云南省埃及伊蚊對擬除蟲菊酯類抗性群體的擊倒抗性基因突變分析
2021-01-06 03:06:14蘭學(xué)梅楊明東姜進(jìn)勇
中國人獸共患病學(xué)報 2020年12期
關(guān)鍵詞:基因突變
蘭學(xué)梅,楊明東,楊 銳,姜進(jìn)勇
埃及伊蚊(AedesAegypti)是新入侵云南的蚊蟲,2002年首先在瑞麗市姐告口岸被發(fā)現(xiàn)[1],其入侵、定殖和擴散能力強,目前已成為云南省景洪市、勐臘縣、瑞麗市和耿馬縣孟定鎮(zhèn)的優(yōu)勢蚊種[2],云南省2013年來多次登革熱疫情暴發(fā)地均有埃及伊蚊分布,是云南省登革熱的重要傳播媒介[3-5]。媒介控制是登革熱疫情處置的主要措施之一,瑞麗、景洪、勐臘和耿馬等地主要使用各種衛(wèi)生化學(xué)殺蟲劑進(jìn)行空間噴灑(超低容量噴霧和熱煙霧)結(jié)合滯留噴灑,來迅速殺滅成蚊從而控制登革熱疫情。各地疫情處置過程中,長期、大量使用擬除蟲菊酯類殺蟲劑進(jìn)行滅蚊控制,埃及伊蚊可能對菊酯類殺蟲劑產(chǎn)生了不同程度的抗藥性[6]。因此,2016年以來,本課題組先后測定了云南省瑞麗市、景洪市、勐臘縣、勐海縣和耿馬縣等登革熱重點地區(qū)的埃及伊蚊成蚊對幾種擬除蟲菊酯類殺蟲劑的敏感性,結(jié)果證實均存在不同程度抗藥性[7]。為進(jìn)一步研究其產(chǎn)生抗性的機制,本研究在以往研究的基礎(chǔ)上,收集上述5個種群的埃及伊蚊野生種群成蚊,在生物測定擬除蟲菊酯類殺蟲劑(氯菊酯和高效氯氟氰菊酯)敏感性的基礎(chǔ)上,檢測其對應(yīng)kdr基因突變情況,用于豐富我省埃及伊蚊群體kdr基因突變數(shù)據(jù),為探討分子檢測結(jié)果與生物測定結(jié)果之間的關(guān)聯(lián)性提供科學(xué)依據(jù)。
1 材料與方法
1.1供試蚊蟲來源 2016-2018年,在云南省景洪市、勐臘縣、勐海縣、瑞麗市和耿馬縣(東、南、西、北和中)5個片區(qū),采集埃及伊蚊的幼蟲和蛹(均采自疫點及其周圍室內(nèi)外花瓶、水桶、泡菜壇、飲水機水槽、輪胎及其他廢棄容器等)。羽化和鑒定[8]后,飼喂鼠血(豚鼠,產(chǎn)地:普洱),分類飼養(yǎng)繁殖至子一代(F1),進(jìn)行對殺蟲劑的生物學(xué)測定。隨即進(jìn)行埃及伊蚊成蚊對3.20%氯菊酯和0.08%高效氯氟氰菊酯的抗藥性測定(藥膜由中國疾病預(yù)防控制中心傳染病預(yù)防控制所提供),實驗結(jié)果均為抗性。24 h后死亡和存活樣本單管單只分別收集,并置于75%乙醇中保存。
1.2kdr基因擴增及電泳 收集的埃及伊蚊按照試劑盒(購自天根生化科技(北京)有限公司)說明書,提取單蚊基因組DNA。參照文獻(xiàn)[9]合成引物,擴增1 016和1 534兩個位點的突變情況。各引物的PCR反應(yīng)體系相同:包含2×Taq預(yù)混液12.5 μL,10 μmol/L,非特異性正反引物各1.0 μL,特異性內(nèi)引物1.0 μL,DNA模板2.0 μL,加雙蒸滅菌水至 25.0 μL。反應(yīng)條件為:94 ℃ 3 min;94 ℃ 30 s,60 ℃ 30 s,72℃ 1 min,35個循環(huán);72 ℃延伸7min。擴增產(chǎn)物經(jīng)1.5%瓊脂糖凝膠電泳檢測后,觀察電泳條帶大小。V1016G等位基因檢測中,M2-For和M2-Rev為非特異性外引物,用于大片段擴增(592 bp);特異性內(nèi)引物M2-V和M2-G擴增片段長度為348 bp(僅有V,敏感純合子;僅有G,抗性純合子;VG并存,抗性雜合子)。F1534C等位基因檢測中,M3-For和M3-Rev為非特異性外引物,用于大片段擴增(517 bp);特異性內(nèi)引物M3-F和M3-C擴增片段長度為284 bp(僅有F,敏感純合子;僅有C,抗性純合子;FC并存,抗性雜合子)。見圖1。

