托芬那酸通過下調LOXL2表達抑制人胃癌SGC-7901細胞侵襲①
2020-12-24 08:32:16徐松濤饒淑梅馬永超
中國免疫學雜志 2020年21期
程 琦 徐松濤 饒淑梅 馬永超
(漯河醫學高等專科學校,漯河 462002)
胃癌是威脅人類生命健康的主要惡性腫瘤之一。全世界每年死于胃癌的患者約78.3萬例,死亡率僅次于肺癌,位居惡性腫瘤譜第二位[1]。我國每年新增胃癌患者約67.9萬例,死亡49.8萬例,成為我國癌癥死亡的第二大原因[2]。90%以上癌癥患者的死亡是由腫瘤細胞侵襲和遠處轉移導致的[3]。因此侵襲和轉移是影響胃癌治療效果和生存預后的重要制約因素。托芬那酸(tolfenamic acid,TA)是一種廣泛應用的非甾體抗炎藥,具有抗炎、鎮痛、解熱等作用[4]。近年來,TA對癌癥的化學預防作用已獲大量臨床和流行病學證據支持[5]。多項研究證實,TA可通過抑制轉錄因子特異性蛋白1 (specificity protein 1,SP1)表達發揮抗腫瘤作用,包括神經母細胞瘤、結腸癌、卵巢癌、尤文肉瘤和胰腺癌等[6-10]。賴氨酰氧化酶樣蛋白2(lysyl oxidase-like 2,LOXL2)是受SP1調控的下游分子,在胃癌的侵襲和轉移過程中發揮重要作用[11-13]。但目前關于TA對胃癌細胞遷移和侵襲的作用鮮有報道。本研究擬通過分子生物學方法觀察TA對人胃癌SGC-7901細胞遷移和侵襲的影響,并探究其可能的生物學機制。
1 材料與方法
1.1材料 人胃癌SGC-7901細胞購自中國科學院上海生命科學研究院細胞資源中心;胎牛血清(FBS)和RPMI1640培養基購自Hyclone公司;TA購自Selleck公司;CCK-8試劑盒購自武漢博士德生物公司;結晶紫染色液、TRIzol試劑、RIPA裂解液和ECL化學發光試劑盒購自上海碧云天生物技術有限公司;Matrigel基質膠和Transwell小室購自Corning公司;靶向LOXL2的shRNA干擾質粒(pRS-LOXL2)和無義對照質粒(pRS-scrambled)購自美國Origene公司;脂質體(Lipofectamine?2000)購自Invitrogen公司;逆轉錄-聚合酶鏈式反應(RT-PCR)試劑盒購自大連寶生物公司;兔抗人SP1、LOXL2、Snail、E-鈣黏蛋白(E-cadherin,E-cad)、基質金屬蛋白酶9(matrix metalloproteinase 9,MMP9)、GAPDH單克隆抗體和辣根過氧化物酶標記山羊抗兔IgG購自Abcam公司;LOXL2、GAPDH引物由大連寶生物公司合成。
1.2方法
