GTPBP4基因對結腸癌HCT116細胞生物學行為及裸鼠成瘤能力的影響及機制
2020-12-02 01:12:20周曉青楊志惠
臨床與實驗病理學雜志 2020年10期
唐 丹,段 怡,周曉青,楊志惠
2018年全球癌癥調查數據顯示,結直腸癌(colorectal cancer, CRC)發病率位居惡性腫瘤的第3位,病死率已躍居第2位[1-2]。CRC治療通常以化療為主,但化療毒性高,機體反應相對較低。因此,研發并探尋新的治療途徑尤為關鍵[3]。分子靶向治療是干預腫瘤細胞內信號傳導通路中一些關鍵分子,阻斷信號通路的傳導,起到抑制腫瘤細胞生長的作用,并且對周圍正常細胞毒副作用較小。在發揮抗腫瘤作用方面,由于分子靶向藥物的快速發展和臨床應用,腫瘤的系統性治療已經取得重大進展[4]。CRC的特征是具有遺傳多樣性,因此從CRC發病機制的不同角度探尋新的分子靶點尤為關鍵。GTP結合蛋白4(GTP binding protein 4, GTPBP4)基因,即核仁G蛋白(nucleolar G protein, Nog1)、腦紅蛋白(neuroglobin, NGB)、慢性腎功能衰竭基因(chronic renal failure gene-1, CRFG)。GTPBP4基因定位于染色體10p14-p15,編碼的GTPBP4蛋白廣泛表達于從錐蟲到人類的真核生物細胞核,是核仁中重要的功能蛋白[5]。在本課題前期的體外實驗中發現,沉默GTPBP4基因后,結腸癌細胞株RKO[6]及HT29[7]發生G0/G1期阻滯,細胞凋亡率增加,細胞增殖受抑。原癌基因c-Jun屬于Jun家族,是激活蛋白-1(activating protein-1, AP-1)的重要組成部分[8],并且其與眾多腫瘤關系密切。有研究表明,由LMP1誘導的c-Jun/Jun B異二聚體可以上調Cyclin D1啟動子的活性和表達,Cyclin D1的過表達加速了細胞周期的進展。結合前期的實驗結果,以及通過進一步實驗來驗證沉默GTPBP4基因對結腸癌細胞裸鼠成瘤能力的影響,并探究其與原癌基因c-Jun之間的聯系。
1 材料與方法
1.1 主要材料人結腸癌HCT116細胞株購自中國科學院上海細胞庫;shRNA慢病毒載體購自上海吉凱基因化學公司;細胞周期、細胞凋亡試劑盒購自BD公司;CCK-8試劑購自碧云天公司;4~6周齡體重16~18g BALB/c裸鼠購自成都達碩實驗動物公司,許可證號SCXK(川)2015-030;兔抗人GTPBP4單克隆抗體、兔抗人Cyclin D1單克隆抗體購自Abcam公司;鼠抗人BCL-2單克隆抗體、鼠抗人Bax單克隆抗體、c-Jun兔抗人單克隆抗體購自CST公司;倒置顯微鏡購自Olympus公司。
1.2 方法
1.2.1穩定低表達GTPBP4細胞株篩選 消化對數生長期的HCT116細胞并以每毫升5×104個的細胞密度接種于6孔板中;對6孔板細胞進行慢病毒感染(MOI=20);感染72~96 h后,倒置熒光顯微鏡下觀察熒光表達。確定細胞表達綠色熒光后,將細胞培養基更換為含有2 μg/mL嘌呤霉素的完全培養基,每3天更換1次含嘌呤霉素的培養基,篩選維持3周左右。qRT-PCR及Western blot實驗檢測穩定細胞株GTPBP4基因沉默效率。
