3 株發酵香腸源乳酸菌體外功能特性的比較
2020-12-29 03:01:48馮美琴欒曉旭
食品科學 2020年24期
關鍵詞:能力
馮美琴,欒曉旭,孫 健,
(1.金陵科技學院動物科學與技術學院,江蘇 南京 210038;2.南京農業大學食品科技學院,國家肉品質量安全控制工程技術研究中心,江蘇 南京 210095)
脂肪的過度氧化是發酵香腸加工、貯存過程中面臨的主要質量問題之一,常常造成產品風味、色澤和質地劣變而使發酵香腸的品質降低、貯期縮短[1-2],因此控制脂肪氧化程度對于發酵香腸的生產至關重要。添加抗氧化劑是降低產品氧化變質的有效手段。量大價低、抗氧化效果好的合成抗氧化劑如丁基羥基茴香醚、二丁基羥基甲苯和特丁基對苯二酚等常被添加在產品中用來抑制脂肪的氧化[3]。然而,研究發現合成抗氧化劑存在一定的安全性和毒性問題,所以安全、無毒的天然抗氧化劑的開發應用引起了研究者們的廣泛關注[4-6]。VE、VC、多酚類化合物、竹葉提取物及迷迭香提取物等被發現能夠有效地抑制脂肪的氧化。但這些天然抗氧化劑因成本過高或影響食品的感官,在實際使用上存在著局限性。
研究發現,在發酵肉制品中添加具有抗氧化潛能的發酵劑是減緩脂肪氧化的有效方法[7]。乳酸菌是香腸發酵成熟過程中重要的發酵劑之一。乳酸菌通過發酵不僅可以快速產酸、抑制腐敗菌和致病菌的生長繁殖,還能夠賦予產品獨特的風味、提高產品的營養價值,保證產品的品質與安全性[8]。大量研究表明,許多乳酸菌都具有抗氧化活性,但是不同菌株之間的抗氧化能力存在差異性,因此篩選具有高抗氧化活性的乳酸菌作為天然抗氧化劑提高發酵肉制品的抗氧化功能具有重要的實踐意義[9]。乳酸菌在自然界廣泛存在,而來源于傳統發酵肉制品的乳酸菌更利于用作肉制品發酵劑。李默等[10]從金華火腿等5 種發酵肉制品中成功篩選到了1 株具有較高抗氧化活性的肉制品發酵劑希臘魏斯氏菌L23。郭慧芬等[11]通過體外體內抗氧化實驗從6 種傳統發酵肉類產品中分離篩選出1 株抗氧化活性較強的菌株X31,最終鑒定為戊糖片球菌。Chen Qian等[7]報道了分離自哈爾濱紅腸的戊糖片球菌R1具有較高的抗氧化活性、超氧化物歧化酶(superoxide dismutase,SOD)活性和谷胱甘肽過氧化物酶(glutathione peroxidase,GSH-Px)活性,用該菌株接種生產香腸可以顯著降低脂肪氧化程度。Chen Qian等[12]進一步研究發現,用該戊糖片球菌和彎曲乳桿菌、清酒乳桿菌、木糖葡萄球菌混菌發酵生產香腸可以延緩肌肉蛋白質氧化。相較于泡菜、酸奶等其他發酵食品,有關中國傳統發酵肉制品中抗氧化乳酸菌的篩選與應用的報道較少。
本課題組前期從12 種傳統發酵香腸中篩選到3 株符合肉品發酵劑要求的乳酸菌,即發酵乳桿菌GZ114、植物乳桿菌NJ107和KM119[13]。本研究擬以1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)自由基清除率、羥自由基清除能力、超氧陰離子自由基清除能力、Fe2+螯合能力、還原能力等體外抗氧化指標和SOD、GSH-Px和過氧化氫酶(catalase,CAT)等酶活力指標系統評價3 株乳酸菌發酵上清液、菌體細胞和破碎提取物的體外抗氧化活性,并通過對3 株菌的疏水能力、耐模擬胃腸液消化能力和耐膽鹽能力檢測初步評價其益生特性,以期為具有較高抗氧化活性的功能性肉制品發酵劑的開發與應用提供理論依據。
1 材料與方法