1.2.1細胞培養 SGC-7901細胞用含10% FBS的RPMI1640培養液置于37℃、5% CO2的細胞培養箱中培養;每2 d換液1次,待細胞覆蓋率達到90%時按1∶3比例傳代1次。
1.2.2藥物配制 將TA溶于二甲基亞砜(DMSO)制成100 mmol/L的儲備液。實驗前用完全培養液稀釋至所需濃度,其中DMSO的終濃度為0.1%,對照組為含0.1% DMSO的細胞培養液。
1.2.3CCK-8法檢測細胞活力 將SGC-7901細胞以5 000個/孔接種于96孔板,常規培養24 h;去上清,加入100 μl含不同劑量TA(0、3、6、12、24、48、96 μmol/L)的細胞培養液,每個劑量設5個復孔,常規培養24 h或48 h;每孔加入10 μl CCK-8溶液,37℃孵育1 h,用酶標儀檢測各孔450 nm處OD值;依據公式計算細胞活力,細胞活力(%)=(OD藥物組/OD對照組)×100%,采用SPSS18.0軟件的Probit回歸模型計算半數抑制濃度(IC50)。
1.2.4Transwell法檢測細胞侵襲 以不同劑量TA(0、3、6、12 μmol/L)處理SGC-7901細胞24 h,然后收集各組細胞,用無血清RPMI1640培養基制成密度為2×105個/ml的細胞懸液;用RPMI1640培養基按1∶5比例稀釋Matrigel基質膠,取50 μl加至上室底部,37℃孵育1 h;下室中加入600 μl含10% FBS的RPMI1640培養基,上室中加入200 μl細胞懸液,在37℃、5 % CO2條件下培養24 h;用棉簽擦去上室內的細胞,甲醇固定30 min,0.1%結晶紫染色20 min;顯微鏡下計數上室下表面附著的侵襲細胞。
1.2.5質粒轉染 用Lipofectamine?2000將質粒pRS-LOXL2和pRS-scrambled分別轉染SGC-7901細胞,細胞轉染按說明書進行;轉染pRS-LOXL2質粒的SGC-7901細胞命名為shLOXL2組,轉染pRS-scrambled質粒的SGC-7901細胞命名為shControl組;轉染48 h后,收集各組細胞進行后續實驗。
1.2.6RT-PCR檢測mRNA水平 用TRIzol試劑提取總RNA,取2 μg總RNA進行反轉錄,反轉錄條件:42℃ 60 min,70℃ 15 min。取2 μl進行PCR反應,反應條件為:94℃ 1 min,94℃ 30 s,60℃ 30 s,72℃ 1 min,32個循環。引物采用Primer premier 5.0軟件設計,經Blast驗證。LOXL2上游引物:5′-CTGCAAGTTCAATGCCGAGT-3′,下游引物:5′-AGTTTTGGCCACACACCATC-3′;GAPDH上游引物:5′-AATTCCATGGCACCGTCAAG-3′,下游引物:5′-GGGCAGAGATGATGACCCTT-3′。PCR產物經1%瓊脂糖凝膠電泳,紫外燈下拍照。