圖1 埃及伊蚊鈉離子通道擊倒抗性基因AS-PCR檢測示意圖
1.3數(shù)據(jù)分析 運用SPSS20.0統(tǒng)計軟件進(jìn)行統(tǒng)計分析,采用χ2檢驗分析kdr各基因突變型間的相關(guān)性,檢驗水準(zhǔn)為α=0.05。
2 結(jié) 果
采集自景洪市、勐臘縣、勐海縣、瑞麗市和耿馬縣5個地區(qū),共計580份雌性埃及伊蚊樣本被用于V1016G和F1534C擊倒抗性突變的檢測。5個重點地區(qū)埃及伊蚊均發(fā)現(xiàn)有V1016G和F1534C突變存在,其中,V1016G的突變率為99.83%(579/580),5個地區(qū)V1016G的突變率都較高,除了瑞麗市為99.26%外,其余突變率均為100%。F1534C的突變率為46.38%(269/580),5個地區(qū)F1534C的突變率分別為84.16%(85/101)、67.80%(80/118)、49.17%(59/120)、28.89%(39/135)和5.66%(6/106)。V1016G和F1534C突變率差異存在統(tǒng)計學(xué)意義(χ2=421.338,P=0.000),V1016G的突變率高于F1534C(表1)。
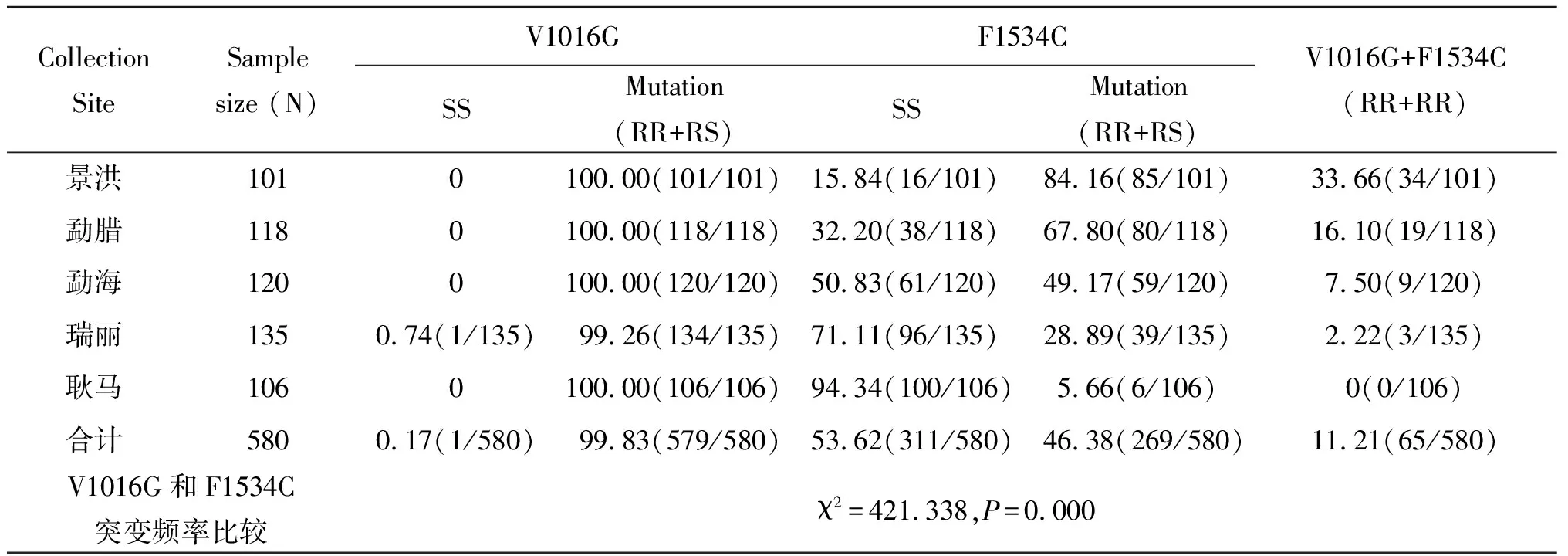
表1 云南省埃及伊蚊對菊酯類殺蟲劑基因突變率(%)測定
2.1不同殺蟲劑V1016G和F1534C突變的比較 埃及伊蚊對氯菊酯抗性的樣本檢測了273份,V1016G和F1534C突變率分別為100.00%(273/273)和50.92%(139/273)。對氯菊酯的V1016G和F1534C突變率進(jìn)行比較,差異存在統(tǒng)計學(xué)意義(χ2=177.583,P=0.000);埃及伊蚊對高效氯氟氰菊酯抗性的樣本檢測了307份,V1016G和F1534C突變率分別為99.67%(306/307)和47.62%(130/273)。對高效氯氟氰菊酯V1016G和F1534C突變率差異存在統(tǒng)計學(xué)意義(χ2=209.802,P=0.000)(見表2、3)。
對氯菊酯和高效氯氟氰菊酯的V1016G突變率進(jìn)行比較,差異無統(tǒng)計學(xué)意義(χ2=0.891,P=0.345);對氯菊酯和高效氯氟氰菊酯的F1534C突變率進(jìn)行比較,差異有統(tǒng)計學(xué)意義(χ2=4.268,P=0.039)(表2、3)。

表2 云南省埃及伊蚊擊倒抗性基因突變頻率測定
2.2不同地區(qū)V1016G和F1534C突變的比較 采用χ2檢驗,5個地區(qū)1 016位點的基因突變率進(jìn)行比較,差異無統(tǒng)計學(xué)意義(χ2=3.302,P=0.509);5個地區(qū)1 534位點的基因突變率進(jìn)行比較,差異有統(tǒng)計學(xué)意義(χ2=167.383,P=0.000)。進(jìn)一步進(jìn)行景洪市與其他4個地區(qū)1 534位點的基因突變率進(jìn)行兩兩比較,其中景洪和勐臘、勐海、瑞麗、耿馬比較的統(tǒng)計結(jié)果分別為:χ2=7.842,P=0.005;χ2=29.578,P=0.000;χ2=70.778,P=0.000;χ2=129.365,P=0.000。