1.2.2平板克隆形成實驗 取處于對數生長期的各組細胞,以每孔200個細胞數接種于6孔板中,每組種板設3個復孔。于37 ℃ 5%CO2恒溫培養箱培養2~3周。培養皿中生長出肉眼可見的細胞集落時,終止細胞培養,以4%多聚甲醛溶液固定6孔板中的細胞15~30 min。500 μL結晶紫溶液,室溫下染色15 min,洗去多余染色液,干燥后計數拍照。克隆形成率=(克隆數/接種細胞數)×100%。
1.2.3裸鼠皮下結腸癌移植瘤模型建立 將10只裸鼠隨機分為兩組后編號,飼養于SPF級動物房。胰酶消化收集對數生長期陰性對照組及實驗組細胞,調整細胞密度為每毫升1×107個,消毒裸鼠右側腋背部皮膚,按每只裸鼠0.2 mL體積接種,沿皮下斜行進針,注射完畢后可見直徑0.5~0.8 cm皮丘形成,表示接種成功。當接種部位形成肉眼可見瘤結節時,每間隔3天用游標卡尺測量瘤體最長徑(a)及與之垂直的短徑(b),最終瘤體體積按照公式V=1/2ab2計算。記錄并分析數據。接種4周后采用脫頸椎法處死裸鼠,剝離皮下瘤結節,拍照后稱量瘤重。瘤組織用10%中性福爾馬林固定,以便行常規HE及免疫組化染色。
1.2.4Western blot法 RIPA裂解細胞提取蛋白,BCA法測定蛋白濃度。加入蛋白上樣緩沖液后沸水中煮沸5~10 min使蛋白變性。配置10%分離膠及5%濃縮膠,濃縮膠恒壓80 V跑膠30 min,分離膠恒壓110 V跑膠90 min。切下相應分子量的膠,以200 mA恒流4 ℃電轉30~90 min。5%牛奶封閉液室溫封閉1 h。4 ℃一抗孵育過夜。次日TBST洗凈多余一抗后,二抗室溫孵育1 h。滴加ECL試劑,于成像儀中曝光成像。Image J軟件分析各條帶灰度值。
1.2.5免疫組化 實驗采用免疫組化EnVision法檢測蛋白表達。4~6 μm厚石蠟切片經二甲苯脫蠟,梯度乙醇水化;按照各抗體說明書選擇檸檬酸鹽緩沖液或EDTA進行抗原熱修復;1 ∶100~1 ∶200稀釋一抗,滴加于組織上,室溫孵育4 h;即用型二抗均勻滴加于組織上,室溫孵育30 min;DAB顯色,蘇木精復染細胞核。乙醇脫水,二甲苯透明,中性樹膠封固。
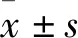
2 結果
2.1 HCT116穩定細胞株沉默效率將嘌呤霉素篩選3周后的實驗組(shGTPBP4)、陰性對照組(shControl)的兩組細胞置于倒置熒光顯微鏡下觀察,見兩組細胞均完全表達GFP綠色熒光蛋白。進一步檢測細胞GTPBP4 mRNA相對表達量及蛋白表達,實驗組GTPBP4 mRNA相對表達量為0.074 2±0.004 4,陰性對照組GTPBP4 mRNA相對表達量為1.003 2±0.099 9,實驗組的表達量明顯低于陰性對照組,差異有統計學意義(P<0.01,圖1)。實驗組穩定細胞株GTPBP4蛋白的表達明顯低于陰性對照組。由兩組相對mRNA表達量計算得出實驗組GTPBP4基因沉默效率為92.60%。