1.2.7Western blot法檢測蛋白水平 收集各組細胞,加入RIPA裂解細胞提取總蛋白,采用紫外分光光度法測定蛋白質濃度;用5×上樣緩沖液稀釋蛋白樣本,沸水浴10 min,SDS-PAGE電泳并轉移至PVDF膜,用5%脫脂奶粉室溫封閉1 h;加入一抗LOXL2(1∶1 000稀釋)、Snail(1∶1 000稀釋)、E-cad(1∶1 000稀釋)、MMP9(1∶1 000稀釋)和GAPDH(1∶2 000稀釋),4℃孵育過夜,TBST洗膜2次;加入二抗(1∶5 000稀釋)室溫孵育1 h,TBST洗膜2次;加入ECL工作液進行發光反應,用化學發光成像系統分析儀拍照。

2 結果
2.1TA對SGC-7901細胞活力的影響 如圖1所示,經不同劑量TA作用24 h或48 h后,與對照組相比,24、48和96 μmol/L TA組細胞活力均顯著降低(P<0.05),且隨藥物劑量的增加呈下降趨勢;3、6和12 μmol/L TA組細胞活力與對照組相比差異無統計學意義(P>0.05)。24 h和48 h的IC50分別為78.06 μmol/L和50.72 μmol/L。
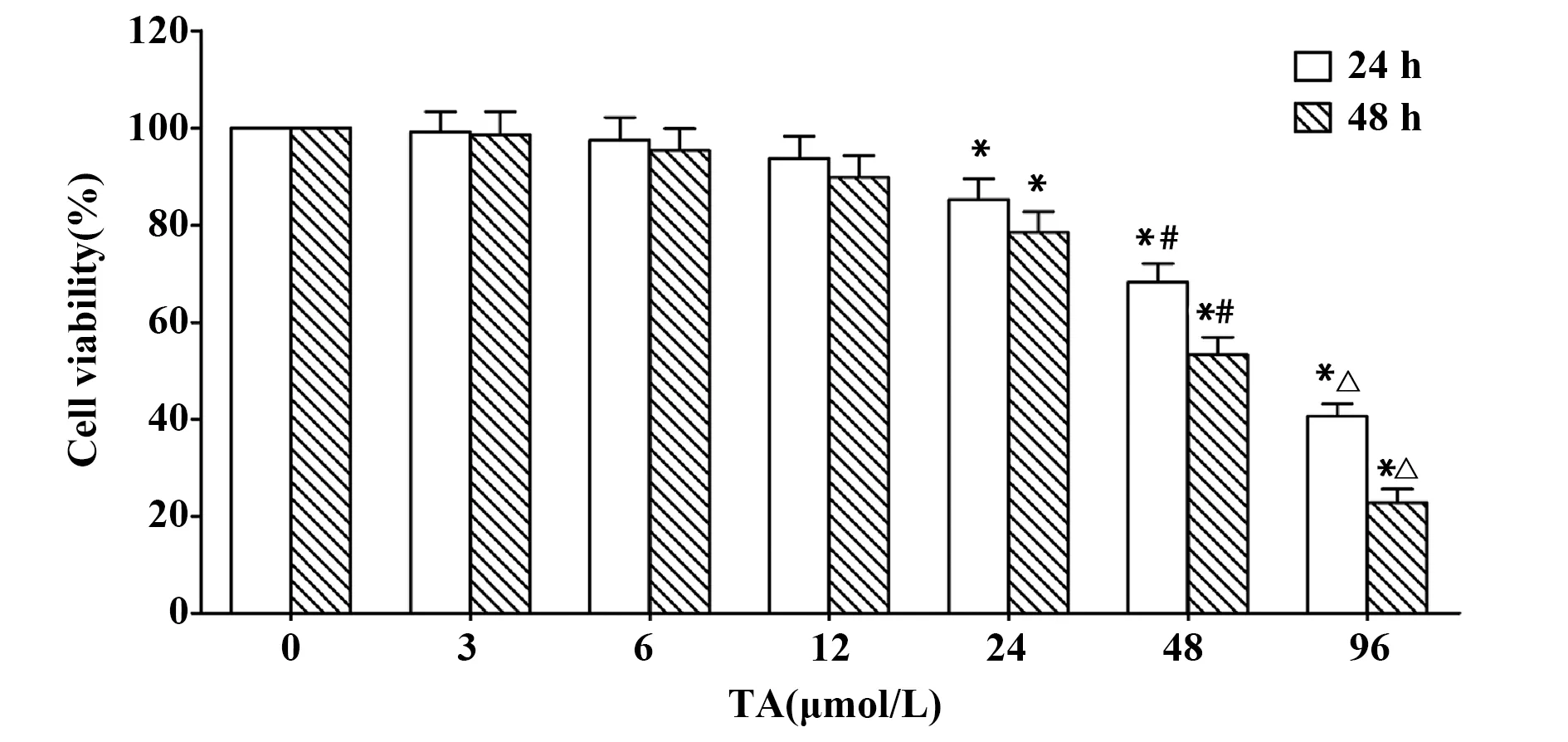
圖1 TA對SGC-7901細胞活力的抑制作用Fig.1 Inhibiting effect of TA on cell viability of SGC-7901 cellsNote:*.P<0.05 vs control group;#.P<0.05 vs 24 μmol/L TA group;△.P<0.05 vs 48 μmol/L TA group.
2.2TA對SGC-7901細胞侵襲的影響 如圖2所示,與對照組相比,3、6和12 μmol/L TA組侵襲細胞數均顯著減少(P<0.05),且隨著藥物劑量的增加而減少(P<0.05)。
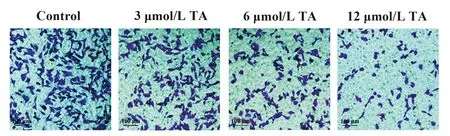
圖2 TA抑制SGC-7901細胞侵襲Fig.2 TA suppressed invasion of SGC-7901 cellsNote:*.P<0.05 vs control group;#.P<0.05 vs 3 μmol/L TA group;△.P<0.05 vs 6 μmol/L TA group.