表3 不同殺蟲劑V1016G和F1534C突變率比較
2.3抗性表型與敏感表型V1016G和F1534C突變的比較 5個地區(qū)埃及伊蚊野生群體對氯菊酯和高效氯氟氰菊酯進(jìn)行抗藥性測定后,實驗存活個體定義為抗性表型,實驗死亡個體定義為敏感表型。抗性表型樣本檢測510份,V1016G和F1534C突變頻率分別為100.00%(510/510)和49.02%(250/510);敏感表型樣本檢測70份,V1016G和F1534C突變頻率分別為98.57%(69/70)和72.86%(51/70)。抗性表型與敏感表型的V1016G突變率進(jìn)行比較,差異有統(tǒng)計學(xué)意義(χ2=7.298,P=0.007);抗性表型與敏感表型的F1534C突變率進(jìn)行比較,差異有統(tǒng)計學(xué)意義(χ2=14.010,P=0.000)。
2.4V1016G和F1534C位點同時純合子突變 580份樣本中,有65份樣本同時存在V1016G和F1534C位點純合子突變,突變率為11.21%;不同地區(qū)同時突變率范圍為0%~33.66%,景洪同時突變率最高為33.66%,瑞麗最低,本次實驗耿馬未發(fā)現(xiàn)同時存在純合子突變(表1)。
3 討 論
擊倒抗性(knockdown resistance,kdr)是引起蚊蟲對擬除蟲菊酯類殺蟲劑產(chǎn)生抗性的機制之一,埃及伊蚊電壓門控鈉離子通道(voltage-gated sodium channel, VGSC)上很多位點已被研究檢測出來,但僅有少數(shù)位點突變被證實與抗性相關(guān)[10-11],如埃及伊蚊不同抗性群體,VGSC基因突變的密碼子有3個(I1011M/V、V1016G/I和F1534C)與DDT和菊酯類殺蟲劑抗性相關(guān)。
本次研究景洪、勐臘、勐海、瑞麗和耿馬的埃及伊蚊存在V1016G和F1534C突變,V1016G的突變以抗性純合子(RR)突變?yōu)橹鳎c師燦南[12]2015年對景洪市城區(qū)埃及伊蚊的檢測結(jié)果一致,緬甸[13]和泰國[14]也發(fā)現(xiàn)相似的突變。然而,Sebastien Marcombe等2014年在老撾埃及伊蚊中發(fā)現(xiàn)存在V1016G和F1534C突變,以F1534C突變占主導(dǎo)地位,突變率高達(dá)96%,而V1016G存在低水平的突變,突變率僅為36%[15],Hitoshi Kawada等報告越南V1016G突變率更低,僅為0.26%[16]。存在不同的原因可能是入侵云南埃及伊蚊與老撾和越南不是同一來源,或是各國使用的殺蟲劑種類、時間和數(shù)量不一致,尚需進(jìn)一步分析和研究。通過兩種殺蟲劑V1016G突變率和5個不同抗性種群V1016G突變率的比較看出,差異均無統(tǒng)計學(xué)意義,進(jìn)一步證實V1016G突變廣泛存在于對擬除蟲菊酯類殺蟲劑產(chǎn)生抗藥性的埃及伊蚊中。Han-Hsuan Chung等研究發(fā)現(xiàn)臺灣地區(qū)的埃及伊蚊S989P和D1763Y突變均與V1016G并發(fā)。S989P和V1016G并存[17-18],以及D1763Y和V1016G同時并存[19]。至于云南的埃及伊蚊中是否存在上述現(xiàn)象,尚需進(jìn)一步研究。