1.1 材料與試劑
1.1.1 菌株
植物乳桿菌(Lactobacillus plantarum KM119,NCBI編號為MG798686)、發酵乳桿菌(L. fermentum GZ114,NCBI編號為MG798687)、植物乳桿菌 (L. plantarum NJ107,NCBI編號為MG798688)均為本實驗室由傳統發酵香腸中分離所得。
1.1.2 試劑
胃蛋白酶、胰蛋白酶、牛膽鹽、8-苯胺-1-萘磺酸(分析純)、DPPH(分析純) 美國Sigma公司;菲啰嗪 上海阿拉丁生化科技股份有限公司;MSA固體培養基、MRS肉湯 青島海博生物科技有限公司;SOD試劑盒、CAT試劑盒、GSH-Px試劑盒 南京建成生物工程研究所;其他常規試劑均為分析純,購于國藥集團化學試劑有限公司。
1.2 儀器與設備
OptiMairTM垂直流超凈工作臺 新加坡藝思高科技有限公司;HVE-50自動高壓滅菌 日本Hirayama公司; ICP260生化培養箱 德國Memmert公司;自動影像分析菌落計數儀 法國Interscience公司;Spectral Max M2e多功能酶標儀 美國伯騰儀器有限公司;HH-42快速恒溫水浴箱 常州國華電器有限公司;SIM-F-124制冰機 日本Sanyo公司;WH-2微型渦旋混合儀 上海滬西分析儀廠有限公司;YP1201N電子天平 上海 精密科學儀器公司;PTF-A300型萬分之一電子天平 瑞士Precisa公司。
1.3 方法
1.3.1 菌種培養及樣品制備
菌種在MRS液體培養基中37 ℃靜置培養18 h,傳代3 次,取最后一次活化的菌液4 ℃、6 000×g離心10 min,分別收集上清液和菌體。上清液經0.22 μm的水系濾膜過濾,即為發酵上清液。菌體用超純水洗滌3 次后重懸,并將菌體數調整為109CFU/mL,分為2 份。1 份菌液作為菌體細胞,另一份經冰浴超聲破碎細胞(300 W,工作2 s,暫停2 s,共10 min),破碎液于4 ℃、10 000×g離心15 min,收集上清液,鏡檢無完整細胞,即為破碎提取物。
1.3.2 抗氧化活性測定
1.3.2.1 DPPH自由基清除能力測定
參考Maryam等[14]的方法,測定DPPH自由基清除活性。
1.3.2.2 羥自由基清除率測定
根據Chen Qian等[7]的方法,測定羥自由基清除率。
1.3.2.3 超氧陰離子自由基清除率測定
參考Das等[15]的方法,測定超氧陰離子自由基清除率。
1.3.2.4 還原力測定
根據李默等[10]的方法,測定還原力。
1.3.2.5 螯合Fe2+能力的測定
根據王剛等[16]的方法,測定Fe2+螯合率。
1.3.2.6 SOD、CAT、GSH-Px活力測定
采用試劑盒,按照說明書使用。
1.3.3 乳酸菌疏水能力測定
參考陳明等[17]的方法,活化好的菌種于MRS液體培養基中在37 ℃培養18 h,4 ℃、10 000×g離心1 min,用pH 6.2 0.1 mol/L的硝酸鉀溶液洗滌2 次,重懸菌液,在600 nm波長處測定溶液吸光度,記為Ac。將菌懸液與二甲苯混勻(3∶1,V/V),室溫孵育10 min,劇烈混勻2 min,室溫靜置分層,測定下層水相的吸光度,記為As,按式(1)計算乳酸菌的疏水能力:1.3.4 乳酸菌耐胃腸液能力測定