2.3TA對侵襲相關蛋白水平的影響 如圖3所示,與對照組相比,經不同劑量TA(3、6、12 μmol/L)處理24 h后,SGC-7901細胞中Snail、MMP9、SP1和LOXL2蛋白水平均顯著下降(P<0.05),且隨TA劑量的增加呈下降趨勢(P<0.05);E-cad蛋白水平則隨TA劑量的增加逐漸升高(P<0.05)。
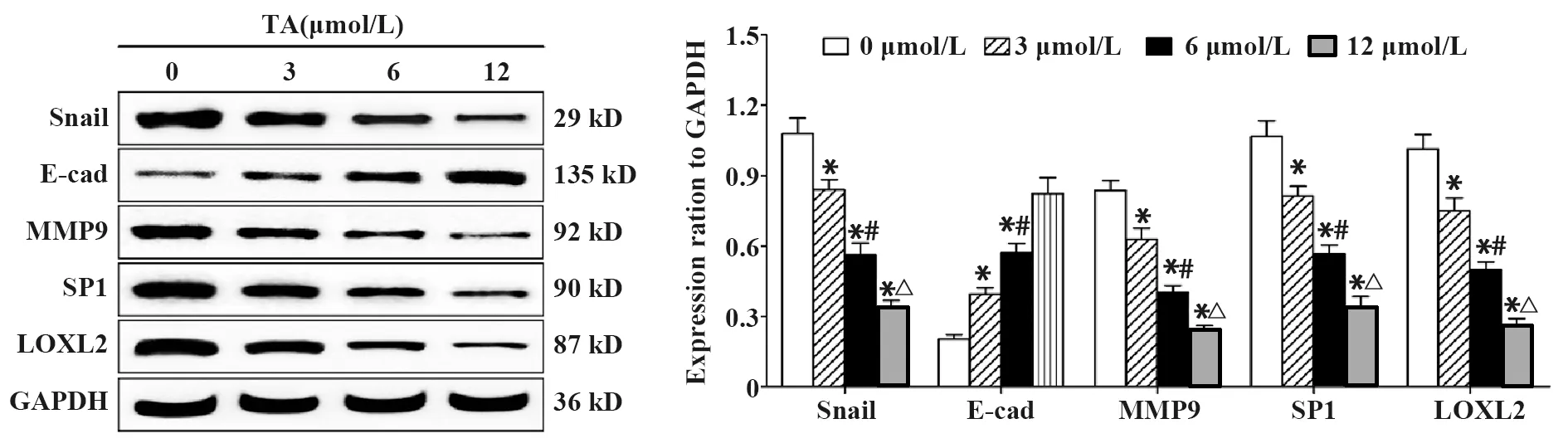
圖3 TA對SGC-7901細胞中Snail、E-cad、MMP9、SP1和LOXL2蛋白水平的影響Fig.3 Effect of TA on protein levels of Snail,E-cad,MMP9,SP1 and LOXL2 in SGC-7901 cellsNote:*.P<0.05 vs control group;#.P<0.05 vs 3 μmol/L TA group;△.P<0.05 vs 6 μmol/L TA group.
2.4TA對LOXL2的mRNA水平的影響 如圖4所示,與對照組相比,3、6和12 μmol/L TA組LOXL2的mRNA水平均顯著下降(P<0.05),且隨TA劑量的增加呈下降趨勢(P<0.05)。