圖1 HCT116穩定細胞株沉默效率檢測:A.qRT-PCR法檢測兩組細胞mRNA相對表達量,**P<0.01;B.Western blot法檢測兩組細胞GTPBP4蛋白表達
2.2 細胞增殖活性檢測將兩組穩定轉染細胞株接種于96孔板中檢測不同時間點450 nm處OD值,接種24、48、72、96 h的陰性對照組OD值分別為0.764 1±0.021 9、1.470 1±0.148 4、2.221 5±0.146 9、3.127 4±0.020 6,實驗組OD值分別為0.536 6±0.029 8、0.899 9±0.023 3、1.152 7±0.050 8、1.687 8±0.341 0,各時間點實驗組OD值均低于陰性對照組,差異有統計學意義(P均<0.001,表1,圖2)。并由此可計算實驗組24、48、72、96 h的細胞增殖平均抑制率分別為29.77%、38.79%、48.11%、46.03%。

表1 兩組不同時間點的OD值

圖2 CCK-8法檢測兩組細胞各時間點的增殖曲線
2.3 沉默GTPBP4基因對HCT116細胞克隆形成能力的影響在細胞平板集落形成實驗中,陰性對照組細胞集落形成數目為(127.00±12.49)個,克隆形成率為63.5%;實驗組細胞集落數目為(15.33±5.51)個,克隆形成率為7.67%,兩組相比,實驗組克隆形成率明顯降低,兩組相比差異有統計學意義(P<0.01,圖3)。

圖3 陰性對照組及實驗組細胞集落形成
2.4 沉默GTPBP4基因對HCT116細胞裸鼠成瘤能力的影響
2.4.1裸鼠移植瘤瘤體體積生長曲線 接種6天后可見皮下結節形成,隨著結節的增長,兩組裸鼠平均體重減少,消瘦明顯,陰性對照組瘤結節最大者于第23天死亡,剩余4只;實驗組裸鼠消瘦明顯,5只全部存活。每間隔3天測量記錄并計算瘤體體積,繪制生長曲線(圖4),結果顯示,于第6天至第26天合計6次測量兩組瘤體體積,自測量第10天起實驗組皮下瘤體體積均較陰性對照組小,兩組相比差異有顯著性(P均<0.05),且隨著時間的增長,實驗組瘤體積較陰性對照組減小更為明顯。

圖4 陰性對照組及實驗組細胞裸鼠瘤體體積生長曲線
2.4.2裸鼠移植瘤瘤體質量 接種細胞后的第28天用脫頸椎法處死裸鼠,剝離皮下瘤結節,微量電子稱進行瘤體稱重,實驗組平均瘤體質量為(0.094 2±0.044 5)g,陰性對照組平均瘤體質量為(0.694 0±0.149 5)g,實驗組瘤體質量明顯小于陰性對照組,差異有統計學意義(P<0.01,圖5)。此結果表明沉默GTPBP4基因能顯著抑制HCT116細胞裸鼠皮下成瘤能力。

圖5 兩組細胞裸鼠皮下移植瘤對比:A.陰性對照組及實驗組移植瘤剝離前對比;B.兩組移植瘤剝離后對比
2.5 Western blot法檢測細胞及裸鼠瘤體組織中蛋白的表達Western blot法檢測陰性對照組及實驗組穩定轉染細胞株及裸鼠移植瘤瘤體各蛋白表達情況,實驗組細胞及瘤體組織中的GTPBP4蛋白表達均明顯降低,說明在裸鼠成瘤過程中HCT116細胞穩定低表達GTPBP4基因,在此基礎上細胞及瘤體組織中c-Jun、Cyclin D1、BCL-2蛋白表達降低,Bax蛋白表達升高(圖6)。

圖6 Western blot法檢測兩組細胞及瘤體組織中GTPBP4、c-Jun、Cyclin D1、BCL-2和Bax蛋白表達水平:A.細胞及組織各蛋白條帶圖;B.各蛋白灰度值分析柱形圖,**P<0.01
2.6 免疫表型瘤體組織免疫組化結果顯示,GTPBP4、c-Jun、Ki-67及Cyclin D1表達均定位于細胞核,BCL-2、Bax表達定位于細胞膜及細胞質(圖7)。陰性對照組GTPBP4、c-Jun、BCL-2、Ki-67及Cyclin D1染色呈棕黃色顆粒,且分布廣泛,呈彌漫強陽性;實驗組瘤體GTPBP4、c-Jun、BCL-2、Ki-67及Cyclin D1染色均較淺,呈散在弱陽性,表達明顯較陰性對照組低;而Bax表達在陰性對照組中較實驗組弱。免疫組化染色結果與Western blot檢測結果一致。Ki-67主要反映腫瘤細胞的增殖活性,Ki-67在陰性對照組中的表達較強,間接表明陰性對照組細胞的增殖能力較強。