5個地區(qū)埃及伊蚊的1 534位點均發(fā)生了基因突變,且不同地區(qū)埃及伊蚊的突變率存在明顯的差異,以景洪最高為84.16%,耿馬最低為5.66%。原因可能是每個地區(qū)殺蟲劑的種類、數(shù)量和使用年限不同所致,也可能與不同地區(qū)埃及伊蚊對擬除蟲菊酯類殺蟲劑的抗性程度不同有關(guān),需檢測不同地區(qū)埃及伊蚊對殺蟲劑的抗性程度類進(jìn)行證實。本研究中,抗性表型與敏感表型V1016G和F1534C突變率均存在差異,兩種位點的突變均以抗性表型的突變率較高,與其他研究者的結(jié)論一致[20-21]。證實V1016G和F1534C突變與埃及伊蚊對擬除蟲菊酯類殺蟲劑產(chǎn)生抗性具有相關(guān)性,是引起殺蟲劑抗性的機制之一。
多個位點突變同時存在的情況下會導(dǎo)致埃及伊蚊對擬除蟲菊酯類殺蟲劑產(chǎn)生更高水平的抗性[13],泰國埃及伊蚊中G1016和C1534 同時突變對溴氰菊酯抗性有相加作用。臺灣地區(qū)的埃及伊蚊中發(fā)現(xiàn)同時存在S989P/V1016G/F1534C或V1016G/F1534C/D1763Y的3個雜合突變的顯現(xiàn)[22],結(jié)果對殺蟲劑具有較高的抗性。云南埃及伊蚊1 016和1 534位點突變同時為抗性純合子(RR)的頻率為11.21%(65/580)遠(yuǎn)遠(yuǎn)高于老撾埃及伊蚊(4/1 076,0.37%)[17]。景洪市同時突變?yōu)榭剐噪s合子的頻率最高為33.66%。可能與不同地區(qū)埃及伊蚊對殺蟲劑產(chǎn)生的抗性程度不同有關(guān)。至于不同位點三重雜合突變的現(xiàn)象是否具有協(xié)同效應(yīng),有待研究。
我省登革熱重點地區(qū)埃及伊蚊野外群體存在V1016G和F1534C突變,V1016G突變廣泛存在,F(xiàn)1534C突變以景洪埃及伊蚊種群為最高,且兩種突變同時存在也是景洪的埃及伊蚊種群最高,從分子生物學(xué)的角度證實了景洪市廣泛使用和不受控制地頻繁使用光穩(wěn)定的擬除蟲菊酯可能是該突變的強大選擇壓力,埃及伊蚊對擬除蟲菊酯類殺蟲劑已經(jīng)產(chǎn)生了高度抗性,應(yīng)注意殺蟲劑的合理選用。我省登革熱高發(fā)地區(qū),應(yīng)定期進(jìn)行蚊蟲生物學(xué)測定、代謝酶活性和擊倒抗性基因突變檢測,從而能夠根據(jù)分子檢測結(jié)果預(yù)測蚊蟲的抗性水平,時時掌握抗性發(fā)生發(fā)展動態(tài),最終實現(xiàn)對蚊蟲種群抗性的“分子監(jiān)測”。
利益沖突:無
猜你喜歡
英語世界(2023年6期)2023-06-30 06:29:10
中國醫(yī)學(xué)影像學(xué)雜志(2021年6期)2021-08-13 08:43:36
中國生殖健康(2020年2期)2021-01-18 02:51:26
小學(xué)生導(dǎo)刊(2018年13期)2018-06-29 03:49:00
中國生殖健康(2018年2期)2018-01-12 13:57:51
現(xiàn)代檢驗醫(yī)學(xué)雜志(2016年4期)2016-11-15 02:01:14
中國現(xiàn)代醫(yī)學(xué)雜志(2015年26期)2015-12-23 11:04:22
鄭州大學(xué)學(xué)報(醫(yī)學(xué)版)(2015年2期)2015-02-27 14:50:44
中華皮膚科雜志(2014年4期)2014-12-19 12:55:49
中國神經(jīng)精神疾病雜志(2014年1期)2014-03-01 03:23:22