模擬胃腸液的配制參考陳明等[17]的方法,將活化好的菌液按10%接種到模擬胃液中,混勻后于37 ℃靜置培養3 h,分別測定第0小時和第3小時培養液的活菌數。培養結束后,將培養液與模擬腸液按1∶9混合均勻,于37 ℃靜置培養3 h,測定培養液的活菌數,存活率按式(2) 計算:

式中:Nc和Ns分別為培養前后的乳酸菌數/(CFU/mL)。
1.3.5 乳酸菌耐膽鹽能力測定
參考劉宏宇等[18]的方法,將活化好的菌種按2%接種到MRS液體培養基中,37 ℃培養18 h,4 ℃、6 000×g離心10 min后收集菌體,洗滌,重懸于含5 mg/mL牛膽鹽的MRS培養基中,于37 ℃培養4 h。分別在第0小時和第4小時進行活菌計數,存活率計算同1.3.4節。
1.4 數據分析
2 結果與分析
2.1 乳酸菌的抗氧化活性比較
2.1.1 DPPH自由基清除能力比較

圖1 乳酸菌對DPPH自由基的清除能力Fig. 1 DPPH radical scavenging ability of lactic acid bacteria
如圖1所示,3 株菌各組分均表現出一定的DPPH自由基清除效果,但清除能力差異較大。發酵上清液的清除率顯著高于其他2 個組分(P<0.05),其中GZ114的清除率最高,可達91.93%。菌體細胞組KM119的清除活性最強,相當于其發酵上清液活性的48%左右;GZ114的清除活性最弱,清除率僅為33.73%。而破碎提取物的清除活性較弱,活性最強的為NJ107,清除率分別比KM119和GZ114高2%和3%左右,但僅為其發酵上清液清除率的10%左右。李默等[10]研究發現,分離自發酵肉制品的30 株乳酸菌的菌體細胞和胞內提取物的DPPH自由基清除活性較弱,而發酵上清液的DPPH自由基清除活性可高達91.24%,與本實驗結果一致。發酵上清液的DPPH自由基清除能力可能與菌株分泌的胞外多糖等胞外代謝產物有關,菌體細胞的清除能力則與菌體表面的化學物質如表面蛋白或多糖有關[19]。結果表明,3 株乳酸菌清除DPPH自由基的活性物質主要存在于胞外代謝產物中和菌體表面,細胞內則較少。
2.1.2 羥自由基清除能力的比較
羥自由基是具有強氧化性的自由基,會引起生物分子的氧化損傷。羥自由基清除劑的加入可猝滅體系中的羥自由基,生成Fe2+,引起體系吸光度的升高[20]。由圖2可知,菌體細胞的羥自由基清除率均高于其他2 個組分,KM119清除活性最強,可達37.61%,與其他菌株間差異顯著(P<0.05)。發酵上清液組活性次之,NJ107清除率最高,約為其菌體細胞活性的73%。破碎提取物組活性最弱,清除率均低于10%,該結果與郭慧芬等[11]的報道類似。王曦等[20]也報道35 株乳酸菌僅菌體細胞表現出一定羥自由基清除能力,無細胞提取物和胞外分泌物的清除活性較弱甚至為零。羥自由基造成的氧化損傷主要源自Fe2+和Cu2+等過渡金屬離子存在時的芬頓反應,抗氧化劑可以通過螯合這些離子抑制羥自由基的產生。

圖2 乳酸菌對羥自由基的清除能力Fig. 2 Hydroxyl radical scavenging ability of lactic acid bacteria
2.1.3 超氧陰離子自由基清除能力的比較

圖3 乳酸菌對超氧陰離子自由基的清除能力Fig. 3 Supernatant cation radical scavenging ability of lactic acid bacteria
