丙泊酚對老年癡呆大鼠AMPK自噬通路及認知功能的影響
2021-01-27 07:59:34鄭仲磊張萬平蘇玉強范曉英
河北醫學 2021年1期
鄭仲磊,張萬平,蘇玉強,薛 沙,范曉英
(西安醫學院第二附屬醫院手麻科,陜西 西安 710038)
老年癡呆患者記憶力和認知能力的下降是由于突觸功能喪失,炎癥信號增強,氧化應激,大腦中的突觸損傷,老年性斑塊增加和神經原纖維纏結的逐步沉積以及神經元變性所致[1]。自噬是一種細胞內分解代謝機制,在病理條件下(如阿爾茨海默病)自噬對神經元具有保護作用。腺苷酸活化蛋白激酶(AMPK)是一種普遍存在的絲氨酸/蘇氨酸蛋白激酶[2,3]。AMPK的表達和激活在阿爾茨海默氏病患者的大腦中也增加,抑制AMPK的激活可通過減少β淀粉樣蛋白(Aβ)的產生和調節真核延伸因子2(eEF2)來改善海馬突觸可塑性的損害[4]。參與記憶功能的大腦區域主要依賴于含有N-甲基-D-天冬氨酸受體2B(NR2B)亞基。在產后發育過程中,NR2B表達穩定下降,NR2B的受體失活更慢,因此與突觸可塑性水平升高有關。小鼠中NR2B表達的下調改善了突觸可塑性和記憶形成,通過改變合成、轉運或降解而導致NR2B水平降低,其突觸可塑性和記憶力均得到改善[5]。丙泊酚是最常用的靜脈麻醉藥,已顯示出減弱含半胱氨酸的天冬氨酸蛋白水解酶(caspase)激活和凋亡而具有神經保護作用[6]。本研究擬探討丙泊酚對老年癡呆大鼠AMPK自噬通路及認知功能的影響。
1 材料與方法
1.1實驗動物及分組:清潔級SD(Sprague Dawley)大鼠100只,購自昆明醫科大學實驗動物中心,雌雄各半,96周齡,體重(520±40)g,動物生產許可證號:SCXK(云)2019-0006,動物使用許可證號:SYXK(云)2019-0008,動物質量合格證號:0019695849。飼養環境:溫度(21±1)℃,濕度(55±5)%。大鼠隨機分成5組:對照組、模型組、鹽酸米諾環素組(50mg/kg)、丙泊酚低劑量組(50mg/kg)、丙泊酚高劑量組(100mg/kg),每組20只。
1.2主要藥品、試劑與儀器:丙泊酚(原料藥,南京科維思生物科技股份有限公司,純度99.98%,批號HJ-1587);鹽酸米諾環素膠囊(惠氏制藥有限公司,規格100mg/粒,批號190513);β樣淀粉蛋白1-42(β-amyloid protein1-42,Aβ1-42)(美國sigma公司,批號L2873);吖啶橙染料、化學發光(ECL)Western印跡檢測試劑盒(碧云天生物技術研究所,批號D01339、P0018FM);聚偏二氟乙烯(PVDF)膜、PureLink Micro-toMidi總RNA分離試劑盒(美國Invitrogen公司,批號FFP36、RRK-10328);RIPA裂解液、TaqMan逆轉錄試劑盒(哈爾濱海基生物科技深圳有限公司,批號KL6379、106734);Bradford蛋白質檢測試劑盒(南京建城生物工程研究所,批號GL1493);聚丙烯酰胺凝膠電泳(SDS-PAGE)(美國賽默飛世爾科技公司,批號SJ-394764);AMPK一抗、NR2B一抗、GAPDH抗體、HRP偶聯二抗(美國Santa Cruz Biotechnology公司,批號ac41789、ac52987、ad33117、ak636548);PBS-Tween20洗液(上海澤葉生物科技有限公司,批號BD-8599);DM500型熒光顯微鏡(德國徠卡公司);3900型RNA TaqMan microRNA分析儀(美國Applied Biosystems公司);qTOWER型實時熒光定量PCR儀(德國耶拿分析儀器股份公司);Epson Perfection 2480型掃描儀(日本長野精工公司)。