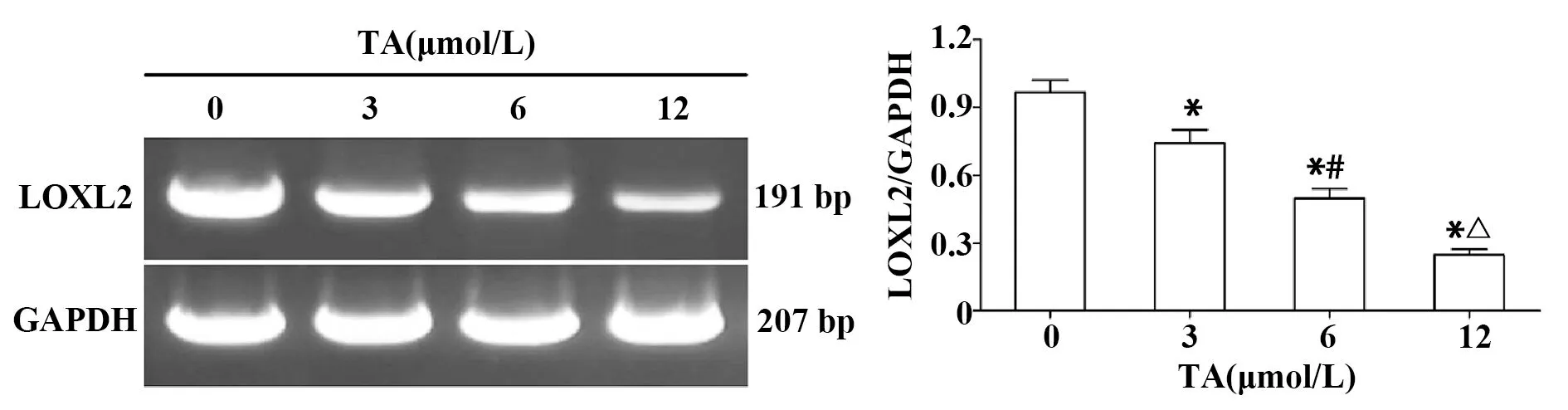
圖4 TA劑量依賴性下調LOXL2的mRNA水平Fig.4 TA down-regulated mRNA level of LOXL2 in a dose-dependent mannerNote:*.P<0.05 vs control group;#.P<0.05 vs 3 μmol/L TA group;△.P<0.05 vs 6 μmol/L TA group.
2.5敲除LOXL2基因對侵襲相關蛋白水平的影響 如圖5所示,與shControl組相比,shLOXL2組LOXL2、Snail和MMP9蛋白水平均顯著下降(P<0.05),E-cad蛋白水平顯著升高(P<0.05)。
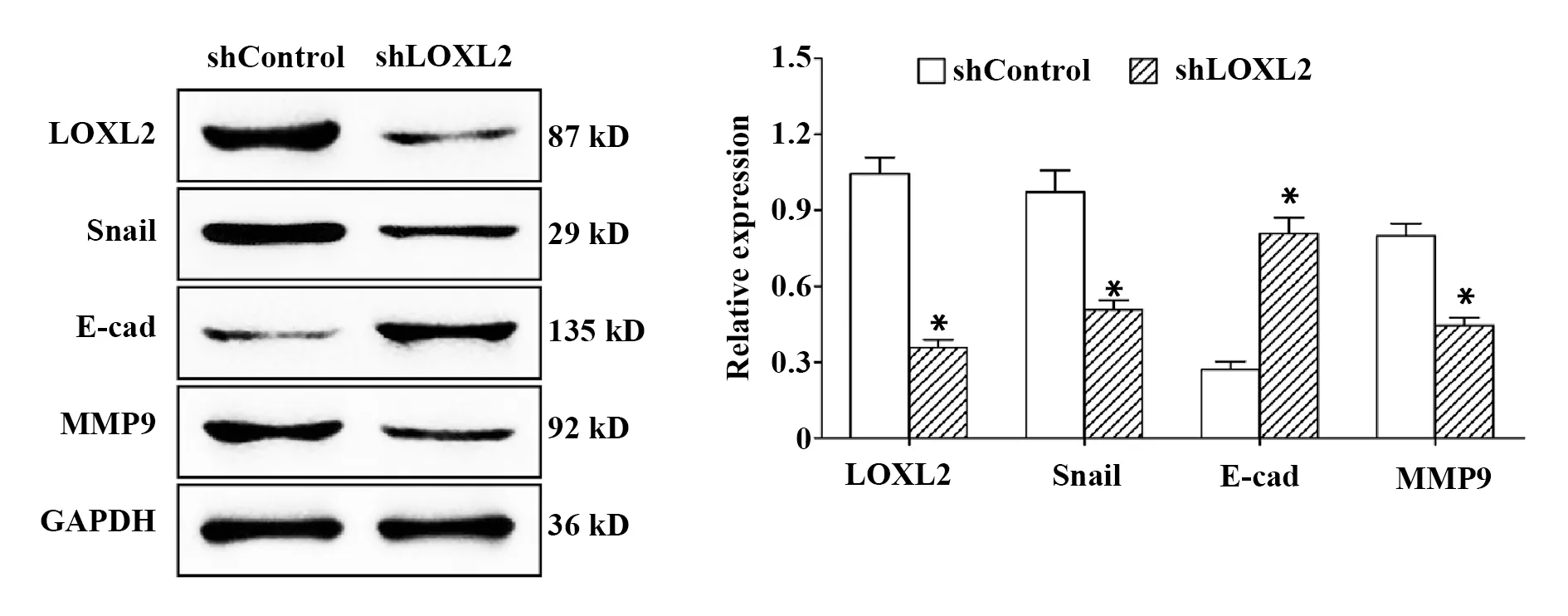
圖5 敲除LOXL2基因對LOXL2、Snail、E-cad和MMP9蛋白水平的影響Fig.5 Effect of LOXL2 gene knockout on protein levels of LOXL2,Snail,E-cad and MMP9Note:*.P<0.05 vs shControl group.