圖7 瘤體組織中各蛋白的表達:GTPBP4、c-Jun表達定位于細胞核,BCL-2、Bax表達定位于細胞膜及細胞質,EnVision法
3 討論
從GTPBP4基因在腫瘤中的研究現狀來看,Liu等[9]發現敲低GTPBP4基因在肝癌中的表達,能降低肝癌細胞增殖能力及裸鼠成瘤能力,這與本課題在結腸癌細胞株中的研究結果相符,進一步通過對基因芯片和通路富集分析,有學者認為ERBB信號通路可能是該基因重點作用的通路之一。在胃癌中的研究表明,GTPBP4基因在癌組織中表達高于癌旁組織,GTPBP4的穩定敲除抑制了細胞的增殖,促進細胞凋亡;從機制上講,GTPBP4與p53的相互作用使p53及其相關信號通路在GTPBP4穩定敲減情況下被激活[10]。此外,高表達GTPBP4的結腸癌患者預后較差,GTPBP4通過抑制RhoA的活性,特異性地誘導絲狀肌動蛋白重排,促進晚期結腸癌的侵襲、轉移[11]。
本組實驗結果發現,沉默GTPBP4基因后,結腸癌HCT116細胞增殖能力明顯受到抑制,體內外成瘤能力明顯減弱。腫瘤Ki-67增殖指數在裸鼠移植瘤中的表達也明顯降低。進一步研究發現GTPBP4表達下調后,原癌基因c-Jun在結腸癌細胞株中的蛋白水平表達隨之下調,并且在裸鼠移植瘤中檢測到的c-Jun蛋白表達也有所降低。由此推測GTPBP4基因與c-Jun基因在結腸癌的發生、進展過程中存在某種聯系。
AP-1是由Fos、Atf、Jun和MAF家族亞單位蛋白組成的二聚體轉錄因子,作為致癌轉化的介質因子[12],AP-1參與包括炎癥、分化、細胞遷移、腫瘤轉移、血管生成和傷口愈合在內的一系列生物學過程[13]。Jun家族蛋白成員包括c-Jun、Jun B和Jun D,c-Jun由核內原癌基因c-Jun編碼,具有正向調節細胞增殖的作用[14]。實驗中GTPBP4表達下調后,c-Jun蛋白及Cyclin D1的表達同時下調,可能是由于c-Jun作為上游調控基因,通過抑制p53腫瘤抑制劑和促進細胞Cyclin D1表達調節增殖進程[15]。這在一定程度上可以解釋GTPBP4下調后引起Cyclin D1表達降低的原因。
在調控凋亡發生方面,有研究報道BCL-2與c-Jun、c-Fos在胃癌中的表達呈正相關,并且晚期癌癥中的表達水平更高,三者共同作用介導胃癌的發展[16]。c-Jun是c-Jun N末端激酶(JNK)的下游靶效應分子,β-半乳糖苷結合蛋白(gal-1)通過JNK/c-Jun/AP-1途徑對T細胞死亡起一定的調控作用[17]。c-Jun N末端激酶JNK1/2/3-、p38-和ERK1/2-MAPK/c-Jun級聯信號通路可能有助于上調胃癌細胞中BARF1誘導的抗凋亡蛋白BCL-2和BCL-xL的表達,這有利于胃癌的發展[18]。Schwarz等[19]的研究中發現抗凋亡蛋白BCL-2 ERK途徑以時間和濃度依賴性方式導致c-Jun在SER73位點磷酸化,以促進更好發揮抑制細胞凋亡的作用。
總之,本實驗探討了下調GTPBP4基因的表達對結腸癌細胞體內及體外細胞增殖能力的影響。結果表明GTPBP4基因在結腸癌中主要是發揮癌基因的功能,促進結腸癌細胞增殖與發展。可能是通過調控c-Jun、Cyclin D1、BCL-2/Bax的表達,發揮促進腫瘤細胞增殖的作用。本實驗證實了GTPBP4基因在結腸癌發生、發展中的作用,為該基因成為分子治療靶標提供了實驗依據。
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
西南國防醫藥(2016年7期)2016-12-01 06:01:15
海峽科技與產業(2016年3期)2016-05-17 04:32:12
中國衛生標準管理(2015年1期)2016-01-14 03:41:26
河南醫學研究(2014年3期)2014-02-27 14:51:48
沈陽醫學院學報(2014年1期)2014-02-16 06:19:24