1.3動物建模及給藥:將Aβ1-42溶解在無菌蒸餾水中,濃度為5μg/L,并在37℃下孵育7d,以獲得聚集形式。模型組、鹽酸米諾環素組、丙泊酚低劑量組、丙泊酚高劑量組大鼠腹膜內注射戊巴比妥鈉(50mg/kg)麻醉,將Aβ1-42(1μL=5μg)微量注射到雙側海馬中。所有微注射均使用10μLHamilton注射器進行,將其用26號不銹鋼針頭緩慢放下。開始注射前,將針頭留在原位3~5min,然后以0.1μL/min的速度緩慢注射,在緩慢拔出針頭之前,將其再留置3~5min[7]。對照組進行相同的操作過程,以相同的立體定位法以0.1μL/min的速率將相同體積的載體溶液(1μLPBS)單次微量注射到海馬的相應區域中。注射后,將頭皮縫合,使大鼠從麻醉中恢復。模型建立成功后,鹽酸米諾環素組、丙泊酚低、高劑量組以10mL/kg的灌胃體積給予相應藥物灌胃,對照組、模型組灌胃10mL/kg的生理鹽水。每天給藥1次,持續4周。
1.4大鼠神經認知功能檢測:Morris水迷宮試驗測定逃避潛伏期時間、經過原平臺位置的次數、原平臺象限停留的時間[8]。
1.5大鼠海馬神經元病理結構觀察:頸椎脫臼處死大鼠,取大鼠海馬組織,常規脫水、包埋,制作切片,蘇木精-伊紅(HE)染色,顯微鏡觀察神經元病理結構變化。
1.6大鼠海馬神經元自噬檢測:吖啶橙(10mg/L)37℃染色海馬組織,持續30min,于熒光顯微鏡下觀察神經元自噬水平(細胞質內若出現酸性紅色自噬小體說明發生自噬;隨機計數200個細胞中自噬小體個數)。
1.7大鼠海馬組織AMPK、NR2B mRNA表達水平的測定:使用PureLink Micro-toMidi總RNA分離試劑盒提取大鼠海馬組織總RNA,使用TaqMan逆轉錄試劑盒在RNA TaqMan microRNA分析儀將(20 ng)RNA逆轉錄為cDNA,逆轉錄程序為16℃退火30min,然后在42℃延伸30min。采用實時熒光定量PCR法檢測AMPK、NR2B的表達,引物由蘇州泓迅生物科技股份有限公司合成。引物序列如下:AMPK正向引物:5'-CCTAAGGATCCAGGGAATCCCAT-3',反向引物:5'-CGGTATACGACCAAATCCGCCGA-3';NR2B正向引物:5'-CCGACGCCAATCTTGTCTCCTGGA-3',反向引物:5'-CGGATCCTGTTCGACAGTCACAAC-3';U6正向引物:5'-TGGAGTCCCTTGTGGTTCTGGAGTC-3',反向引物:5'-CGTGGGACTGACTGTTCGACCTGGA-3'。使用實時熒光定量PCR儀在一式三份的孔中進行循環PCR。程序為:95℃下變性15s,60℃下退火和延伸1min。使用SDS軟件確定循環閾值(Ct)熒光值。采用2-△△Ct法計算AMPK、NR2B mRNA的相對表達量。以U6作為內參基因。