由圖3可知,菌體細胞的超氧陰離子自由基清除活性最強,且各菌株活性較為接近,清除率在94%左右。破碎提取物組中GZ114的清除活性最強(91.2%),顯著高于其他兩菌株(P<0.05),與其發酵上清液能力相當。發酵上清液組NJ107的清除活性最弱,清除率僅為52.73%,但仍高于文獻報道的清除率(36.38%)[21]。乳酸菌存在的抗氧化酶如SOD、GSH-Px和CAT及其他氧化酶類是清除超氧陰離子自由基的重要酶促防御系統[21]。3 株乳酸菌的各組分均具有良好的超氧陰離子自由基清除活性,說明乳酸菌的各個組分中均可能檢測到抗氧化酶活性。
2.1.4 Fe2+螯合能力的比較
過渡金屬離子會促進氧化損傷,而Fe2+是最典型的促氧化劑,若能被螯合則可以阻止脂質過氧化反應的啟動。由圖4可知,菌體細胞組Fe2+螯合活性最高,與其他2 個組分間差異顯著(P<0.05),其中NJ107的Fe2+螯合率最高(77.13%)。發酵上清液組中GZ114活性最強,但僅相當于其菌體細胞活性的37.45%,KM119活性最弱(16%)。破碎提取物組中最高的為NJ107,其螯合率分別比KM119和GZ114高3%和11%左右。以上結果表明,3 株乳酸菌均具有良好的Fe2+螯合活性,菌株及組分間的差異可能是由于不同菌株螯合Fe2+的活性物質的種類、含量和分布不同引起的。Kai等[22]研究發現發酵乳桿菌ME-3和干酪乳桿菌KCTC 3260的抗氧化效果是通過螯合金屬離子實現。由此可推斷3 株乳酸菌可通過螯合金屬離子提高抗氧化水平。
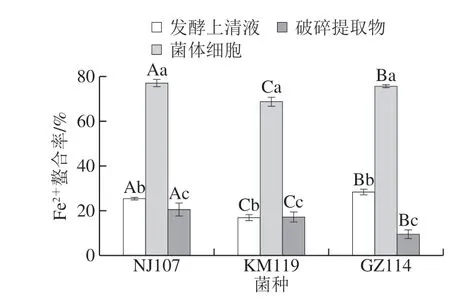
圖4 乳酸菌對Fe2+螯合能力Fig. 4 Fe2+ chelating ability of lactic acid bacteria
2.1.5 還原力的比較
還原力主要是指通過酶(CAT、NADH氧化酶和NADH過氧化物酶等)和非酶復合物(VC、VE和GSH等)減少氧自由基和金屬離子的能力[23-24],常作為評價抗氧化活性的重要指標。由圖5可知,發酵上清液組的還原能力最強,菌株間差異不大,GZ114最高;菌體細胞組其次,約為上清液的60%~65%;破碎提取物組的還原能力最低,其中最高的GZ114也僅相當于其發酵上清液的25.56%。Chen Qian等[7]認為細胞表面蛋白質或多糖可作為還原化合物以減少氧化應激,由此可以推測具有還原活性的胞外分泌物和菌體表面物質可能是3 株乳酸菌表現還原力的主要原因,而破碎提取物的還原能力則可能來自于細胞內的抗氧化成分如NADH、NADPH、谷胱甘肽、尿酸及各類抗氧化酶等。

圖5 乳酸菌的還原能力Fig. 5 Reducing power of lactic acid bacteria
2.1.6 酶活力比較
乳酸菌的酶促抗氧化防御系統被認為是抵御氧化損傷的一道重要屏障,其中抗氧化酶系統主要包括SOD、GSH-Px和CAT等[25]。本研究通過試劑盒測定法對3 株乳酸菌的SOD、GSH-Px和CAT抗氧化物酶水平進行了比較。
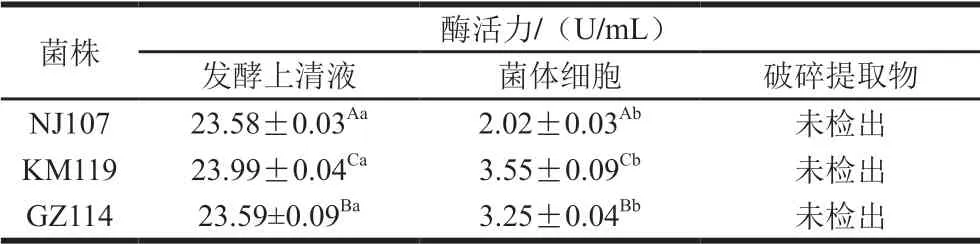
表1 乳酸菌的SOD活力Table 1 SOD activity of lactic acid bacteria
如表1所示,3 株菌發酵上清液中SOD活力為23 U/mL 左右,顯著高于菌體細胞(P<0.05),破碎提取物未檢出SOD活性。結果表明,3 株乳酸菌的SOD主要存在于胞外分泌物中,少部分附著于細胞表面。
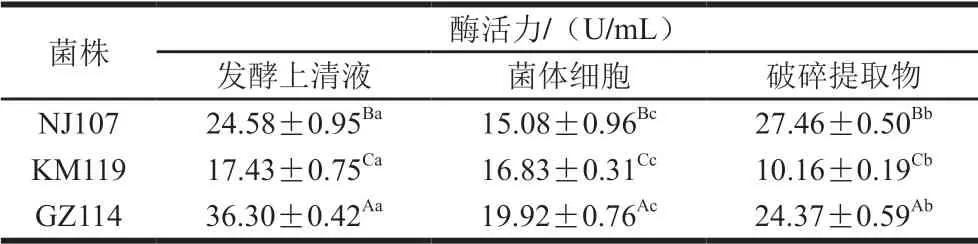