2.6敲除LOXL2基因對SGC-7901細胞侵襲的影響 如圖6所示,與shControl組相比,shLOXL2組侵襲細胞數明顯減少(P<0.05)。
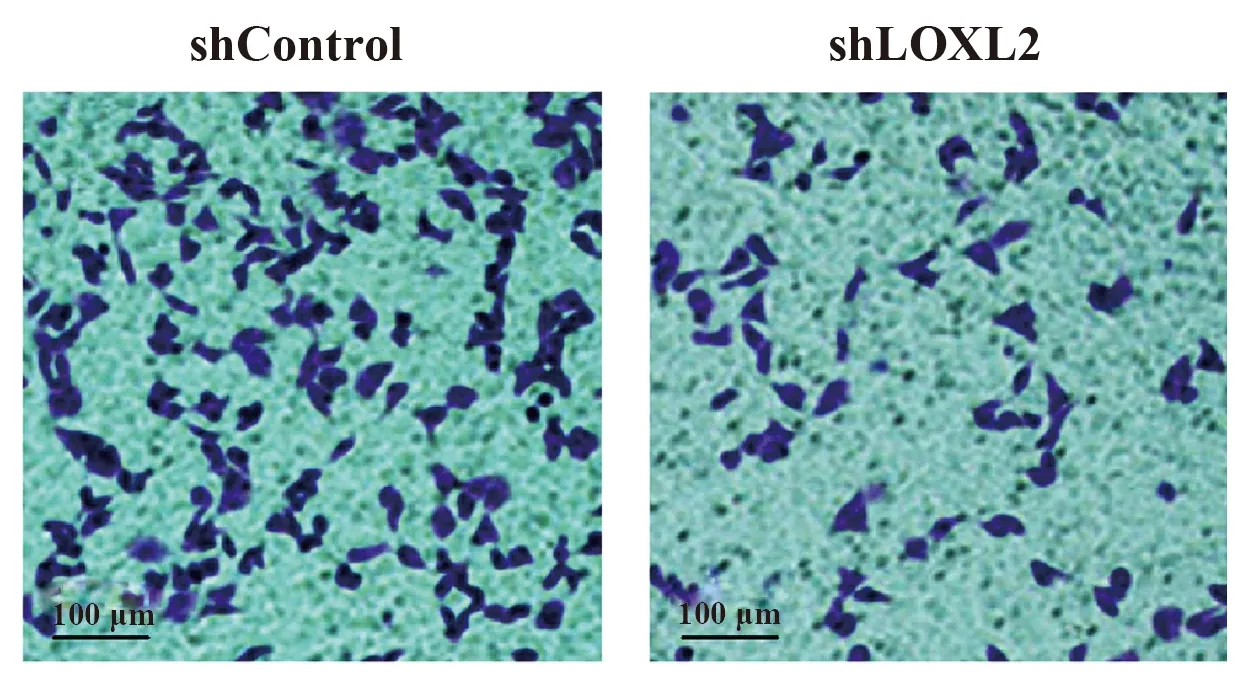
圖6 敲除LOXL2基因抑制SGC-7901細胞侵襲Fig.6 Knockout of LOXL2 gene inhibited invasion of SGC-7901 cellsNote:*.P<0.05 vs shControl group.
