多個防御基因的表達,形成復雜的調控網絡來響應病原菌的侵染或激發子的刺激。本文證明了PevD1能誘導煙草SIPK和WIPK磷酸化激活,但如何進行下游信號傳遞以及調控機制尚不清楚,深入研究這些響應PevD1的差異表達基因將對鑒定植物免疫信號傳導網絡中的關鍵節點具有重要的指導作用。
4 結論
本研究結果表明,PevD1誘導后使本生煙大量基因轉錄重排,通過激活MAPK信號傳導途徑,導致抗性基因上調表達,最終使植物產生抗病性。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