1.8大鼠海馬組織AMPK、NR2B蛋白表達水平的測定:將海馬組織樣本在RIPA裂解液中進行裂解,然后在4℃下以12000×g離心20min。使用Bradford蛋白質檢測試劑盒來計算蛋白質濃度。樣品(每泳道60μg)通過8%SDS-PAGE分離并電轉移到PVDF膜上。在室溫下,用5%脫脂牛奶將膜封閉2h,并在4℃加入AMPK一抗(1∶500稀釋)、NR2B一抗(1∶500稀釋)、GAPDH抗體(1∶1000稀釋)。GAPDH被用作對照。將膜在PBS-Tween20洗液中洗滌10min,將其與HRP偶聯二抗(1∶4000稀釋)孵育2h。通過增強的化學發光(ECL)Western印跡檢測試劑盒將印跡的蛋白條帶可視化,并用X射線膠片曝光。使用Epson Perfection 2480型掃描儀將沖洗過的膠片數字化,使用Glyko Bandscan軟件獲得光密度。所有實驗重復3次。

2 結 果
2.1各組大鼠Morris水迷宮試驗結果比較:模型組逃避潛伏期時間較對照組明顯升高,經過原平臺位置的次數、原平臺象限停留的時間較對照組明顯降低(P<0.05);鹽酸米諾環素組和丙泊酚低、高劑量組逃避潛伏期時間較模型組明顯降低,經過原平臺位置的次數、原平臺象限停留的時間較模型組明顯升高(P<0.05);丙泊酚低劑量組逃避潛伏期時間較鹽酸米諾環素組明顯升高,經過原平臺位置的次數、原平臺象限停留的時間較鹽酸米諾環素組明顯降低(P<0.05);丙泊酚高劑量組與鹽酸米諾環素組相比無明顯改變(P>0.05)。見表1。

表1 各組大鼠Morris水迷宮試驗結果比較
表2 各組大鼠海馬組織神經元自噬小體水平比較
2.2各組大鼠海馬組織神經元自噬小體水平比較:模型組神經元自噬小體水平較對照組明顯降低(P<0.05);鹽酸米諾環素組和丙泊酚低、高劑量組神經元自噬小體水平較模型組明顯升高(P<0.05);丙泊酚低劑量組神經元自噬小體水平較鹽酸米諾環素組明顯降低(P<0.05);丙泊酚高劑量組與鹽酸米諾環素組相比無明顯改變(P>0.05)。見表2。
2.3各組大鼠海馬神經元病理結構比較:對照組海馬區神經元細胞較多,排列整齊;模型組海馬區見片狀神經元壞死,數目減少,核固縮明顯;丙泊酚低劑量組壞死神經元細胞減少;鹽酸米諾環素組、丙泊酚高劑量組海馬區見少量壞死神經元細胞,神經元細胞結構完整。見圖1。
2.4各組大鼠海馬組織AMPK、NR2B mRNA和蛋白表達水平比較:模型組AMPK、NR2B mRNA和蛋白表達水平較對照組明顯升高(P<0.05);鹽酸米諾環素組和丙泊酚低、高劑量組AMPK、NR2B mRNA和蛋白表達水平較模型組明顯降低(P<0.05);丙泊酚低劑量組AMPK、NR2B mRNA和蛋白表達水平較鹽酸米諾環素組明顯升高(P<0.05);丙泊酚高劑量組與鹽酸米諾環素組相比無明顯改變(P>0.05)。見表3、圖2。
表3 各組大鼠海馬組織AMPK NR2B mRNA和蛋白表達水平比較
.
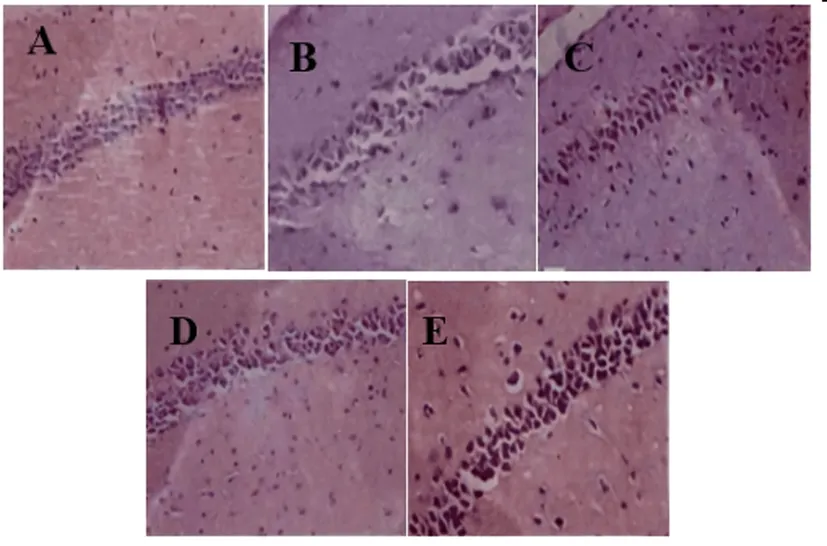
圖1 各組大鼠海馬神經元病理結構圖(HE染色,400倍)
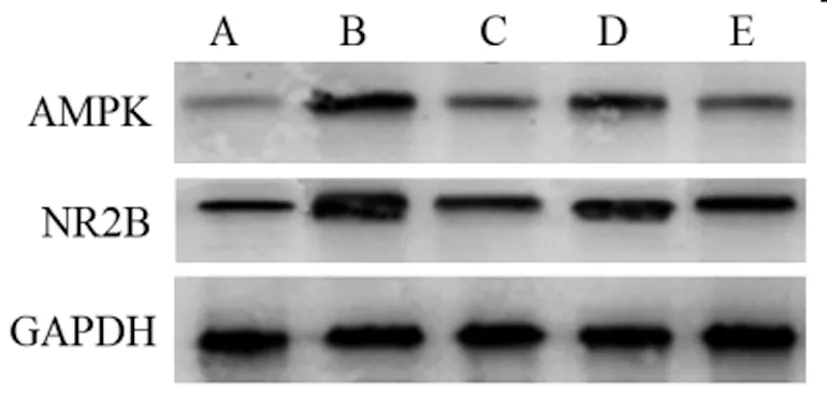
圖2 各組大鼠海馬組織AMPK、NR2B蛋白表達印跡圖
3 討 論