3 討論
有研究證實,TA可通過下調Slug蛋白表達抑制人鼻咽癌細胞遷移和侵襲[14]。本研究顯示,高劑量TA(>12 μmol/L)對人胃癌SGC-7901細胞活力具有顯著抑制作用,并呈現劑量和時間依賴性。為避免細胞活力受損對細胞侵襲能力的影響,本實驗采用低劑量TA(3、6、12 μmol/L)處理SGC-7901細胞24 h,然后行Transwell實驗檢測細胞侵襲。結果顯示,TA可劑量依賴性地抑制SGC-7901細胞侵襲,這可能與其下調SP1、LOXL2、Snail和MMP9蛋白水平,上調E-cad蛋白水平有關。
多項研究表明,LOXL2在惡性腫瘤細胞的侵襲和轉移過程中發揮重要作用,包括胰腺癌、乳腺癌、腎癌和肝癌等[15-18]。Kasashima等[13]研究表明,胃癌病理組織中LOXL2蛋白水平與局部侵襲范圍、淋巴結轉移、肝轉移和腹膜轉移呈正相關,敲除LOXL2基因可顯著抑制人胃癌OCUM-12細胞和NUGC細胞侵襲。動物實驗也證實,抑制LOXL2可阻止裸鼠胃癌移植瘤向肺轉移[12]。本研究顯示,TA可劑量依賴性地下調SGC-7901細胞中LOXL2的mRNA水平,敲除LOXL2基因可抑制SGC-7901細胞侵襲并下調Snail和MMP9蛋白水平并上調E-cad蛋白水平。
E-cad是一種鈣依賴性跨膜糖蛋白,其胞內段通過連環素錨定在細胞骨架上,使相鄰細胞形成穩定連接,其表達下調或缺失是腫瘤細胞發生上皮間充質轉化(epithelial mesenchymal transition,EMT)的重要標志之一。研究表明,E-cad蛋白表達上升會導致癌細胞侵襲能力減弱[19]。敲除LOXL2基因可上調腎癌細胞中E-cad蛋白表達從而抑制其遷移和侵襲[17]。Snail是一種具有鋅指結構的轉錄因子,可直接結合E-cad基因啟動子的E-box作用元件,從而抑制其表達[20]。LOXL2高表達可通過上調Snail蛋白表達促進肝癌細胞侵襲和轉移[18]。Zhu等[21]研究證實,下調Snail蛋白和上調E-cad蛋白會抑制人胃癌SGC-7901細胞侵襲。本研究表明,TA可能通過下調LOXL2/Snail通路促進E-cad蛋白表達,這可能是TA抑制SGC-7901細胞侵襲的機制之一。
Hong等[17]研究證實,敲除LOXL2基因可抑制腎癌細胞中MMP9蛋白表達。MMP9屬基質金屬蛋白酶超家族成員,可高效降解Ⅳ型膠原蛋白。Ⅳ型膠原蛋白是構成細胞外基質和基底膜的主要成分,而細胞外基質和基底膜是防御惡性腫瘤細胞向周圍組織侵襲的天然屏障。惡性腫瘤細胞可通過分泌MMP9破壞細胞外基質和基底膜的完整性,侵犯周圍組織、進而侵入血管和淋巴管向遠處轉移。Wei等[22]研究顯示,上調MMP9蛋白表達可增強SGC-7901細胞的侵襲能力。本研究表明,TA可劑量依賴性地抑制MMP9蛋白表達,這可能與TA下調LOXL2表達有關。
綜上所述,TA可在體外抑制人胃癌SGC-7901細胞侵襲,這可能與其抑制SP1和LOXL2表達,進而下調Snail和MMP9蛋白水平,上調E-cad蛋白水平有關。本研究為豐富TA的抗腫瘤侵襲作用提供新的實驗依據。
猜你喜歡
課堂內外·初中版(科學少年)(2023年10期)2023-12-10 00:43:06
全科護理(2022年10期)2022-12-26 21:19:15
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
國際放射醫學核醫學雜志(2021年10期)2021-02-28 08:41:58
人大建設(2019年12期)2019-05-21 02:55:32
中國衛生標準管理(2015年3期)2016-01-14 03:41:46
醫學研究雜志(2015年9期)2015-07-01 17:28:27
中國當代醫藥(2015年20期)2015-03-01 02:04:29
中國合理用藥探索(2014年11期)2014-03-11 20:30:20
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:52