表2 乳酸菌的GSH-Px活力Table 2 GSH-Px activity of lactic acid bacteria
由表2可知,3 株菌各組分均有一定的GSH-Px活力,發酵上清液組的GSH-Px活力顯著高于菌體細胞和破碎提取物。GZ114發酵上清液酶活力最高(36.30 U/mL), KM119破碎提取物酶活力最低(10.16 U/mL)。NJ107破碎提取物的酶活力達27.46 U/mL,高于其發酵上清液(P<0.05);KM119菌體細胞的酶活力為16.83 U/mL, 顯著高于其破碎提取物(P<0.05)。以上結果表明,GSH-Px在不同乳酸菌菌株中的含量和分布不同。
由表3可知,菌株GZ114三組分的CAT活力較高,發酵上清液中活力最高(4.84 U/mL),與其余2 株菌差異顯著(P<0.05);NJ107次之,其中菌體細胞的活力最高;KM119活力最弱,且菌體細胞中未檢測出CAT。此外,CAT活力總體較弱,顯著低于同組分的SOD和 GSH-Px活力(P<0.05)。
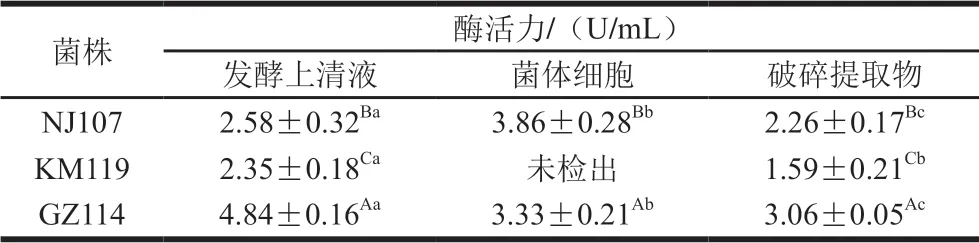
表3 乳酸菌的CAT活力Table 3 CAT activities of lactic acid bacteria
SOD具有清除超氧陰離子自由基的能力,GSH-Px和CAT則能夠清除過氧化物和羥自由基。結合圖2、3可知,3 株乳酸菌的羥自由基清除率與GSH-Px和CAT含量不呈正比,超氧陰離子自由基清除活性與SOD活力也不呈正相關,說明除了抗氧化酶類,3 株乳酸菌還含有其他可以清除羥自由基和超氧陰離子自由基的活性物質[26-28]。以上結果表明,3 株乳酸菌的抗氧化活性是抗氧化酶和其他非酶抗氧化物質共同作用的結果。
2.2 乳酸菌的疏水能力
能夠黏附于宿主細胞的表面是乳酸菌定植于腸道中發揮益生作用的前提,而菌體表面的疏水性與黏附能力有一定的相關性,因此菌體表面疏水性常常被用來評價乳酸菌的黏附性能[29]。如圖6所示,NJ107的疏水能力最強(13.45%),顯著高于其余2 株乳酸菌(P<0.05)。KM119的疏水能力次之,為8.85%,GZ114的疏水能力最差,僅為6.34%。結果表明,菌種不同疏水性有差異,即使菌種相同,在菌株水平上,疏水性也具有差異性,這一結論與占萌等[29]和Murphy[30]的研究一致。研究表明,細菌的表面疏水性與細菌的非特異性黏附密切相關[31-32],疏水性高的乳酸菌可與消化道上皮細胞間形成較強的相互作用力,有益于細菌附著。李清等[33]分析了10 株植物乳桿菌的表面疏水性,證實疏水性與細胞黏附能力呈正相關。由此推測NJ107可能具有較好的黏附能力,有助于其益生功能的發揮。
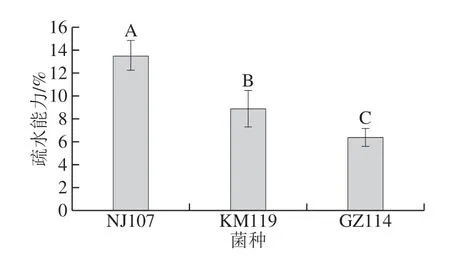
圖6 乳酸菌的疏水能力Fig. 6 Hydrophobic ability of lactic acid bacteria
2.3 乳酸菌耐胃腸道消化能力
乳酸菌只有具備良好的耐胃腸道消化能力,保持較高的存活率才能發揮其益生功能。如表4所示,3 株菌均具有良好的耐消化能力。其中,在模擬胃液(pH 3.0)中消化3 h后,3 株乳酸菌的存活率均在87%以上,NJ107存活率最高,為89.17%,與其他2 株菌差異不顯著 (P>0.05)。繼續在模擬腸液中消化3 h,NJ107仍可保持75.79%的存活率,GZ114次之,均顯著高于KM119的存活率(P<0.05)。Han Qi等[34]發現植物乳桿菌對人體消化液有良好的耐受能力,與本實驗結果類似。總體而言,經模擬胃腸液消化后菌株存活率高低依次為NJ107、GZ114、KM119,且3 株菌的存活率均在70%以上,說明3 株菌對模擬胃腸液都有較好的耐受性。