老年癡呆主要病理變化是神經原纖維纏結,老年斑沉積和神經元細胞死亡。Aβ已被證明是老年斑中的關鍵成分,而tau蛋白的過度磷酸化可促進神經原纖維纏結[9]。有研究表明自噬對老年癡呆具有有益作用[10]。Aβ1-42或β樣淀粉蛋白1-40(Aβ1-40)及其前體淀粉樣前體蛋白(APP)和β切割的APP的羧基末端結構域富含自噬空泡(AVs)在自噬中異常積累。AVs積累和溶酶體蛋白水解缺陷的自噬-溶酶體途徑功能失調發生在老年癡呆腦神經中。AV的大量積累是老年癡呆癥中Aβ產生的原因,因此,自噬缺陷有助于異常的老年癡呆進程。本次研究發現,模型組逃避潛伏期時間高于對照組,經過原平臺位置次數、原平臺象限留時間、神經元自噬小體水平低于對照組;鹽酸米諾環素組和丙泊酚劑量組逃避潛伏期時間明顯低于模型組,經過原平臺位置的次數、原平臺象限停留的時間、神經元自噬小體水平高于模型組,且隨著丙泊酚給藥劑量的增加,呈劑量依賴性。這說明丙泊酚能降低老年癡呆大鼠神經認知功能損害,增加海馬神經元自噬水平。大鼠海馬神經元結構病理學結果顯示,對照組海馬區神經元細胞結構正常;模型組海馬區見片狀神經元壞死,數目減少,核固縮明顯;丙泊酚低劑量組壞死神經元細胞減少;鹽酸米諾環素組、丙泊酚高劑量組海馬區見少量壞死神經元細胞,神經元細胞結構完整。這提示丙泊酚能明顯減輕老年癡呆大鼠海馬神經元損傷程度。
AMPK通路的激活在自噬中具有重要作用。AMPK是一種能量傳感器,可以調節急性和慢性腦損傷中的炎癥反應,氧化應激和突觸可塑性。活化的AMPK通過下調磷脂酰肌醇3-激酶/p38絲裂原活化蛋白激酶(PI3K/p38)、絲裂原活化蛋白激酶(MAPK)、核因子kappa B(NF-kB)和Nod -類受體蛋白3(NLRP3)的產生[11]。手術后老年大鼠海馬中的AMPK激活增強,而成年幼鼠海馬中的AMPK激活增強,這與認知障礙發生的年齡依賴性有關[12]。此外,抑制AMPK激活不僅顯著改善了老年大鼠的認知障礙,而且還限制了小膠質細胞的激活,NR2B表達的下調以及手術后老年大鼠海馬中Tau和Tau磷酸化表達的上調[13]。這些數據表明AMPK可能是上游信號分子,啟動了認知障礙的病理過程。本研究結果顯示:模型組AMPK、NR2B mRNA和蛋白表達水平明顯高于對照組;鹽酸米諾環素組和丙泊酚低、高劑量組AMPK、NR2B mRNA和蛋白表達水平明顯低于模型組,且隨著丙泊酚給藥劑量的增加,AMPK、NR2B mRNA和蛋白表達水平逐漸降低,呈劑量依賴性。這提示丙泊酚抑制AMPK信號通路誘導的大鼠神經元自噬來削弱老年癡呆大鼠神經損傷。
綜上所述,丙泊酚能增加老年癡呆大鼠神經元自噬水平進而明顯減輕海馬神經元損傷程度,其機制與丙泊酚抑制AMPK信號通路誘導的大鼠神經元自噬來削弱老年癡呆大鼠神經損傷有關。
猜你喜歡
課堂內外·初中版(科學少年)(2023年10期)2023-12-10 00:43:06
全科護理(2022年10期)2022-12-26 21:19:15
中國合理用藥探索(2022年1期)2022-11-26 00:22:32
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
鄉村科技(2021年33期)2021-03-16 02:26:54
國際放射醫學核醫學雜志(2021年10期)2021-02-28 08:41:58
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
大灰狼(2015年6期)2015-07-16 21:01:00
作文與考試·小學高年級版(2015年17期)2015-05-30 10:48:04