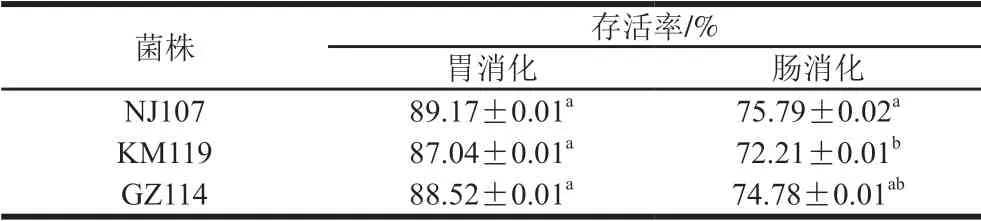
表4 乳酸菌在模擬胃腸液中消化后的存活率Table 4 Survival rates of lactic acid bacteria after digestion in simulated gastrointestinal
2.4 乳酸菌耐膽鹽能力
腸液的主要成分為胰液和膽汁,因此益生菌要在人體腸道內發揮作用,除要具有良好的耐胃腸道酶系消化能力外,還應具有一定的膽鹽耐受能力。膽鹽在腸道中質量濃度一般為0.3~3 mg/mL,本實驗設計了5 mg/mL的高膽鹽質量濃度比較3 株菌對膽鹽的耐受能力。由表5可知,在5 mg/mL的高膽鹽環境中培養4 h后,KM119的存活率高達91.45%,顯著高于其他2 株菌(P<0.05),NJ107和GZ114的耐膽鹽能力相對稍低,4 h后的存活率在76%左右。劉宏宇等[18]研究發現乳酸菌對5 mg/mL膽鹽的耐受能力較差,與本研究結果不同,這可能與菌種差異或菌株來源有關。結果表明,3 株菌均具有良好的耐膽鹽能力,具備作為功能性發酵劑開發利用的潛力。

表5 乳酸菌的膽鹽耐受能力Table 5 Bile salt tolerance capacities of lactic acid bacteria
3 結 論
本實驗比較了分離自傳統發酵香腸、具有良好發酵特性的3 株乳酸菌的抗氧化活性、表面疏水性、耐胃腸道消化能力和耐膽鹽能力。結果表明,3 株菌均具有良好的體外抗氧化活性,但不同的抗氧化指標測得的菌株間和各組分間的抗氧化活性差異較大,這可能與抗氧化反應機制及菌株中抗氧化活性物質的分布、種類、濃度不同有關。綜合而言,3 株菌的發酵上清液和菌體細胞的抗氧化活性明顯高于破碎提取物,為今后3 株乳酸菌作為天然抗氧化劑的應用提供了參考依據。菌株表面疏水性高低依次為NJ107>GZ114>KM119,3 株菌在模擬胃腸道酶系消化、0.5%高濃度膽鹽環境下均能保持較高的存活率。因此,3 株乳酸菌均可作為高抗氧化活性的功能性肉品發酵劑用于實際生產或益生性產品的進一步研究。
猜你喜歡
發明與創新(2022年30期)2022-10-03 08:40:56
中學生數理化·七年級數學人教版(2022年6期)2022-06-05 06:50:58
意林(兒童繪本)(2020年2期)2021-01-07 02:12:04
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
作文成功之路·小學版(2020年5期)2020-06-11 12:48:46
意林(兒童繪本)(2019年9期)2019-10-15 08:51:46
中國生殖健康(2019年10期)2019-01-07 01:21:14
人大建設(2018年6期)2018-08-16 07:23:10
新高考(英語進階)(2018年1期)2018-04-18 14:00:11
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
